
Early Evidence That Soil Dryness Causes Widespread Decline in Grassland Productivity in China
 , , and
, , and
Abstract
:1. Introduction
- (1)
- To analyze the interannual variability of SM and GPP in Chinese grasslands and to explore the mechanisms of SM regulation of GPP in Chinese grassland ecosystems during historical and future periods.
- (2)
- To compare the correlation between the effects of SM on ecosystem GPP in different soil layers, and analyze the conditional probability of drought in soils of different soil layers causing a decline in GPP in Chinese grasslands.
- (3)
- To calculate the difference between the probability of ecosystem loss due to soil drought minus the probability of ecosystem loss due to atmospheric drought and determine the key moisture constraints controlling GPP in Chinese grasslands.
2. Materials and Methods
2.1. Materials
2.1.1. GPP Datasets
2.1.2. SM Datasets
2.1.3. VPD Datasets
2.1.4. Definition of Warm Season and Screening for Warm-Season GPP, SM, and VPD
2.2. Methods
2.2.1. Interannual Correlation Measures
2.2.2. Bivariate Linkage to Calculate the Probability of Conditions under Soil (or Atmospheric) Drought Conditions
3. Results
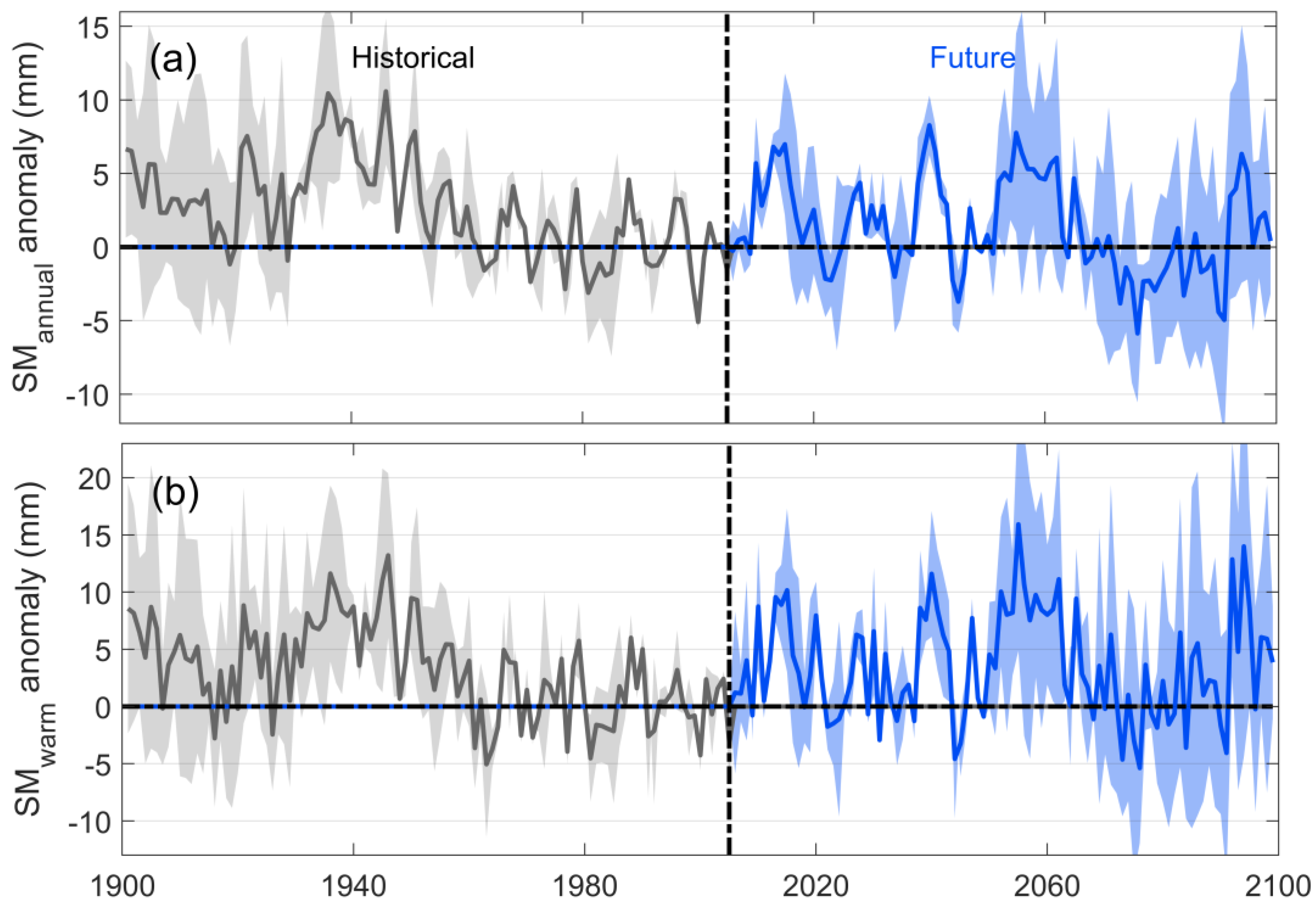
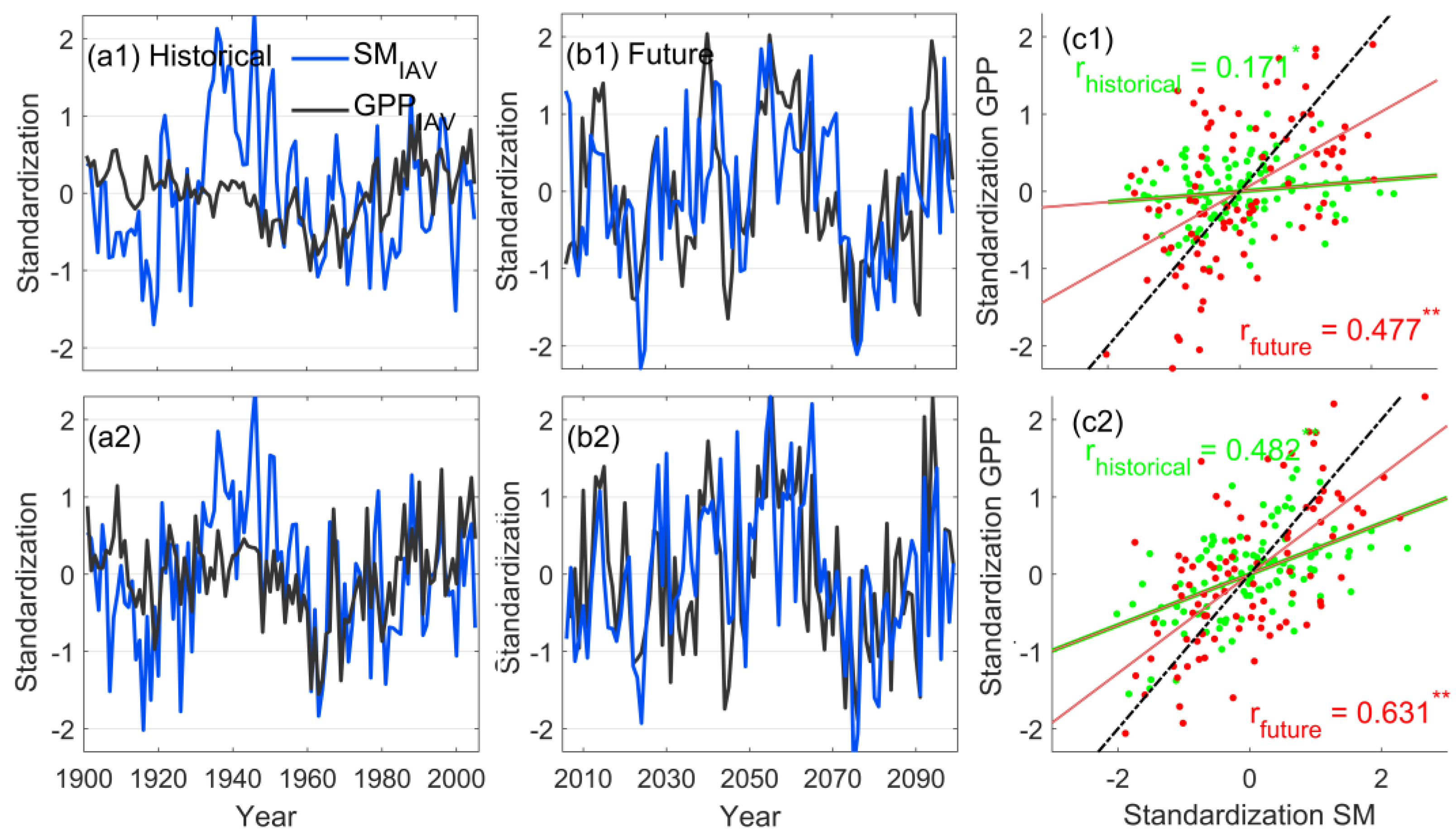
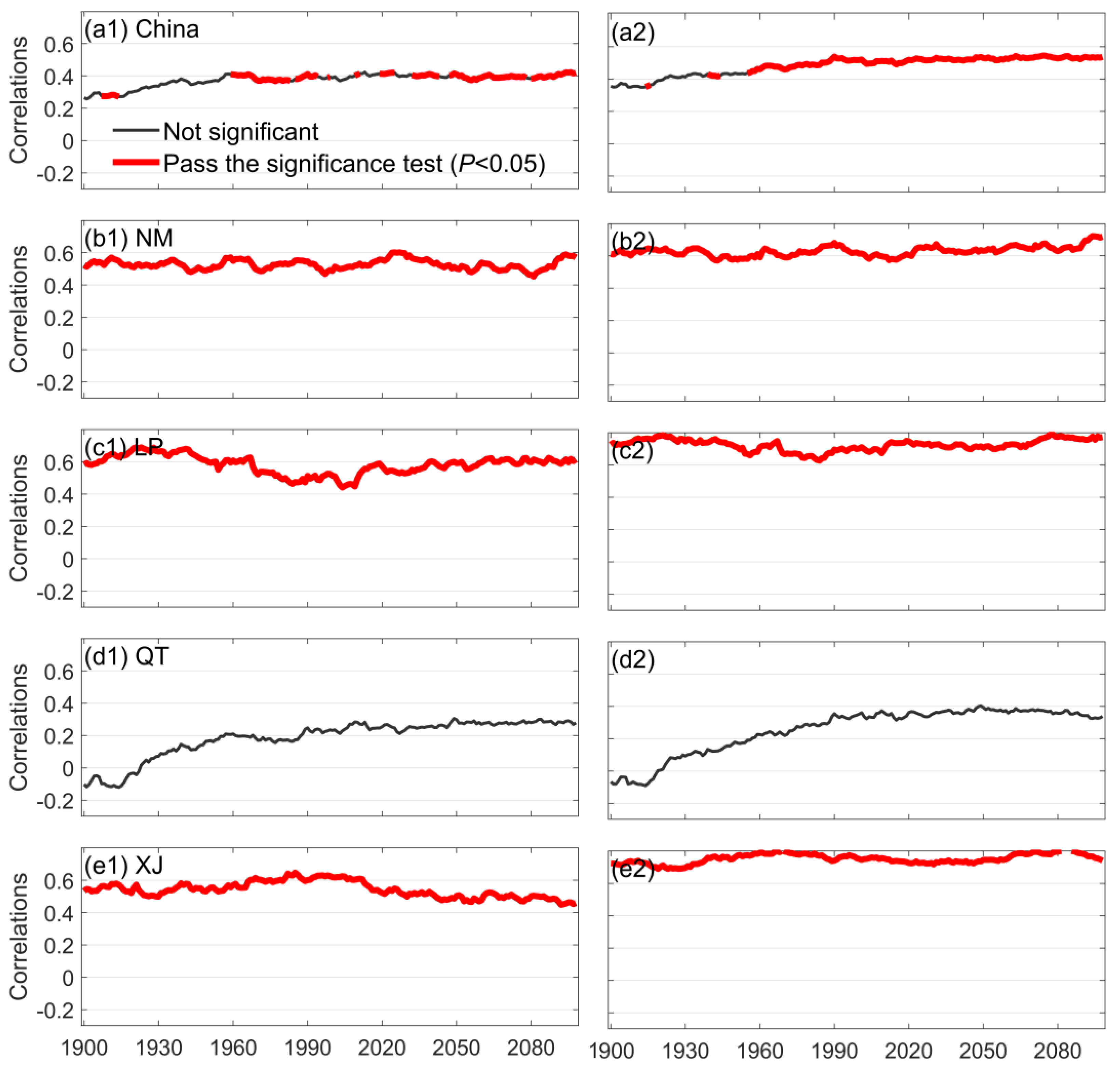
3.1. Characteristics of Long-Term Changes in Chinese Grassland SM and Its Constraints on Ecosystem GPP
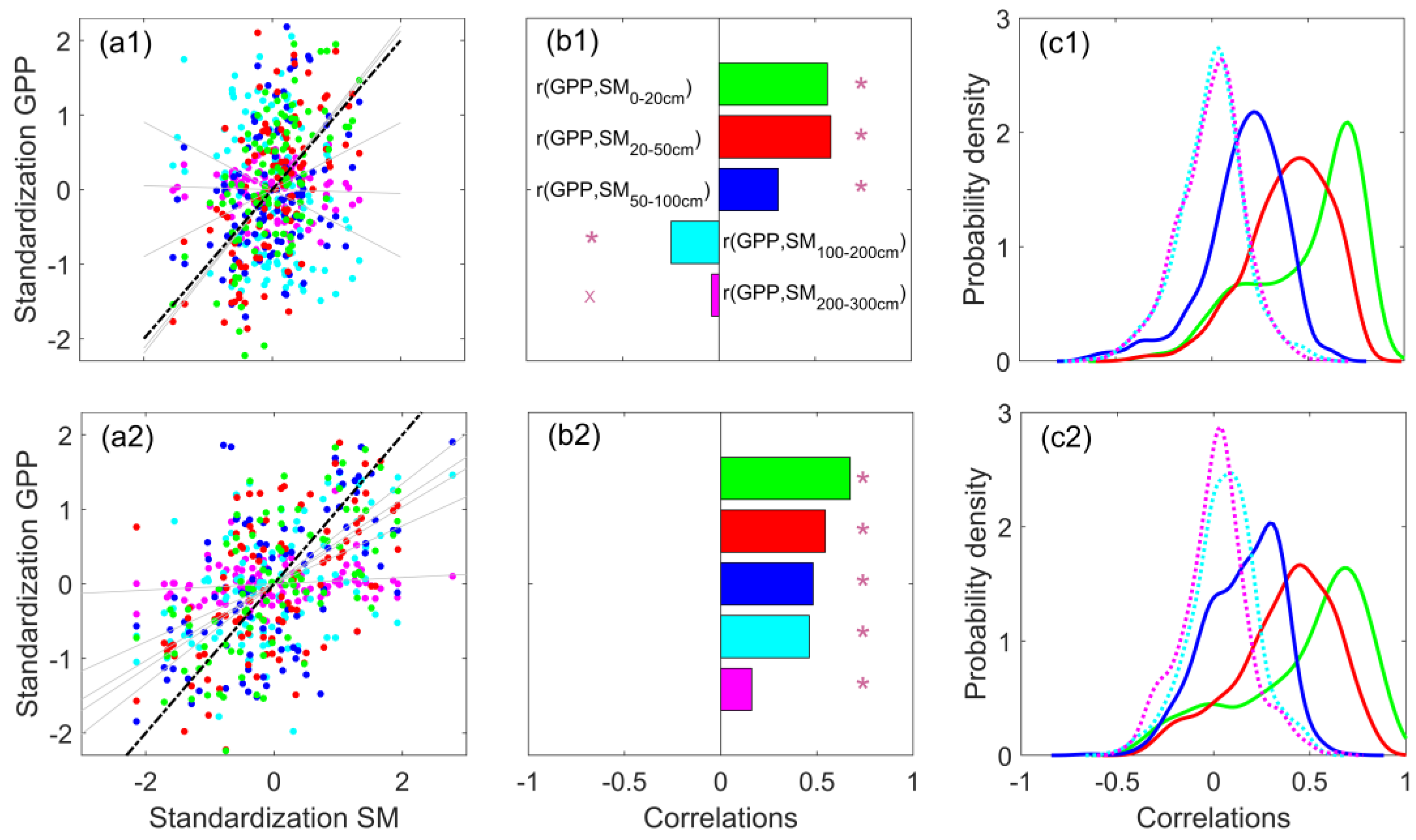
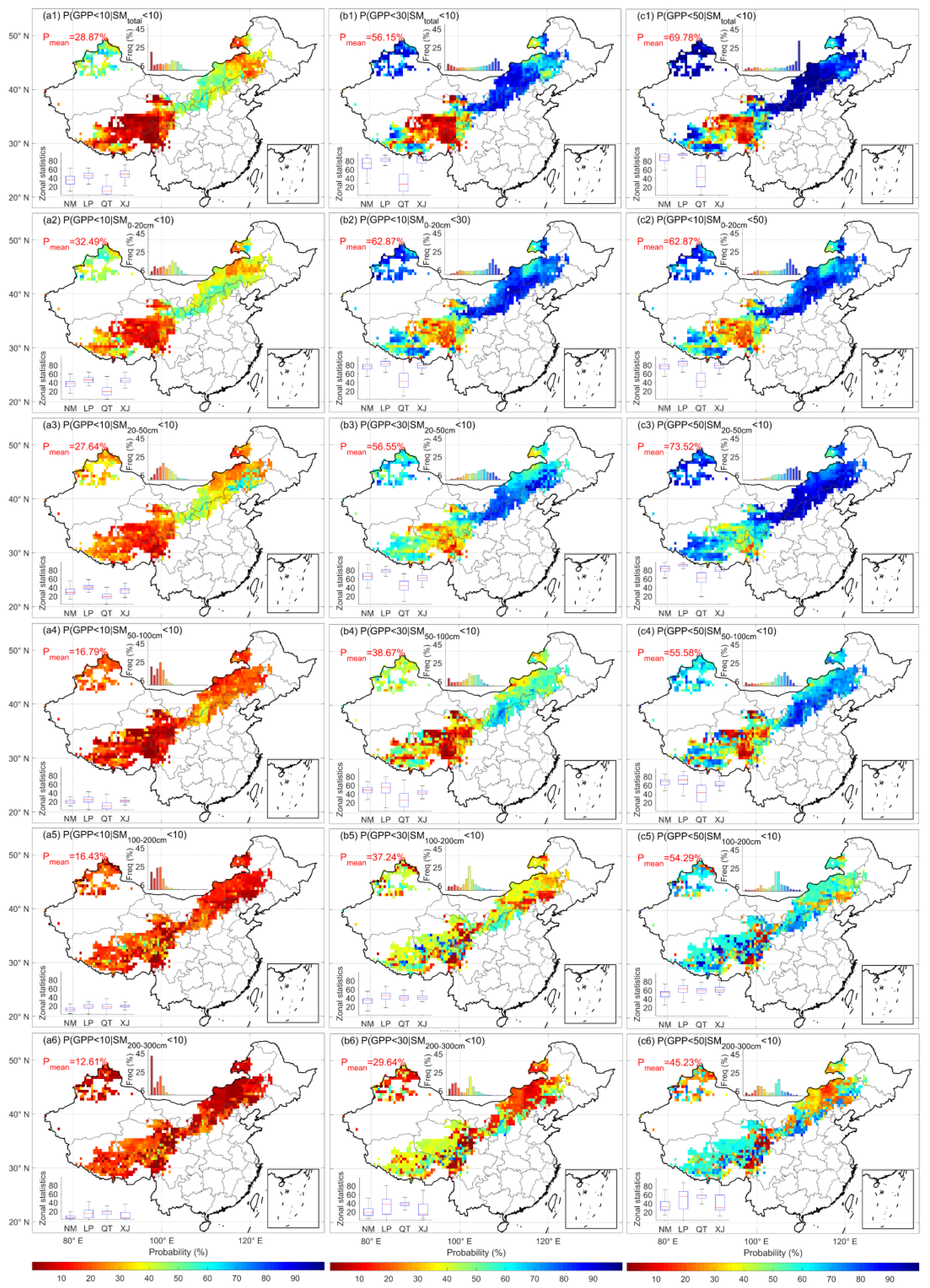
3.2. Comparison of the Regulation of GPP by Different Soil Layers
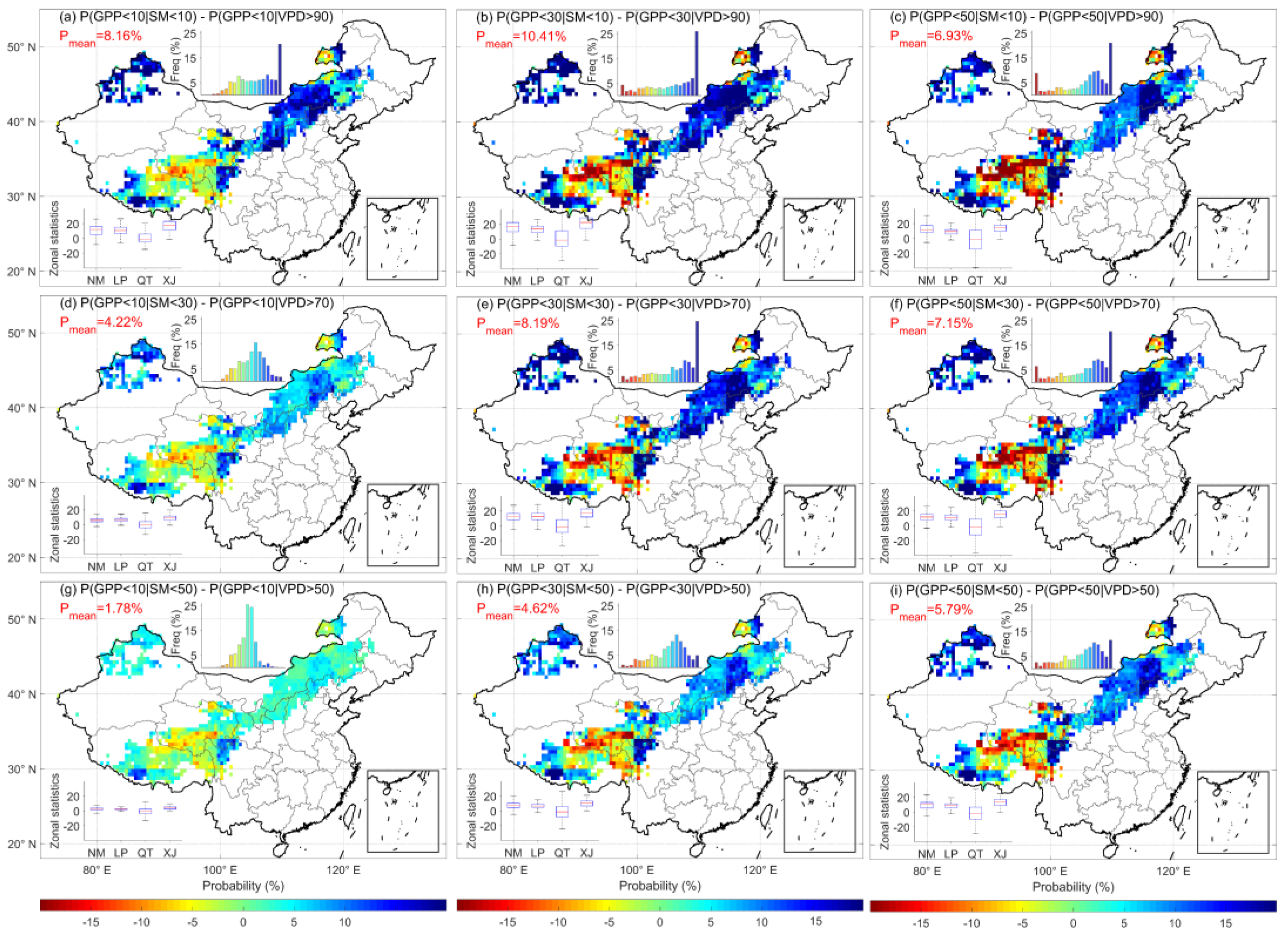
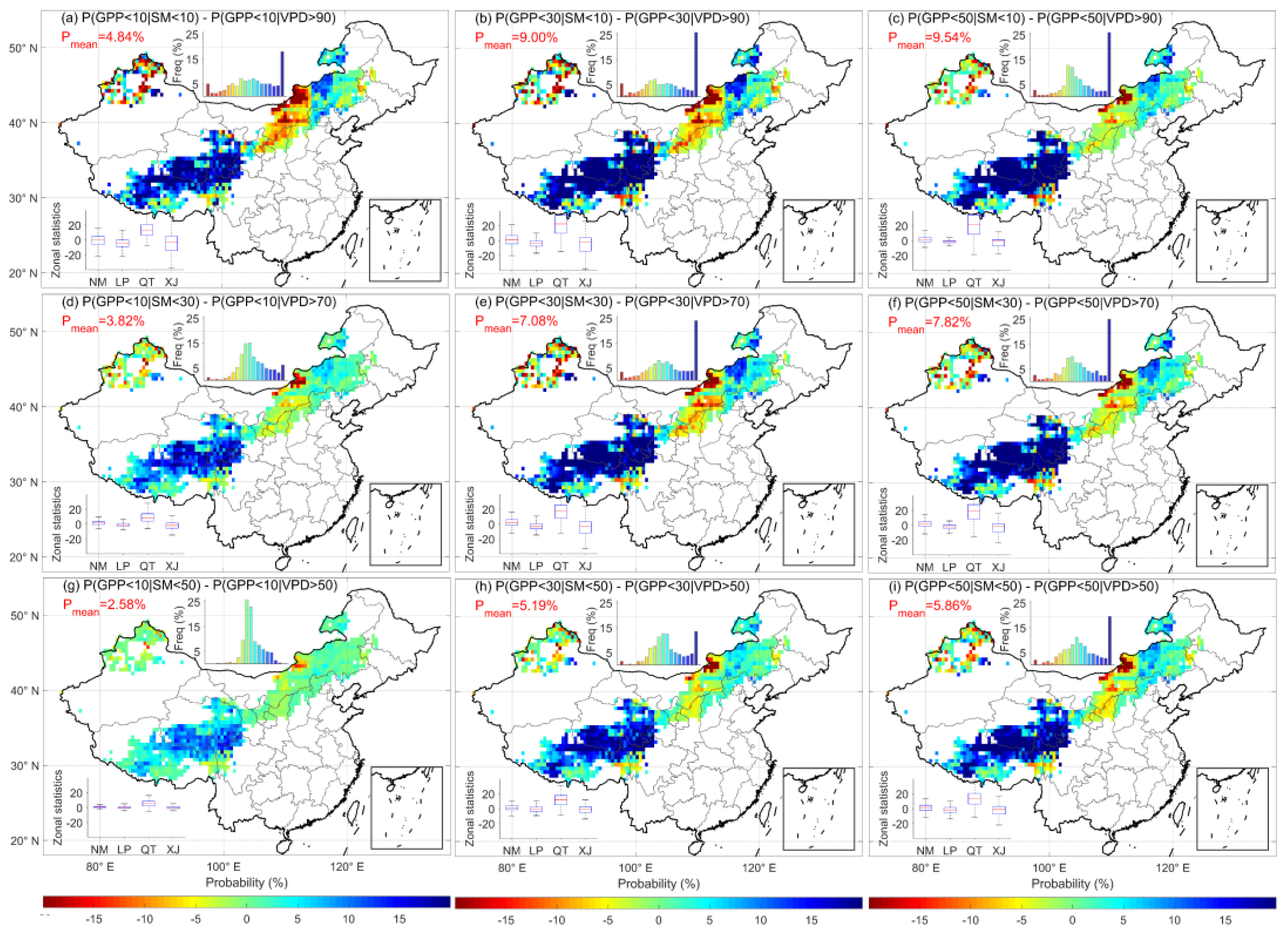
3.3. Comparison of the Probability of High-VPD and Low-SM Events Leading to Ecosystem GPP Deficits
4. Discussion
4.1. Soil Moisture More Strongly Regulates Carbon Balance Than Atmospheric Indicators in Chinese Grasslands
4.2. Soil Moisture Is a Key Water Constraint Controlling the Grassland Productivity in China
5. Conclusions
- (1)
- No significant trends were found for soil moisture in the historical or future periods, and its long-term change was mainly reflected through interannual fluctuations. Soil moisture showed a highly significant positive correlation with ecosystem GPP in the time series, indicating that when soil water decreases, it causes a decrease in ecosystem GPP. Moreover, the correlation between SM and GPP was higher in the warm season than annually, and higher in the future period than in the historical period, representing a stronger constraint on GPP in Chinese grasslands in the warm season and a deeper constraint in the future period than in the historical period.
- (2)
- Using the LPJmL model’s soil moisture data at different soil depths and analyzing their relationship with ecosystem GPP, it was found that the correlation between shallow-soil moisture (0–50 cm) and GPP was clearly higher than that of deeper soils, and the probability of an ecosystem GPP deficit due to a shortage of soil water in the shallow layer was much higher than that of soil water in the middle and deep layers.
- (3)
- In probabilistic terms, soil drought has a higher probability of initiating the loss of ecosystem GPP than atmospheric drought, with moisture scarcity originating from the soil becoming the main aspect that constrains ecosystem GPP. In the future, with the rapid rise of global VPD, the probability of ecosystem GPP loss induced by atmospheric drought increases and overtakes soil drought as the main water constraint in some regions.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogee, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Xia, J.; Li, Z.; Fang, C.; Deng, X. Spatio-temporal dynamics of water-related disaster risk in the Yangtze River Economic Belt from 2000 to 2015. Resour. Conserv. Recycl. 2020, 161, 104851. [Google Scholar] [CrossRef]
- Xiao, J.; Sun, G.; Chen, J.; Chen, H.; Chen, S.; Dong, G.; Gao, S.; Guo, H.; Guo, J.; Han, S.; et al. Carbon fluxes, evapotranspiration, and water use efficiency of terrestrial ecosystems in China. Agric. For. Meteorol. 2013, 182, 76–90. [Google Scholar] [CrossRef]
- Yang, Y.; Guan, H.; Batelaan, O.; McVicar, T.; Long, D.; Piao, S.; Liang, W.; Liu, B.; Jin, Z.; Simmons, C. Contrasting response of water use efficiency to drought across global terrestrial ecosystems. Sci. Rep. 2016, 6, 23283. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xiao, X.; Zhou, S.; Ciais, P.; McCarthy, H.; Luo, Y. Canopy and physiological control of GPP during drought and heatwave. Geophys. Res. Lett. 2016, 43, 3325–3333. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, V.; Zscheischler, J.; Ciais, P.; Gudmundsson, L.; Sitch, S.; Seneviratne, S.I. Sensitivity of atmospheric CO2 growth rate to observed changes in terrestrial water storage. Nature 2018, 560, 628–631. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Ren, J. Simulation of Gross Primary Productivity Using Multiple Light Use Efficiency Models. Land 2021, 10, 329. [Google Scholar] [CrossRef]
- Liancourt, P.; Sharkhuu, A.; Ariuntsetseg, L.; Boldgiv, B.; Helliker, B.R.; Plante, A.F.; Petraitis, P.S.; Casper, B.B. Temporal and spatial variation in how vegetation alters the soil moisture response to climate manipulation. Plant and Soil 2011, 351, 249–261. [Google Scholar] [CrossRef]
- Han, Z.; Huang, Q.; Huang, S.; Leng, G.; Bai, Q.; Liang, H.; Wang, L.; Zhao, J.; Fang, W. Spatial-temporal dynamics of agricultural drought in the Loess Plateau under a changing environment: Characteristics and potential influencing factors. Agric. Water Manag. 2021, 244, 106540. [Google Scholar] [CrossRef]
- He, P.; Sun, Z.; Han, Z.; Ma, X.; Zhao, P.; Liu, Y.F.; Ma, J. Divergent trends of water storage observed via gravity satellite across distinct areas in China. Water 2020, 12, 2862. [Google Scholar] [CrossRef]
- Sehgal, V.; Gaur, N.; Mohanty, B. Global surface soil moisture drydown patterns. Water Resour. Res. 2021, 57, e2020WR027588. [Google Scholar] [CrossRef]
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5, eaax1396. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Gao, G.; Fu, B.; Lu, Y. Responses of shelterbelt stand transpiration to drought and groundwater variations in an arid inland river basin of Northwest China. J. Hydrol. 2015, 531, 738–748. [Google Scholar] [CrossRef]
- Novick, K.; Ficklin, D.; Stoy, P.; Williams, C.A.; Bohrer, G.; Oishi, A.C.; Papuga, S.A.; Blanken, P.D.; Noormets, A.; Sulman, B.N.; et al. The increasing importance of atmospheric demand for ecosystem water and carbon fluxes. Nat. Clim. Change 2016, 6, 1023–1027. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Gudmundsson, L.; Hauser, M.; Qin, D.; Li, S.; Seneviratne, S.I. Soil moisture dominates dryness stress on ecosystem production globally. Nat. Commun. 2020, 11, 4892. [Google Scholar] [CrossRef]
- Kang, L.; Han, X.; Zhang, Z.; Sun, O.J. Grassland ecosystems in China: Review of current knowledge and research advancement. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2007, 362, 997–1008. [Google Scholar] [CrossRef]
- Jia, W.; Liu, M.; Wang, D.; He, H.; Shi, P.; Li, Y.; Wang, Y. Uncertainty in simulating regional gross primary productivity from satellite-based models over northern China grassland. Ecol. Indic. 2018, 88, 134–143. [Google Scholar] [CrossRef]
- Liu, Y.; Lei, H. Responses of natural vegetation dynamics to climate drivers in China from 1982 to 2011. Remote Sens. 2015, 7, 10243–10268. [Google Scholar] [CrossRef] [Green Version]
- Rui, Z.; Zhao, X.; Zuo, X.; Degen, A.; Yulin, L.; Liu, X.; Luo, Y.; Qu, H.; Lian, J.; Wang, R. Drought-induced shift from a carbon sink to a carbon source in the grasslands of Inner Mongolia, China. Catena 2020, 195, 104845. [Google Scholar] [CrossRef]
- He, P.; Ma, X.; Sun, Z. Interannual variability in summer climate change controls GPP long-term changes. Environ. Res. 2022, 212, 113409. [Google Scholar] [CrossRef]
- Cheng, S.; Huang, J.; Ji, F.; Lin, L. Uncertainties of soil moisture in historical simulations and future projections: Uncertainties of Soil Moisture. J. Geophys. Res. Atmos. 2017, 122, 2239–2253. [Google Scholar] [CrossRef]
- Buck, A. New equations for computing vapor pressure and enhancement Factor. J. Appl. Meteorol. 1981, 20, 1527–1532. [Google Scholar] [CrossRef]
- Zscheischler, J.; Seneviratne, S. Dependence of drivers affects risks associated with compound events. Sci. Adv. 2017, 3, e1700263. [Google Scholar] [CrossRef] [Green Version]
- Bishara, A.; Hittner, J. Testing the significance of a correlation with nonnormal data: Comparison of pearson, spearman, transformation, and resampling approaches. Psychol. Methods 2012, 17, 399–417. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Huang, S.; Huang, Q.; Leng, G.; Han, Z.; Zhao, J.; Guo, Y. Vegetation vulnerability and resistance to hydrometeorological stresses in water- and energy-limited watersheds based on a Bayesian framework. Catena 2021, 196, 104879. [Google Scholar] [CrossRef]
- Jones, M.; Hardle, W. Applied nonparametric regression. Economica 1992, 58, 538. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Dijkstra, P.; Koch, G.; Penuelas, J.; Hungate, B. Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation. Glob. Change Biol. 2011, 17, 927–942. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.; Chen, J.; Chu, H.; Lafortezza, R.; Dong, G.; Abraha, M.; Ochirbat, B.; John, R.; Ouyang, Z.; Zhang, Y.; et al. Grassland productivity and carbon sequestration in Mongolian grasslands: The underlying mechanisms and nomadic implications. Environ. Res. 2017, 159, 124–134. [Google Scholar] [CrossRef]
- Arca, V.; Power, S.A.; Delgado-Baquerizo, M.; Pendall, E.; Ochoa-Hueso, R. Seasonal effects of altered precipitation regimes on ecosystem-level CO2 fluxes and their drivers in a grassland from Eastern Australia. Plant Soil 2021, 460, 435–451. [Google Scholar] [CrossRef]
- Ren, H.; Xu, Z.; Huang, J.; Lü, X.; Zeng, D.-H.; Yuan, Z.; Han, X.; Fang, Y. Increased precipitation induces a positive plant-soil feedback in a semi-arid grassland. Plant Soil 2014, 389, 211–223. [Google Scholar] [CrossRef]
- Suttle, K.; Thomsen, M.; Power, M. Species interactions reverse grassland responses to changing climate. Science 2007, 315, 640–642. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Wang, X.; Lu, T.; Du, H.; Ciais, P.; Lin, X. Divergent seasonal responses of carbon fluxes to extreme droughts over China. Agric. For. Meteorol. 2023, 328, 109253. [Google Scholar] [CrossRef]
- Ru, J.; Zhou, Y.; Hui, D.; Zheng, M.; Wan, S. Shifts of growing-season precipitation peaks decrease soil respiration in a semiarid grassland. Glob. Change Biol. 2017, 24, 1001–1011. [Google Scholar] [CrossRef]
- Huxman, T.; Snyder, K.; Tissue, D.; Leffler, A.J.; Ogle, K.; Pockman, W.; Sandquist, D.; Potts, D.; Schwinning, S. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 2004, 141, 254–268. [Google Scholar] [CrossRef]
- Nielsen, U.; Ball, B. Impacts of altered precipitation regimes on soil communities and biogeochemistry in arid and semi-arid ecosystems. Glob. Change Biol. 2014, 21, 1407–1421. [Google Scholar] [CrossRef]
- Parton, W.; Morgan, J.; Smith, D.; Del Grosso, S.; Prihodko, L.; Lecain, D.; Kelly, R.; Lutz, S. Impact of precipitation dynamics on net ecosystem productivity. Glob. Change Biol. 2012, 18, 915–927. [Google Scholar] [CrossRef]
- Parker, N.; Patrignani, A. Reconstructing Precipitation Events Using Co-located Soil Moisture Information. J. Hydrometeorol. 2021, 22, 3275–3290. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Ma, X.; Zongjiu, S.; Han, Z.; Ma, S.; Meng, X. Compound drought constrains gross primary productivity in Chinese grasslands. Environ. Res. Lett. 2022, 17, 104054. [Google Scholar] [CrossRef]
- Allen-Diaz, B.H. Water Table and Plant Species Relationships in Sierra Nevada Meadows. Am. Midl. Nat. 1991, 126, 30–43. [Google Scholar] [CrossRef]
- Darrouzet-Nardi, A.; D’Antonio, C.M.; Dawson, T.E. Depth of water acquisition by invading shrubs and resident herbs in a Sierra Nevada meadow. Plant Soil 2006, 285, 31–43. [Google Scholar] [CrossRef]
- Hoekstra, N.; Finn, J.; Hofer, D.; Luscher, A. The effect of drought and interspecific interactions on depth of water uptake in deep- and shallow-rooting grassland species as determined by delta O-18 natural abundance. Biogeosciences 2014, 11, 4493–4506. [Google Scholar] [CrossRef] [Green Version]
- Yongming, C.; Liu, L.; Cheng, L.; Fa, K.; Liu, X.; Huo, Z.; Huang, G. A shift in the dominant role of atmospheric vapor pressure deficit and soil moisture on vegetation greening in China. J. Hydrol. 2023, 615, 128680. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Copula | Expression of Distribution Function C(u,v) | Range of θ Values |
|---|---|---|
| Clayton | ||
| Frank | ||
| Gumbel | ||
| t | ||
| Gaussian |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, P.; Zeng, Y.; Wang, N.; Han, Z.; Meng, X.; Dong, T.; Ma, X.; Ma, S.; Ma, J.; Sun, Z. Early Evidence That Soil Dryness Causes Widespread Decline in Grassland Productivity in China. Land 2023, 12, 484. https://doi.org/10.3390/land12020484
He P, Zeng Y, Wang N, Han Z, Meng X, Dong T, Ma X, Ma S, Ma J, Sun Z. Early Evidence That Soil Dryness Causes Widespread Decline in Grassland Productivity in China. Land. 2023; 12(2):484. https://doi.org/10.3390/land12020484
Chicago/Turabian StyleHe, Panxing, Yiyan Zeng, Ningfei Wang, Zhiming Han, Xiaoyu Meng, Tong Dong, Xiaoliang Ma, Shangqian Ma, Jun Ma, and Zongjiu Sun. 2023. "Early Evidence That Soil Dryness Causes Widespread Decline in Grassland Productivity in China" Land 12, no. 2: 484. https://doi.org/10.3390/land12020484