
People, Crops, and Bee Farming: Landscape Models for a Symbiotic Network in Greece
Abstract
:1. Introduction
1.1. Bees’ Ecological Services in Regenerative Agriculture and Posed Threats
1.2. Disturbed Environments and Apiculture in the Greek Context
2. Materials and Methods
2.1. Landscape Configuration and Landscape Composition as Determinant Contexts
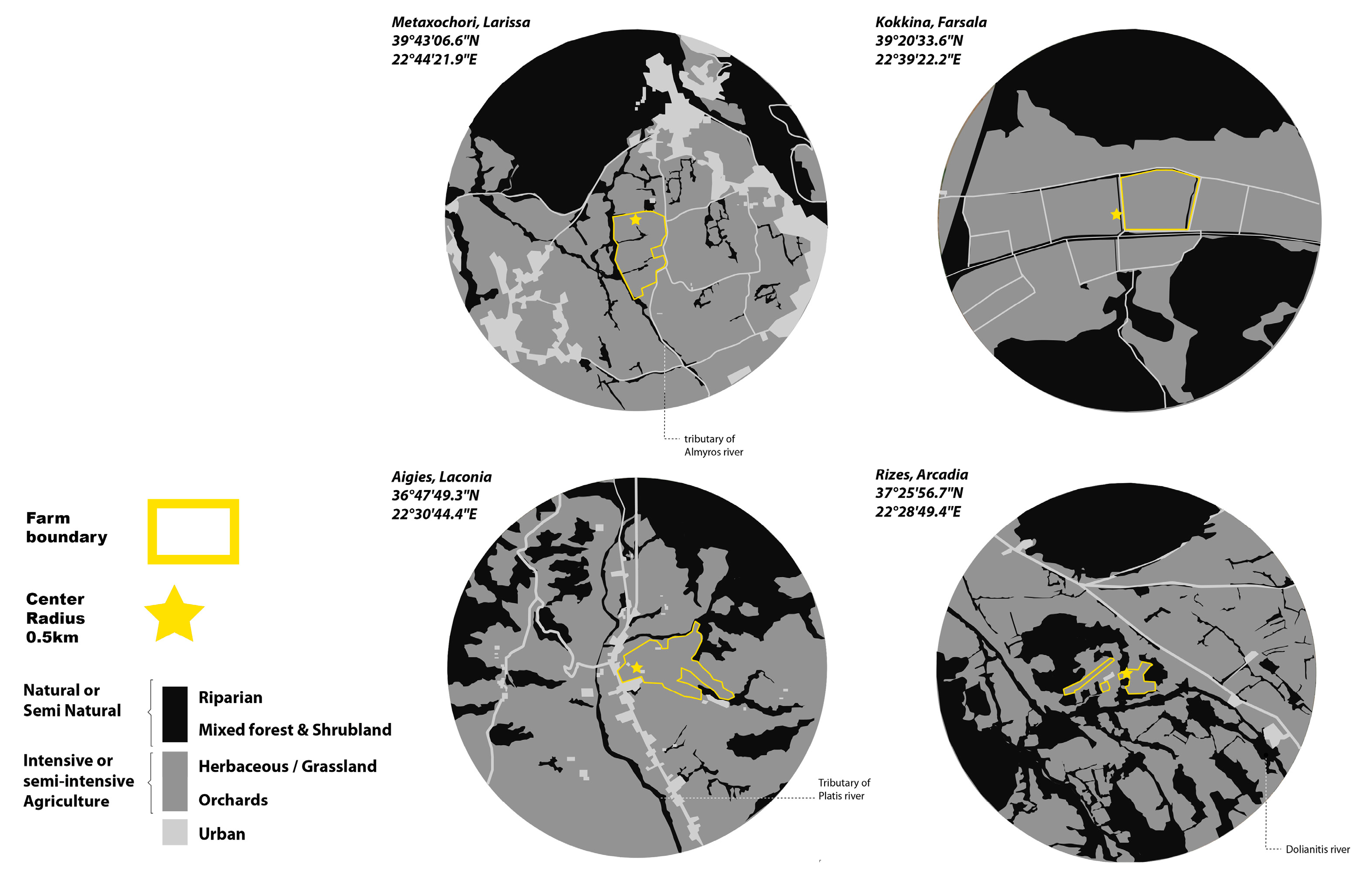
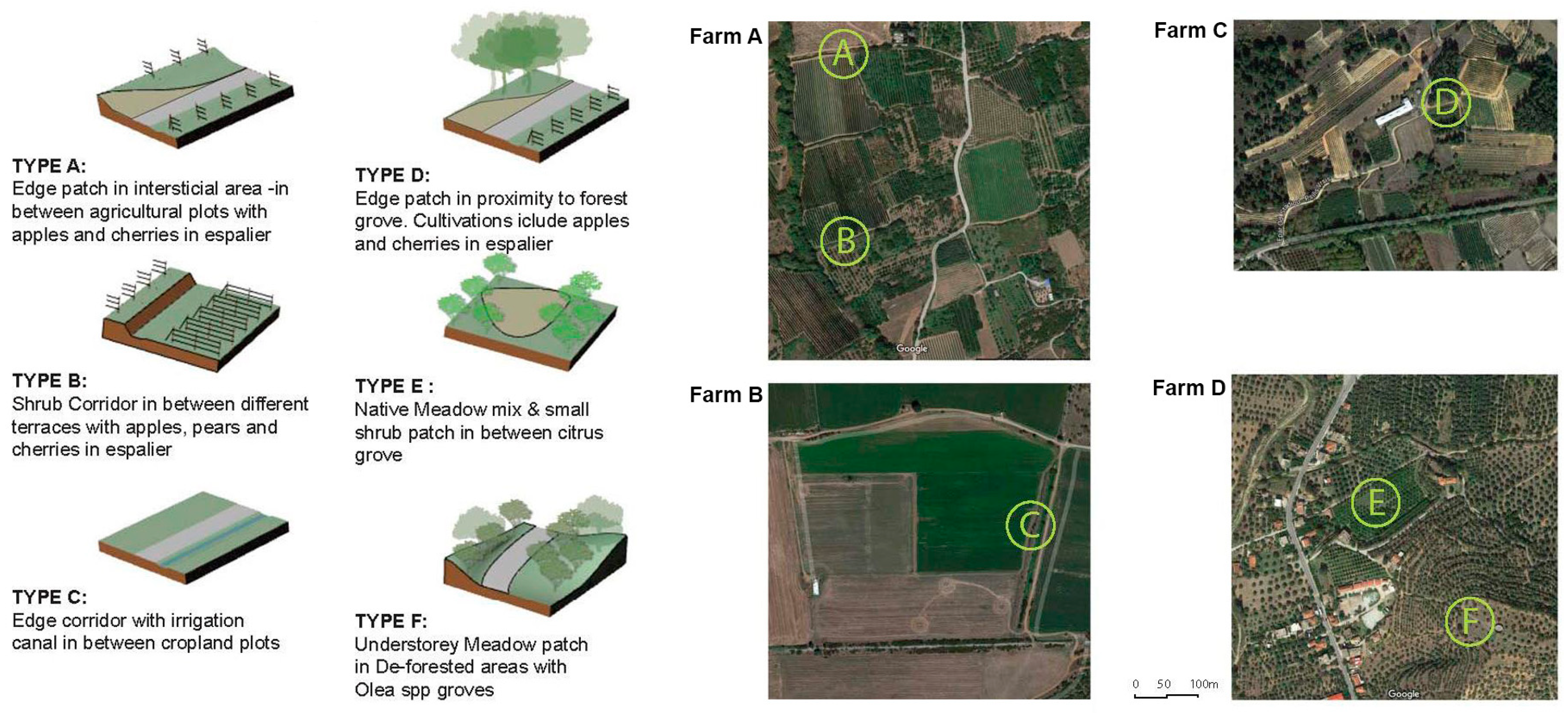
2.2. Study Areas
2.3. Methodology
- (a)
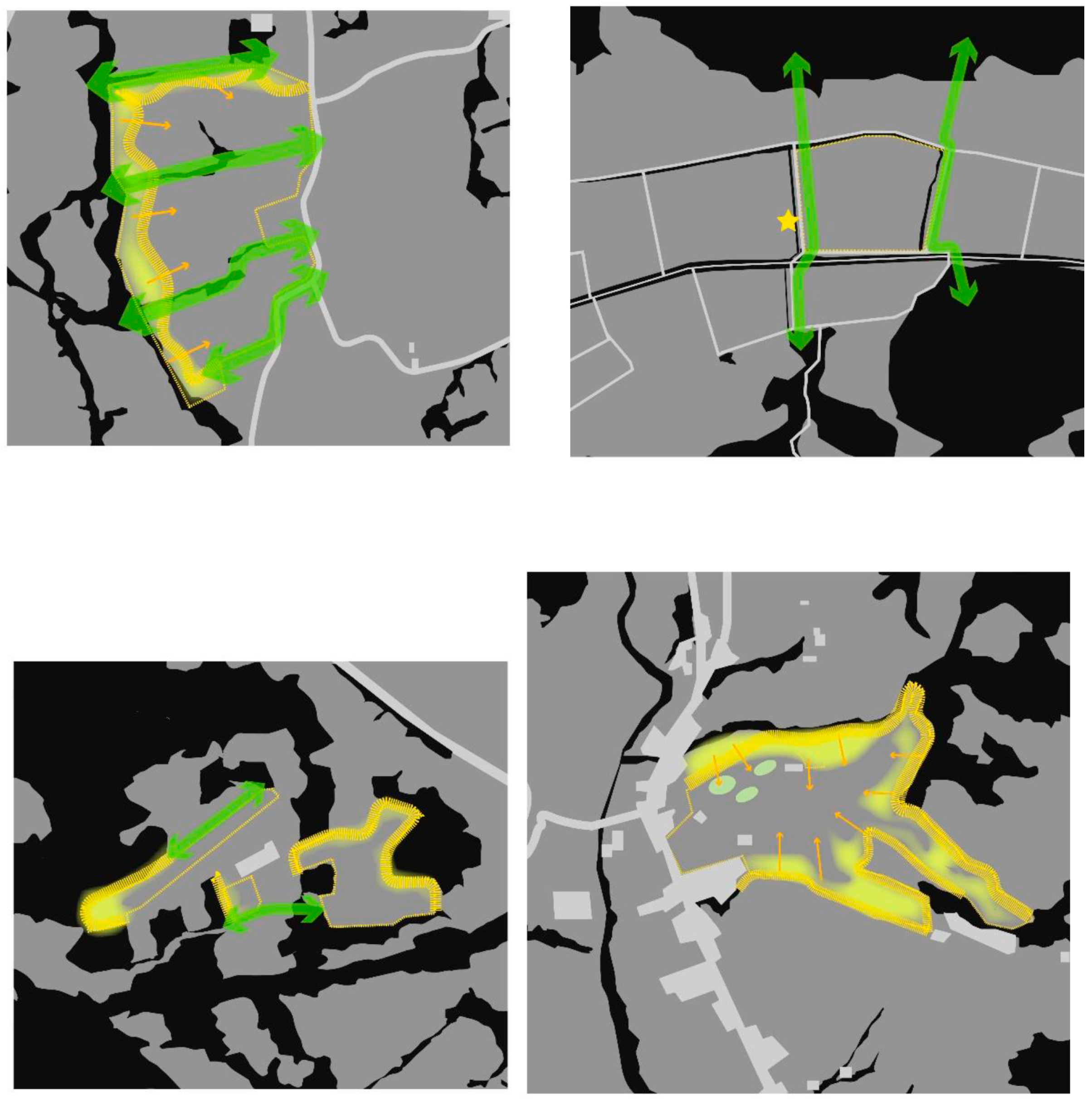
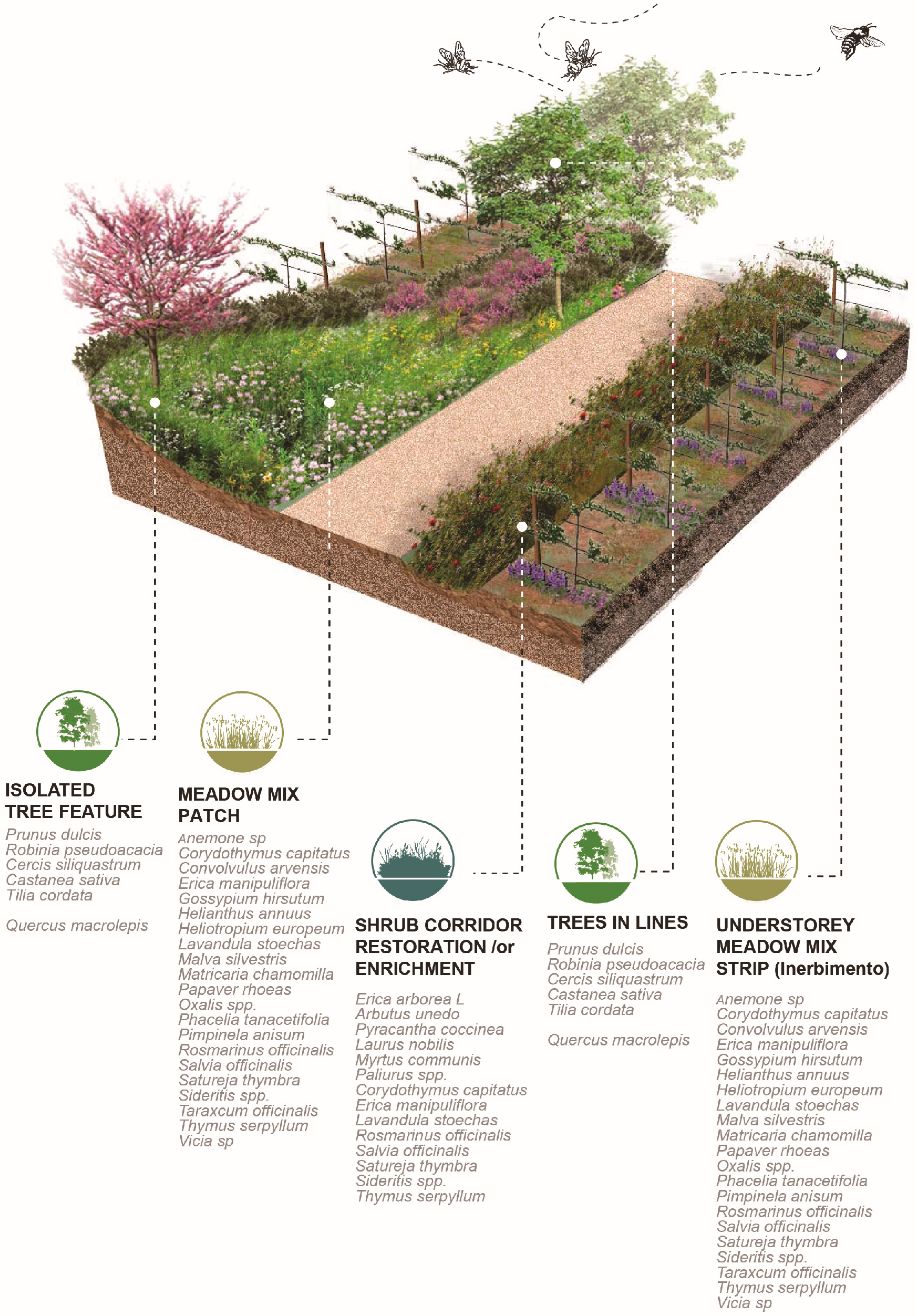
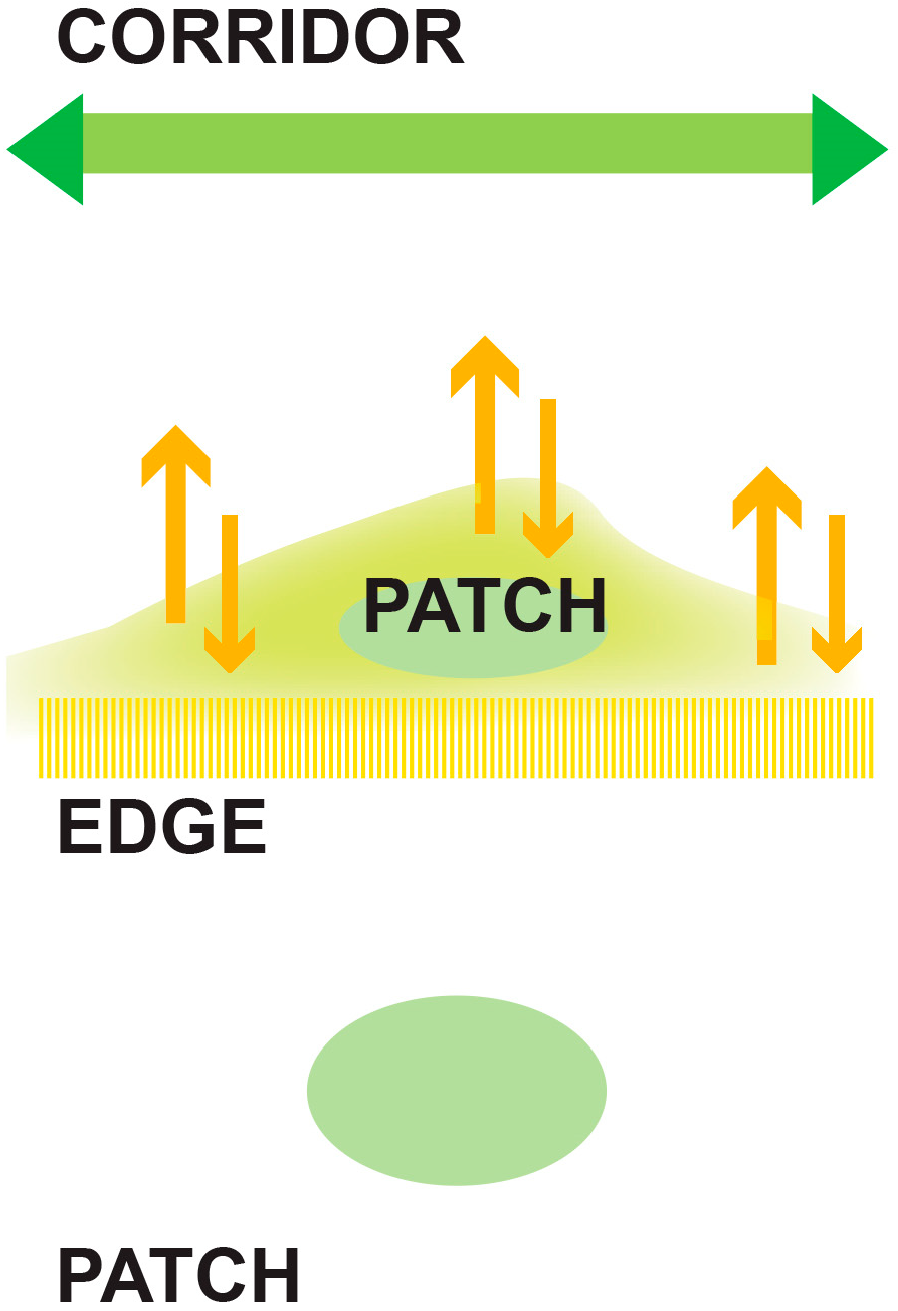
- Corridors, which signify the degree to which the landscape facilitates or impedes movements between resource patches. Corridors facilitate the movement of pollinators. Provision for the intensification of landscape corridors increases functional connectivity by allowing the development and exchange of material, energy, and food.
- (b)
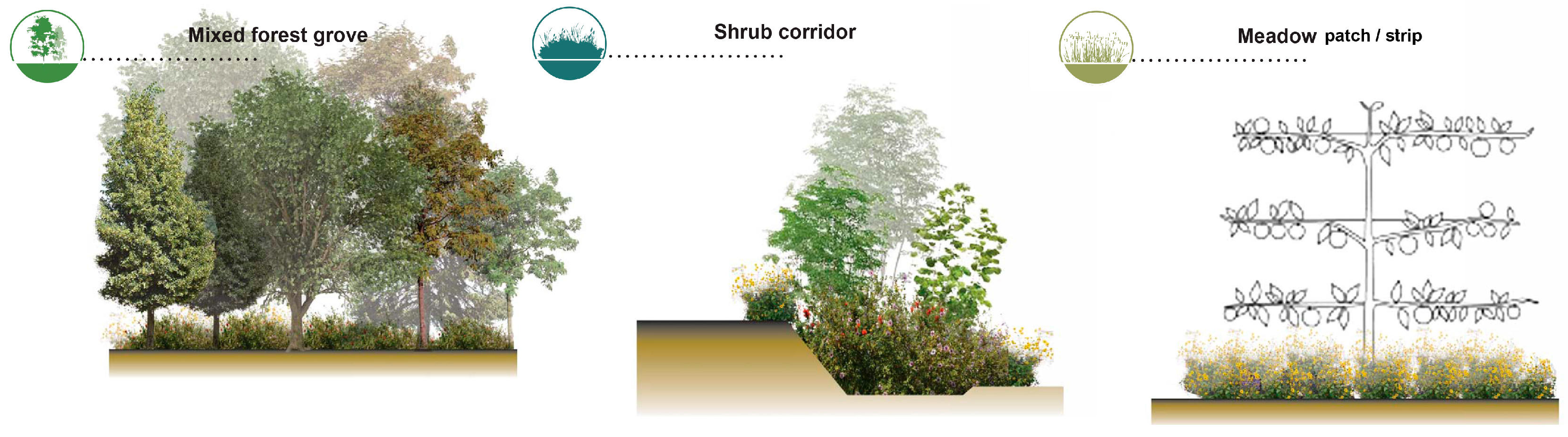
- Patches are defined as relatively homogeneous areas that differ from their surroundings. Their shape and size are critical for the spatial configuration of the landscape, since it disrupts or connects habitats.
- (c)
3. Results
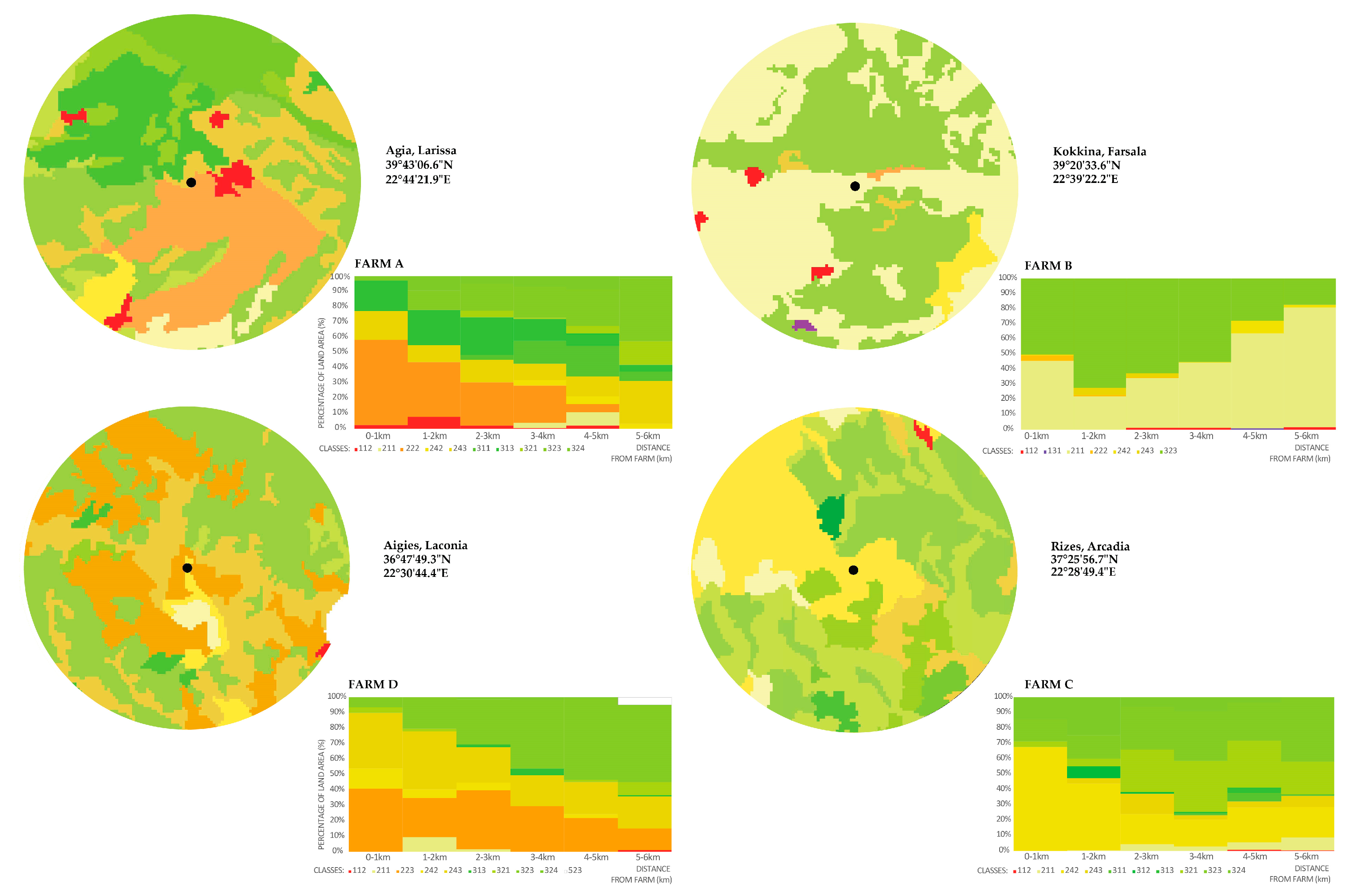
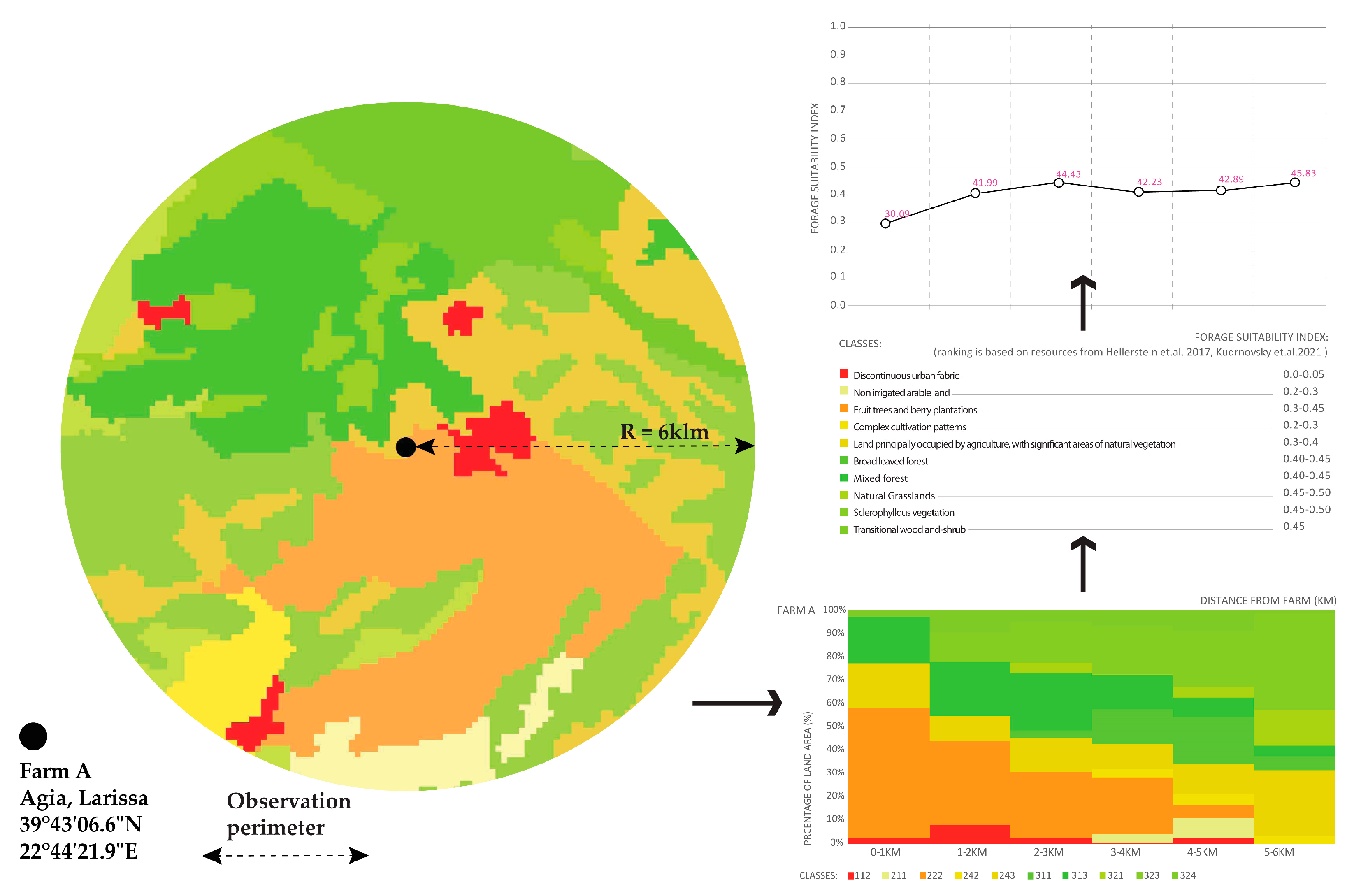
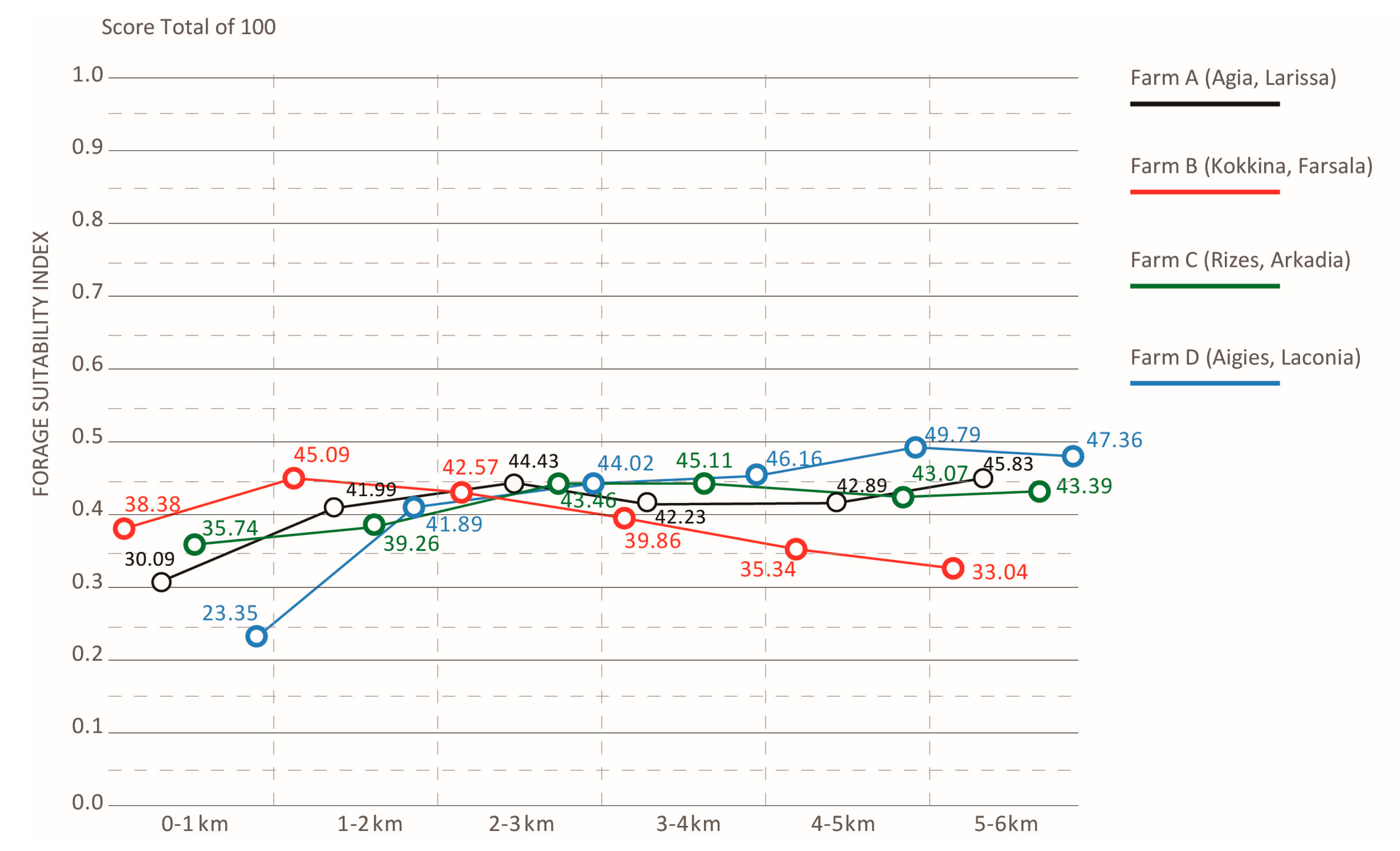
- Indexing foraging suitability and associating it with land cover classes from each farm helped us to understand the suitability of the context of each class for hovering pollinators. Within the range of up to 1 km from each farm, which that reaches the extent of foraging for native pollinators, farms A, B, and C seem to possess an adequate foraging profile variation from 0.3 to 0.38 as compared to farm D that has a moderate mean value of 0.23. For all study farms, the mean value of foraging suitability within the flight range of managed bees (0–6 km) ranges in adequate levels from 0.39 for farm C to 0.42 for farm D (Chart 4).
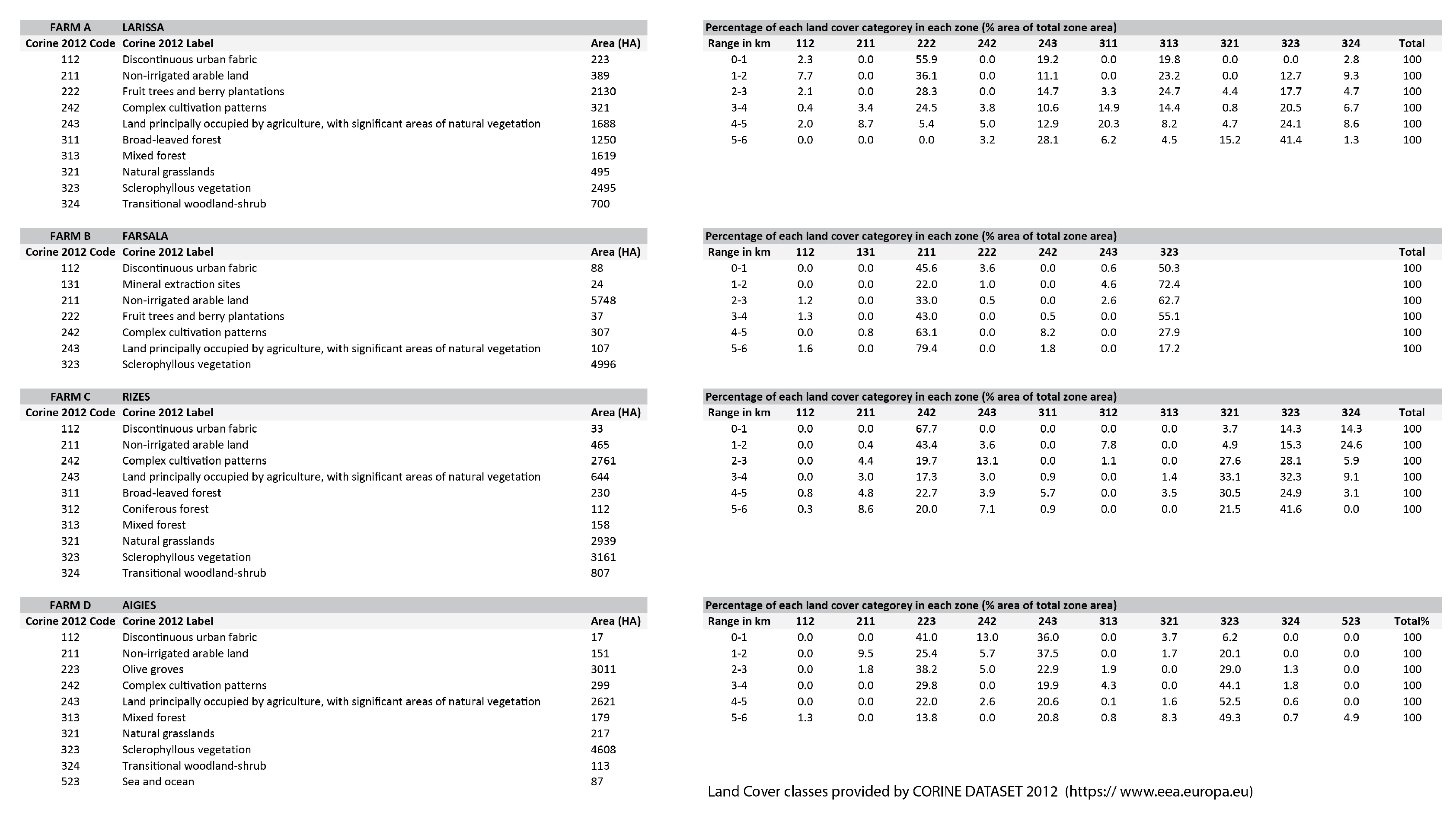
- By looking at the comparative change of land cover classes in farm A, the area remains highly agricultural. Almost 60% of the coverage within a 1 km distance from the farm is fruit trees and berry plantations. This cultivation diminishes significantly within 4–5 km from the farm. Mixed forest contributes to a quarter of the land coverage for a distance of up to 3 km from the farm and later changes to sclerophyllous scrub vegetation and transitional woodland-shrub communities, which are the dominant varieties at a 6 km distance from the farm, improving significantly the foraging conditions for the bees. In farm B, non-irrigated arable land almost doubles in coverage as we move away from the center, while sclerophyllous vegetation remains the most dominant community within and up to a 4 km distance from the farm. The land cover change in farm C revealed that almost 70% of the coverage within the first 1 km from the farm comprises complex cultivation patterns, such as viniculture and land principally occupied by agriculture, with significant areas of natural vegetation. However, natural grasslands and sclerophyllous vegetation gradually rise to half of the land coverage up to a 3 km distance and continue to dominate almost two-thirds of the area up to 6 km. Coniferous forest and transitional woodland-shrub areas comprise one-third of the land coverage within the first 2 km distance from the farm. Finally, in farm D, almost 70% of the coverage within the first 1 km from the farm comprises land principally occupied by olive grove agriculture. This land cover class diminishes to half of the coverage, gradually giving place to Sclerophyllous vegetation. However, field observations have shown that this land cover class has been seriously affected by recent wildfires.
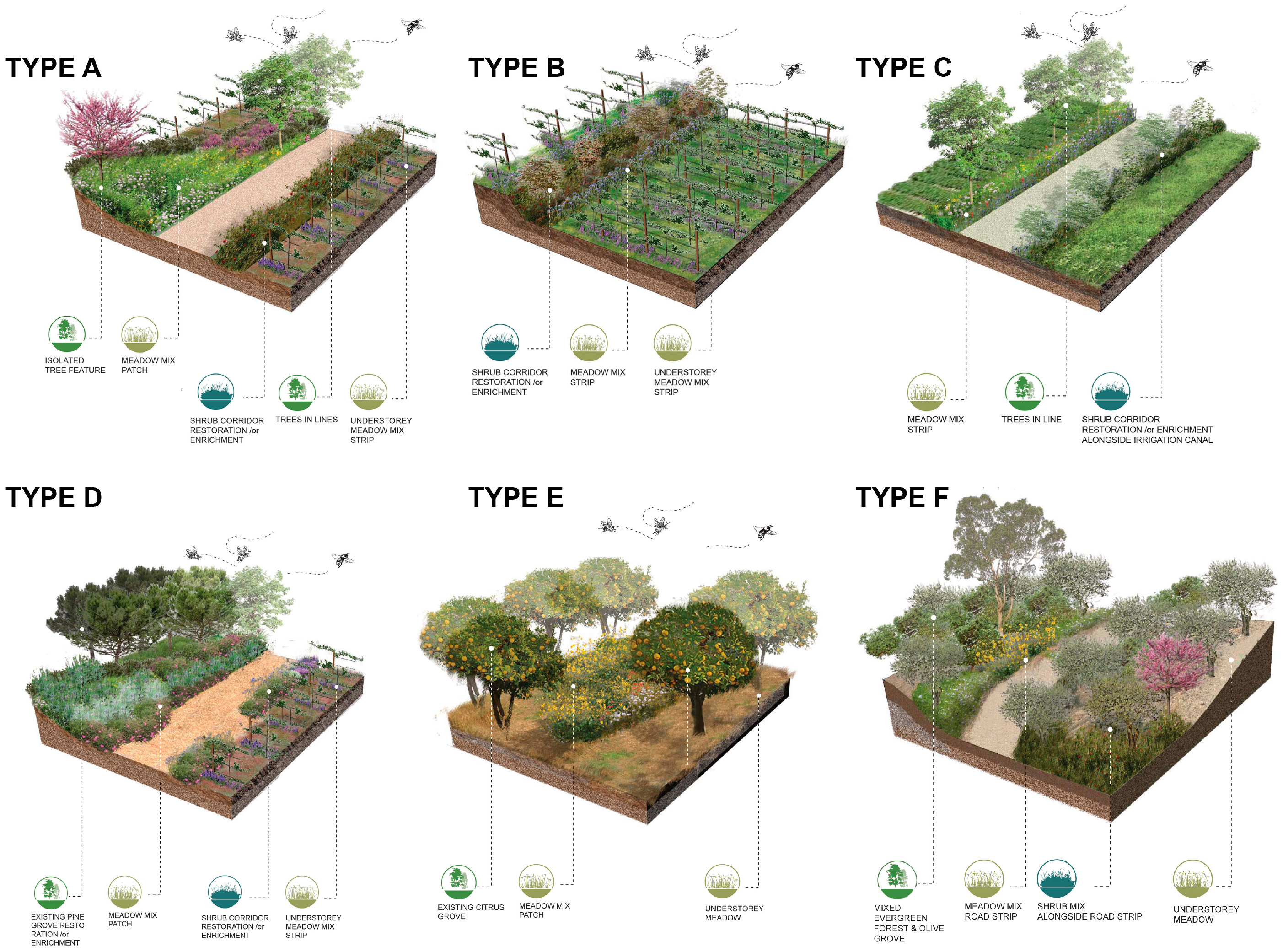
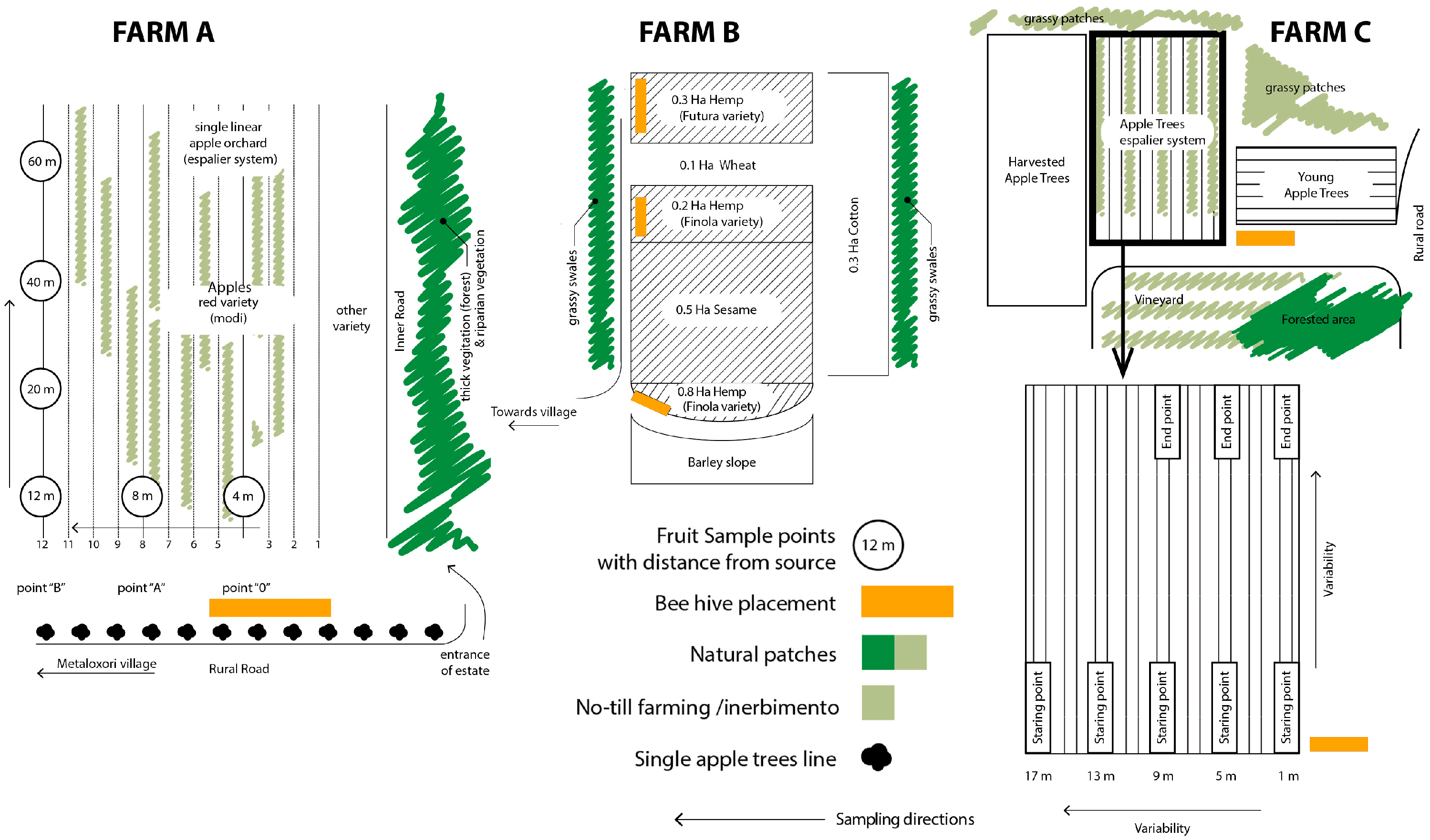
- A pilot test implementing managed bees in the farms (Scheme 2) delivered the following observations: In farm A, the applied conservation scheme included conservation of shrub corridors between different farming plots (type B model) and grassy meadow strips between rows of espaliered apples and cherries (partial implementation of landscape model type A). Honeybees were placed near the apple orchard of the Modi variety. Measurements were performed in late August, before the harvest of the variety. The main sampling method estimated the mean diameter every 10 fruits at the beginning of the rows as a function of the distance from the hives (at 5–15 and 29 m and so on). We observed an almost negative correlation: fruits appear larger when further away from the hive than when closer to it. However, this difference was not statistically significant (<5%); see Table 2.
- In farm B, no landscape model actions were followed. The installation of beehives in hemp plots presented a negative impact on hemp cultivation, as bees visited mail cannabis flowers to forage pollen and showed a preference for grazing on sesame cultivated in proximity to the hemp fields. Sesame proved to be an antagonist to hemp, as it has large white inflorescences and a longer flowering period than hemp. In farm C, type D and B landscape conservation models were partially applied. Measurements in the apple orchard indicated that there is a clear tendency for larger and heavier fruits the closer we are to the hives. A relatively small difference in the diameter of the fruit means a much larger increase in the total volume and therefore the weight of the fruit. So, the installation of bees gave from 8% (min.) to +12% (max.) larger fruits, corresponding to 32% (min. to 51% (max.) heavier fruits, making a significant impact on the production/harvest (yield) as shown in Table 3.
- If these data are juxtaposed to the foraging suitability profile of each plot, then farm A and farm C with comparable crops and foraging suitability rankings delivered different results, not because of the landscape models in the inter-habitat matrix of the farm system, but because the treatment of the farmers in farm A varied to that in farm C. Owners in farm A declared the use of chemical plant growth regulators, an external parameter, that deprived bees of their ecological service operations. In farm B, it was evident that competitive crops in the surrounding areas make the symbiotic profile of bees and farmers more complicated.
- A cross-relational observation of the results acquired from the different tiers of analysis revealed that farm A would benefit from edge patches on the west side of the plot and the conservation of wild corridors between the terraces with the apples in espaliers. For farm B, large patchiness of orchards and croplands interrupts forested areas, necessitating the reconstruction of wild corridors running from north to south to interconnect naturally vegetated patches. In farm C, landscape configuration suggests that there are adequate corridors with good flow of energy and mass between habitats westward and south of the farm. Provisions for edge patches and minor corridor strips embedded with the espalier systems could further foster or regulate connectivity locally between different cultivated patches. In farm D, large patchiness of orchards and olive cultivations requires interspersion of forested patches at the edge of olive groves for the enrichment of deforested areas. Distinct meadow patches within the orchards will increase biodiversity and provide pools for pollen and nectar resources within citrus monocultures.
4. Discussion
5. Conclusions
- (i)
- Beekeepers can acquire new locations for the placement of their hives, with the possibility of bees to forage and nest in low-competition areas [71], exploiting less pristine natural areas on which wild pollinators depend.
- (ii)
- Farmers can enjoy enhanced yield from the presence of managed bees.
- (iii)
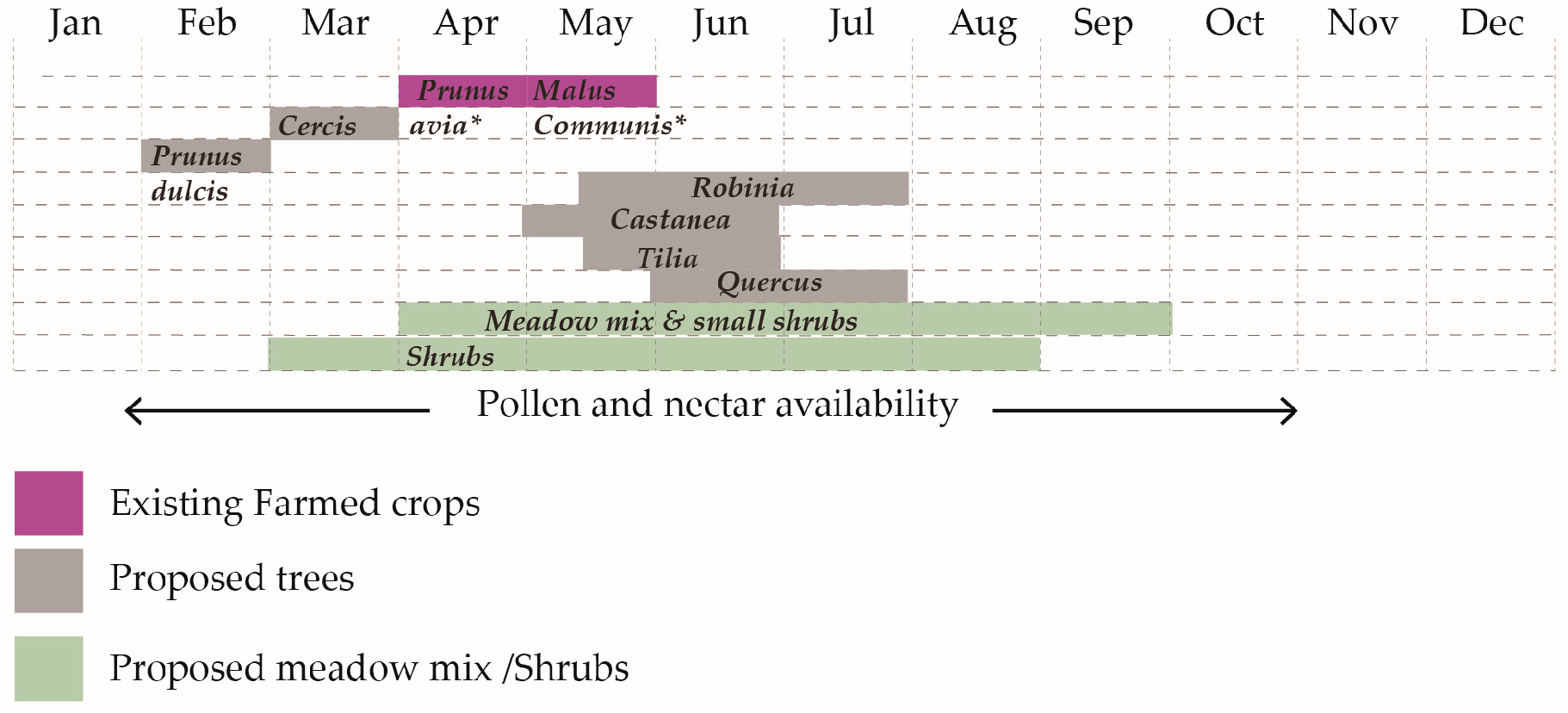
- Landscape models through a balanced approach of introduced floral patches and reserved semi-natural corridors and patches can offer extended resources of pollen and nectar, assuming the possibility to have winter hives in one location instead of constantly moving hives in different blossoming regions. This contributes to a circular model of agriculture, closing nutrient loops for bees, thus reducing dependencies on external inputs [8].
- (iv)
- Intensifying floral richness through design in a single-crop farm that hosts beehives can offer extended resources of pollen and nectar, minimizing the competition for resources amongst wild pollinators and honeybees.
- (v)
- The foraging index could be introduced in technological applications as a monitoring indicator of the larger foraging capacity of a specific hive, helping beekeepers assess the larger geographical context of an agricultural site and determine locations for establishment based on the bees’ dependence on nutrition parameters.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Etxegarai-Legarreta, O.; Sanchez-Famoso, V. The Role of Beekeeping in the Generation of Goods and Services: The Interrelation between Environmental, Socioeconomic, and Sociocultural Utilities. Agriculture 2022, 12, 551. [Google Scholar] [CrossRef]
- Patel, V.; Pauli, N.; Biggs, E.; Barbour, L.; Boruff, B. Why bees are critical for achieving sustainable development. Ambio 2021, 50, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Newton, P.; Civita, N.; Frankel-Goldwater, L.; Bartel, K.; Johns, C. What Is Regenerative Agriculture? A Review of Scholar and Practitioner Definitions Based on Processes and Outcomes. Front. Sustain. Food Syst. 2020, 4, 577723. [Google Scholar] [CrossRef]
- Francis, C.; Harwood, R.; Parr, J. The potential for regenerative agriculture in the developing world. Am. J. Altern. Agric. 1986, 1, 65–74. [Google Scholar] [CrossRef]
- Bullock, J.M.; McCracken, M.E.; Bowes, J.M.; Chapman, R.E.; Graves, R.A.; Hinsley, A.S.; Hutchins, G.M.; Nowakowski, M.; Nicholls, D.J.E.; Oakley, S. Does agri-environmental management enhance biodiversity and multiple ecosystem services?: A farm-scale experiment. Agric. Ecosyst. Environ. 2021, 320, 107582. [Google Scholar] [CrossRef]
- Constantin, M.; Strat, G.; Deaconu, M.E.; Patarlageanu, S.R. Innovative Agri-food Value Chain Management through a Unique Urban Ecosystem. Manag. Res. Pract. 2021, 13, 5–22. Available online: https://mrp.ase.ro/no133/f1.pdf (accessed on 27 December 2022).
- Van der Sluijs, J.P.; Vaage, N.S. Pollinators and Global Food Security: The Need for Holistic. Food Ethics 2016, 1, 75–91. [Google Scholar] [CrossRef]
- Rauw, W.M.; Gomez-Raya, L.; Star, L.; Overland, M.; Delezie, E.; Grivins, M.; Hamann, K.T.; Pietropaoli, M.; Klaassen, M.T.; Klemetsdal, G.; et al. Sustainable development in circular agriculture: An illustrative bee↺legume↺poultry example. Sustain. Dev. 2022. [Google Scholar] [CrossRef]
- UN General Assembly. Transforming Our World: The 2030 Agenda for Sustainable Development; 2015, A/RES/70/1. Available online: https://www.refworld.org/docid/57b6e3e44.html (accessed on 17 January 2023).
- European Commission, Directorate-General for Environment. The EU Pollinator’s Initiative; Publications Office: Luxembourg, 2019. Available online: https://op.europa.eu/en/publication-detail/-/publication/8e6e7bf0-7d03-11e9-9f05-01aa75ed71a1 (accessed on 17 January 2023).
- European Commission, Directorate-General for Communication. European Green Deal: Delivering on Our Targets; Publications Office of the European Union: Luxembourg, 2021; Available online: https://data.europa.eu/doi/10.2775/373022 (accessed on 17 January 2023).
- Muir, C. Re-Designing Farming. Foreground 2021. Available online: https://www.foreground.com.au/agriculture-environment/re-designing-farming/ (accessed on 17 January 2023).
- Lowicki, D.; Fagiewicz, K. A new model of pollination services potential using a landscape approach: A case study of post-mining area in Poland. Ecosyst. Serv. 2021, 52, 101370. [Google Scholar] [CrossRef]
- Albrecht, M.; Knecth, A.; Riesen, M.; Rutz, T.; Ganser, D. Time since establishment drives bee and hoverfly diversity, abundance of crop-pollinating bees and aphidophagous hoverflies in perennial wildflower. Basic Appl. Ecol. 2021, 57, 102–114. [Google Scholar] [CrossRef]
- Altieri, M.A. Agroecology: A new research and development paradigm for world agriculture. Agric. Ecosyst. Environ. 1989, 27, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Martinez, C.; Gonzalez-Estévez, M.A.; Cursach, J.; Lazaro, A. Pollinator richness, pollination networks, and diet adjustment along local and landscape gradients of resource diversity. Ecol. Appl. 2022, 32, e2634. [Google Scholar] [CrossRef] [PubMed]
- Beyer, N.; Gabriel, D.; Westphal, C. Landscape composition modifies pollinator densities, foraging behavior and yield formation in faba beans. Basic Appl. Ecol. 2022, 61, 30–40. [Google Scholar] [CrossRef]
- Ekroos, J.; Jakobsson, A.; Wideen, J.; Herbertsson, L.; Rundlöf, M.; Smith, H.G. Effects of landscape composition and configuration on pollination in a native herb: A field experiment. Oecologia 2015, 179, 509–518. [Google Scholar] [CrossRef]
- Banaszak-Cibicka, W.; Ratyńska, H.; Dylewski, Ł. Features of urban green space favourable for large and diverse bee populations (Hymenoptera: Apoidea: Apiformes). Urban For. Urban Green. 2016, 20, 448–452. [Google Scholar] [CrossRef]
- Betts, M.G.; Hadley, A.; Kormann, U. The landscape ecology of pollination. Landsc. Ecol. 2019, 34, 961–966. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kaloveloni, A.; Petanidou, T. Assessing Climate Change Impacts on Island Bees: The Aegean Archipelago. Biology 2022, 11, 552. [Google Scholar] [CrossRef]
- Ke, Q.; Chen, S.; Zhao, D.; Li, M.; Su, C. Effects of Land-Use Change on the Pollination Services for Litchi and Longan Orchards: A Case Study of Huizhou, China. Land 2022, 11, 1073. [Google Scholar] [CrossRef]
- De Palma, A.; Abrahamczyk, S.; Aizen, M.A.; Albrecht, M.; Basset, Y.; Bates, A.; Blake, R.J.; Boutin, C.; Bugter, R.; Connop, S.; et al. Predicting bee community responses to land-use changes: Effects of geographic and taxonomic biases. Sci Rep. 2016, 6, 31153. [Google Scholar] [CrossRef]
- Samuelson, A.E.; Leadbeater, E. A land classification protocol for pollinator ecology research: An urbanization case study. Ecol Evol. 2018, 8, 5598–5610. [Google Scholar] [CrossRef]
- Hellerstein, D.; Hitaj, C.; Smith, D.; Davis, A. Land Use, Land Cover, and Pollinator Health: A Review and Trend Analysis. In United States Department of Agriculture, Economic Research Service; Report no 232; U.S. Department of Agriculture, Economic Research Service: Washington, DC, USA, 2017. Available online: https://www.researchgate.net/publication/319130660_Land_Use_Land_Cover_and_Pollinator_Health_A_Review_and_Trend_Analysis (accessed on 27 December 2022).
- Nicholls, C.I.; Altieri, M.A. Agro-ecological approaches to enhance resilience. Farming Matters 2012, 14–17. Available online: https://archive.foodfirst.org/wp-content/uploads/2016/02/Farming-Matters-28-2-Agroecological-approaches-to-enhance-resilience.pdf (accessed on 27 December 2022).
- Holland, J.M.; Smith, B.M.; Storkey, J.; Lutman, P.J.W.; Aebischer, N.J. Managing habitats on English farmland for insect pollinator conservation. Biol. Conserv. 2015, 182, 215–222. [Google Scholar] [CrossRef]
- O’Donoghue, T.; Minasny, B.; Mc Bratney, A. Regenerative Agriculture and Its Potential to Improve Farmscape Function. Sustainability 2022, 14, 5815. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Klein, A.M.; Tscharntke, T. Spatiotemporal variation in the diversity of hymenoptera across a tropical habitat gradient. Ecology 2005, 86, 3296–3302. [Google Scholar] [CrossRef]
- Marnasidis, S.; Kantartzis, A.; Malesios, C.; Hatjina, F.; Arabatzis, G.; Verikouki, E. Mapping Priority Areas for Apiculture Development with the Use of Geographical Information Systems. Agriculture 2021, 11, 182. [Google Scholar] [CrossRef]
- Cole, L.J.; Kleijn, D.; Dicks, L.V.; Stout, J.C.; Potts, S.G.; Albrecht, M.; Balzan, M.V.; Bartomeus, I.; Bebeli, P.J.; Bevk, D.; et al. A critical analysis of the potential for EU Common Agricultural Policy measures to support wild pollinators on farmland. J. Appl. Ecol. 2020, 57, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, I. Permaculture, regeneration and AAFMs discussion shift. In Proceedings of the XXVII European Society for Rural Sociology Congress, Krakow, Poland, 24–27 July 2017. [Google Scholar]
- Ngugi, R.K.; Nyariki, D.M. Rural livelihoods in the arid and semi-arid environments of Kenya: Sustainable alternatives and challenges. Agric. Hum. 2005, 22, 65–71. [Google Scholar] [CrossRef]
- López-Pintor, A.; Sanz-Cañada, J.; Salas, E.; Rescia, A.J. Assessment of Agri-Environmental Externalities in Spanish Socio-Ecological Landscapes of Olive Groves. Sustainability 2018, 10, 2640. [Google Scholar] [CrossRef]
- Kevan, P.G.; Phillips, T.P. The economic impacts of pollinator declines: An approach to assessing the consequences. Conserv. Ecol. 2001, 5, 8. Available online: http://www.consecol.org/vol5/iss1/art8/ (accessed on 27 December 2022). [CrossRef]
- Hanley, N.; Breeze, D.T.; Ellis, C.; Goulson, D. Measuring the economic value of pollination services: Principles, evidence and knowledge gaps. Ecosyst. Serv. 2015, 14, 124–132. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Pollinator Partnership. Available online: http://pollinator.org (accessed on 27 December 2022).
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Rizzardo, R.; Milfont, M.; Da Silva, E.M.S.; Freitas, B.M. Apis mellifera pollination improves agronomic productivity of anemophilous castor bean (Ricinus communis). Ann. Braz. Acad. Sci. 2012, 84, 1137–1145. Available online: https://www.scielo.br/j/aabc/a/KZ7QXcsp6jpkkVgT8P4nzKv/?format=pdf&lang=en (accessed on 27 December 2022). [CrossRef] [PubMed]
- Lautenbach, S.; Seppelt, R.; Liebscher, J.; Dormann, C.F. Spatial and Temporal Trends of Global Pollination Benefit. PLoS ONE 2012, 7, e35954. [Google Scholar] [CrossRef] [PubMed]
- Gilpin, A.M.; O’Brien, C.; Kobel, C.; Brettell, L.E.; Cook, J.M.; Power, S.A. Co-flowering plants support diverse pollinator populations and facilitate pollinator visitation to sweet cherry crops. Basic Appl. Ecol. 2022, 63, 36–48. [Google Scholar] [CrossRef]
- Földesi, R.; Kovács-Hostyánszki, A.; Korosi, A.; Somay, L.; Elek, Z.; Marko, V.; Sarospataki, M.; Bakos, R.; Varga, A.; Nyisztor, K.; et al. Relationships between wild bees, hoverflies and pollination success in apple orchards with different landscape contexts. Agric. For. Entomol. 2015, 18, 68–75. [Google Scholar] [CrossRef]
- The Guardian, BEES are Worth Billions to Farmers across the Globe, Study Suggests, 2015. Available online: https://www.theguardian.com/environment/2015/jun/17/bees-are-worth-billions-tofarmers-across-the-globe-study-suggests (accessed on 27 December 2022).
- Winfree, R.; Griswold, T.; Kremen, C. Effect of Human Disturbance on Bee Communities in a Forested Ecosystem. Conserv. Biol. 2007, 21, 213–223. [Google Scholar] [CrossRef]
- Bernhardt, C.E.; Mitchell, R.J.; Michaels, H.J. Effects of Population Size and Density on Pollinator Visitation, Pollinator Behavior, and Pollen Tube Abundance in Lupinus Perennis. Int. J. Pla 2008, 169, 944–953. [Google Scholar] [CrossRef]
- Petanidou, T.; Kallimanis, A.S.; Lazarina, M.; Tscheulin, T.; Devalez, J.; Stefanaki, A.; Hanlidou, E.; Vujic, A.; Kaloveloni, A.; Sgardelis, S.P. Climate drives plant–pollinator interactions even along small-scale climate gradients: The case of the Aegean. Plant Biol. 2018, 20, 176–183. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; Dombeck, E.; Gerber, J.; Gerber, J.; Knuth, K.A.; Mueller, N.D.; Mueller, M.; Ziv, G.; Klein, A.M. Global malnutrition overlaps with pollinator-dependent micronutrient production. Proc. R. Soc. B 2014, 281, 20141799. [Google Scholar] [CrossRef]
- Piri Sahragard, H.; Ajorlo, M.; Karami, P. Landscape structure and suitable habitat analysis for effective restoration planning in semi-arid mountain forests. Ecol. Process. 2021, 10, 17. [Google Scholar] [CrossRef]
- Allen-Wardell, G.; Bernhardt, P.; Bitner, R.; Burquez, A.; Buchmann, S.; Cane, J.; Cox, P.A.; Dalton, V.; Feinsinger, P.; Ingram, M.; et al. The potential consequences of pollinator declines on the conservation of biodiversity and stability of food crop yields. Conserv. Biol. 1998, 12, 8–17. [Google Scholar] [CrossRef] [Green Version]
- EFTEC. The Economic, Social and Ecological Value of Ecosystem Services: A Literature Review; Department for Environment, Food and Rural Affairs (DEFRA), Economics for the Environment Consultancy: London, UK, 2005. Available online: https://www.cbd.int/financial/values/unitedkingdom-valueliterature.pdf (accessed on 27 December 2022).
- Douglas, A. Landis, Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 2017, 18, 1–12. [Google Scholar] [CrossRef]
- Ballogh, A. The rise and fall of monoculture farming. Horizon 2021. Available online: https://ec.europa.eu/research-and-innovation/en/horizon-magazine/rise-and-fall-monoculture-farming (accessed on 27 December 2022).
- Honey Bee Health Coalition. Available online: https://honeybeehealthcoalition.org/program/bee-integrated-demonstration-project (accessed on 27 December 2022).
- Martinez-Núñez, C.; Manzaneda, A.J.; Isla, J.; Tarifa, R.; Calvo, G.; Molina, J.L.; Salido, T.; Ruiz, C.; Gutierrez, J.E.; Rey, P.J. Low-intensity management benefits solitary bees in olive groves. J. Appl. Ecol. 2020, 57, 111–120. [Google Scholar] [CrossRef]
- Olives Alive. Available online: https://olivaresvivos.com/en/ (accessed on 27 December 2022).
- Wezel, A.; Casagrande, M.; Celette, F.; Vian, J.-F.; Ferrer, A.; Peigné, J. Agroecological practices for sustainable agriculture. A review. Agron. Sustain. Dev. 2014, 34, 1–20. [Google Scholar] [CrossRef]
- Costantza, R.; D’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, k.; Naeem, S.; O’ Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capita. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Herrera, C.M. Gradual replacement of wild bees by honeybees in flowers of the Mediterranean Basin over the last 50 years. Proc. R. Soc. B 2020, 287, 20192657. [Google Scholar] [CrossRef]
- Petanidou, T.; Lambor, E. A land for flowers and bees: Studying pollination ecology in Mediterranean communities. Plant Biosyst. 2005, 139, 279–294. [Google Scholar] [CrossRef]
- Hope, K. Honey: The Untapped Potential of a Greek Superfood, Economia 2022. Available online: https://www.economia.gr/en/honey-the-untapped-potential-of-a-greek-superfood/ (accessed on 27 December 2022).
- Makri, P.; Papanagiotou, P.; Papanagiotou, E. Efficiency And Economic Analysis Of Greek Beekeeping Farms. Bulg. J. Agric. Sci. 2015, 21, 479–484. Available online: https://www.agrojournal.org/21/03-02.pdf (accessed on 27 December 2022).
- Marchal, L.; Prager, A.; Villadiego, A. Beekeepers in Western Europe Fear for Their Future. Equal. Times 2022. Available online: https://www.equaltimes.org/beekeepers-in-western-europe-fear#.Y87lFD1Bypo (accessed on 27 December 2022).
- Panagos, P.; Ballabio, C.; Poesen, J.; Lugato, E.; Scarpa, S.; Montanarella, L.; Borrelli, P. A soil erosion indicator for supporting agricultural, environmental and climate policies in the European Union. Remote Sens. 2020, 12, 1365. [Google Scholar] [CrossRef]
- OECD. OECD Environmental Performance Reviews: Greece 2020; OECD Environmental Performance Reviews; OECD Publishing: Paris, France, 2020. [Google Scholar] [CrossRef]
- Gomiero, T. Soil Degradation, Land Scarcity and Food Security: Reviewing a Complex Challenge. Sustainability 2016, 8, 281. [Google Scholar] [CrossRef]
- Ferreira, C.S.S.; Seifollahi-Aghmiuni, S.; Destouni, G.; Ghajarnia, N.; Kalantari, Z. Soil degradation in the European Mediterranean region: Processes, status and consequences. Sci. Total Environ. 2022, 805, 150106. [Google Scholar] [CrossRef] [PubMed]
- Lister, N.; Brocki, M.; Ament, R. Integrated adaptive design for wildlife movement under climate change. Front. Ecol. Environ. 2015, 13, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Lázaro, A.; Müller, A.; Ebmer, A.W.; Dathe, H.H.; Scheuchl, E.; Schwarz, M.; Risch, S.; Pauly, A.; Devalez, J.; Tscheulin, T.; et al. Impacts of beekeeping on wild bee diversity and pollination networks in the Aegean Archipelago. Ecography 2021, 44, 1353–1365. [Google Scholar] [CrossRef]
- Casanelles-Abella, J.; Fontana, S.; Fournier, B.; Frey, D.; Moretti, M. Low Resource Availability Drives Feeding Niche Partitioning between Wild Bees and Honeybees in a European City. Ecol. Appl. 2023, 33, e2727. [Google Scholar] [CrossRef]
- Henry, M.; Rodet, G. Controlling the impact of the managed honeybee on wild bees in protected areas. Sci. Rep. 2018, 8, 9308. [Google Scholar] [CrossRef]
- Cannon, D. Is Beekeeping Sustainable? The Australian Beekeper. 2014. Available online: http://www.theabk.com.au/articles/2016/8/3/is-beekeeping-sustainable (accessed on 27 December 2022).
- Nicholson, C.C.; Ward, K.L.; Williams, N.M.; Isaacs, R.; Mason, K.S.; Wilson, J.K.; Brokaw, J.; Gut, L.J.; Rothwell, N.L.; Wood, T.J.; et al. Mismatched outcomes for biodiversity and ecosystem services: Testing the responses of crop pollinators and wild bee biodiversity to habitat enhancement. Ecol. Lett. 2020, 23, 326–335. [Google Scholar] [CrossRef]
- Zeunert, J. Landscape Architecture and Environmental Sustainability, 1st ed.; Bloomsbury Publishing: London, UK, 2017; pp. 40–60. [Google Scholar]
- Klein, A.; Cunningham, S.A.; Bos, M.; Steffan-Dewenter, I. Advances in Pollination Ecology from Tropical Plantation Crops. Ecology 2008, 89, 935–943. [Google Scholar] [CrossRef]
- Donoso, S.; Murúa, M. Floral Patches and Their Impact on Pollinator Attraction and Yield Production on Cucurbita maxima Var. Paine in Central Chile. Diversity 2021, 13, 608. [Google Scholar] [CrossRef]
- Zhou, Y.; Lambrides, C.J.; Li, J.; Xu, Q.; Toh, R.; Tian, S.; Yang, P.; Yang, H.; Ryder, M.; Denton, M.D. Nitrifying Microbes in the Rhizosphere of Perennial Grasses are Modified by Biological Nitrification Inhibition. Microorganisms 2020, 8, 1687. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef]
- Bennett, A.F.; Radford, J.Q.; Haslem, A. Properties of land mosaics: Implications for nature conservation in agricultural environments. Biol. Conserv. 2006, 133, 250–264. [Google Scholar] [CrossRef]
- Mac Nally, R.; Soderquist, T.; Tzaros, C. The conservation value of mesic gullies in dry forest landscapes: Avian assemblages in the box-ironbark ecosystem of southern Australia. Biol. Conserv. 2000, 93, 293–302. [Google Scholar] [CrossRef]
- Varah, A.; Jones, H.; Smith, J.; Smith, J.; Potts, S.J. Temperate agroforestry systems provide greater pollination service than monoculture. Agric. Ecosyst. Environ. 2020, 301, 107031. [Google Scholar] [CrossRef]
- Rands, S.A.; Whitney, H.M. Effects of pollinator density-dependent preferences on field margin visitations in the midst of agricultural monocultures: A modelling approach. Ecol. Model. 2010, 221, 1310–1316. [Google Scholar] [CrossRef]
- Walker, K.J.; Stevens, P.A.; Stevens, D.P.; Mountford, J.O.; Manchester, S.J.; Pywell, R. F The restoration and re-creation of species-rich lowland grassland on land formerly managed for intensive agriculture in the UK. Biol. Conserv. 2004, 119, 1–18. [Google Scholar] [CrossRef]
- Polus, E.; Vandewoestijne, S.; Choutt, J.; Baguette, M. Tracking the effects of one century of habitat loss and fragmentation on calcareous grassland butterfly communities. Biodivers. Conserv. 2007, 16, 3423–3436. [Google Scholar] [CrossRef]
- Hadley, A.; Betts, M. The effects of landscape fragmentation on pollination dynamics: Absence of evidence not evidence of absence. Biol. Rev. Camb. Philos. Soc. 2012, 87, 526–544. [Google Scholar] [CrossRef]
- Kudrnovsky, H.; Ellmauer, T. Report for a List of Annex I Habitat Types Important for Pollinators. 2020. Available online: https://www.eionet.europa.eu/etcs/etc-bd/products/etc-bd-reports/etc-bd-technical-paper-1-2020-report-for-a-list-of-annex-i-habitat-types-important-for-pollinators (accessed on 27 December 2022).
- Bartholomée, O.; Lavorel, S. Disentangling the diversity of definitions for the pollination ecosystem service and associated estimation methods. Ecol. Indic. 2019, 107, 105576. [Google Scholar] [CrossRef]
- Arianoutsou, M.; Delipetrou, P.; Dimopoulos, P.; Economidou, E.; Karagiannakidou, V.; Konstantinidis, P.; Panagiotidis, P.; Panitsa, M.; Tsiourlis, G. Habitat types in Greece. In Directive 92/43/EEC, The Project of Habitats in Greece, Network Natura 2000; Dafis, S., Papastergiadou, E., Georgiou, K., Babalonas, D., Georgiadis, T., Papageorgiou, M., Lazaridou, T., Tsiaousi, V., Eds.; Goulandris Museum of Natural History-Greek Center of Biotopes Ecotypes: Thessaloniki, Greek, 1997; pp. 402–435. [Google Scholar]
- FILOTIS. Database for the Natural Environment of Greece. Available online: http://filotis.itia.ntua.gr (accessed on 27 December 2022).
- Beekman, M.; Ratnieks, F.L.W. Long-range foraging by the honey-bee, Apis mellifera L. Funct. Ecol. 2000, 14, 490–496. [Google Scholar] [CrossRef]
- Copernicus, Land Monitoring Service. Available online: https://land.copernicus.eu/pan-european/corine-land-cover (accessed on 27 December 2022).
- Davies, C.E.; Moss, D.; Hill, M.O. EUNIS Habitat Classification Revised. European Topic Centre on Nature Protection and Biodiversity. 2004. Available online: https://www.eea.europa.eu/data-and-maps/data/eunis-habitat-classification/documentation/eunis-2004-report.pdf/download (accessed on 27 December 2022).
- Ricketts, T.H.J.; Regetz, I.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S.S.; Klein, A.M.; Mayfield, M.M.; et al. Landscape Effects on Crop Pollination Services: Are There General Patterns? Ecol. Lett. 2008, 11, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, L.A.I.; Steffan-Dewenter, C.; Kremen, C.; Morales, J.M.; Bommarco, R.; Cunningham, S.A.; Carvalheiro, L.G.; Chacoff, N.P.; Dudenhöffer, J.H.; Greenleaf, S.S.; et al. Stability of Pollination Services Decreases With Isolation From Natural Areas Despite Honey Bee Visits. Ecol. Lett. 2011, 14, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Forman, R.T.T. Land Mosaics: The Ecology of Landscapes and Regions, 1st ed.; Cambridge University Press: Cambridge, UK, 1995; pp. 406–474. [Google Scholar]
- Ogilvie, J.E.; Thomson, J.D. Site fidelity by bees drives pollination facilitation in sequentially blooming plant species. Ecology 2016, 97, 1442–1451. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef]
- Dimou, M.; Tananaki, C.; Liolios, V.; Thrasyvoulou, A. Pollen Foraging by Honey Bees (Apis mellifera L.) in Greece: Botanical and Geographical Origin. J. Apic. Sci. 2014, 58, 11–23. [Google Scholar] [CrossRef] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Farm A | Farm B | Farm C | Farm D |
|---|---|---|---|---|
| Place, regional unit geo-reference location of farm | Metaxochori, Larissa (39°43′06.6″ N, 22°44′21.9″ E) | Kokkina, Farsala (39°20′33.6″ N, 22°39′22.2″ E | Rizes, Arkadia (37°25′56.7″ N, 22°28′49.4″ E | Aigies, Laconia (39°20′33.6″ N, 22°39′22.2″ E |
| Climate data during farm-blooming season * Temperature variation/rainy days | March–May 10–20 °C * | July and September 25–32 °C * | March–May 15–23 °C * | February–May 25–32 °C * |
| 825 mm */20 days * | 640 mm */14 days * | 764 mm */18 days * | 534 mm */11 days * | |
| Altitude/slope | 250 m/gradient 3.5% | 300 m/plain | 700 m/gradient 3–5% | 100 m/Hills |
| Soil | Rich in organic matter, medium–shallow ground depth, fertile, average texture | Medium depth, clay-loam type, organic matter in moderate-to-low levels | Medium–shallow ground depth, clay-loam type, fertile, calcium deficiency | Medium–shallow ground depth, clay-loam type, fertile, excess calcium |
| Bio-climatic environment Vegetation zones | Zone Quercetalia ilicis Subarea Quercion ilicis | Zone Quercetalia ilicis Subarea Quercion ilicis | Zone Quercetalia pubescentis Subarea Ostryo carpinion | Zone Quercetalia ilicis subarea Oleo ceratonion growth Oleo ceratonietum |
| Cultivations, varieties | Apple varieties: Firikia, Gala, Jonas, Pink Lady; pear; cherries | Sesame, durum wheat, hemp | Gala apples, Granny Smith apples, Jeromine apples, Super Starkin apples | Olive trees, citrus fruit, lemon trees, tangerine, Valencia oranges, oranges |
| Plot size and cultivation type and crop details, age of trees in years (y) | 5.5 ha, not certified organic, irrigated, 1000 trees/ha, espaliered, apples: 5–45 y | 12 ha, organic, irrigated, 200 plants/m2 | 1.5–2.0 ha organic, irrigated, espaliered, 1200–1300 trees/ha, apples: 5–6 y | 20 ha, organic, irrigated, 40 trees/ha, >35 y |
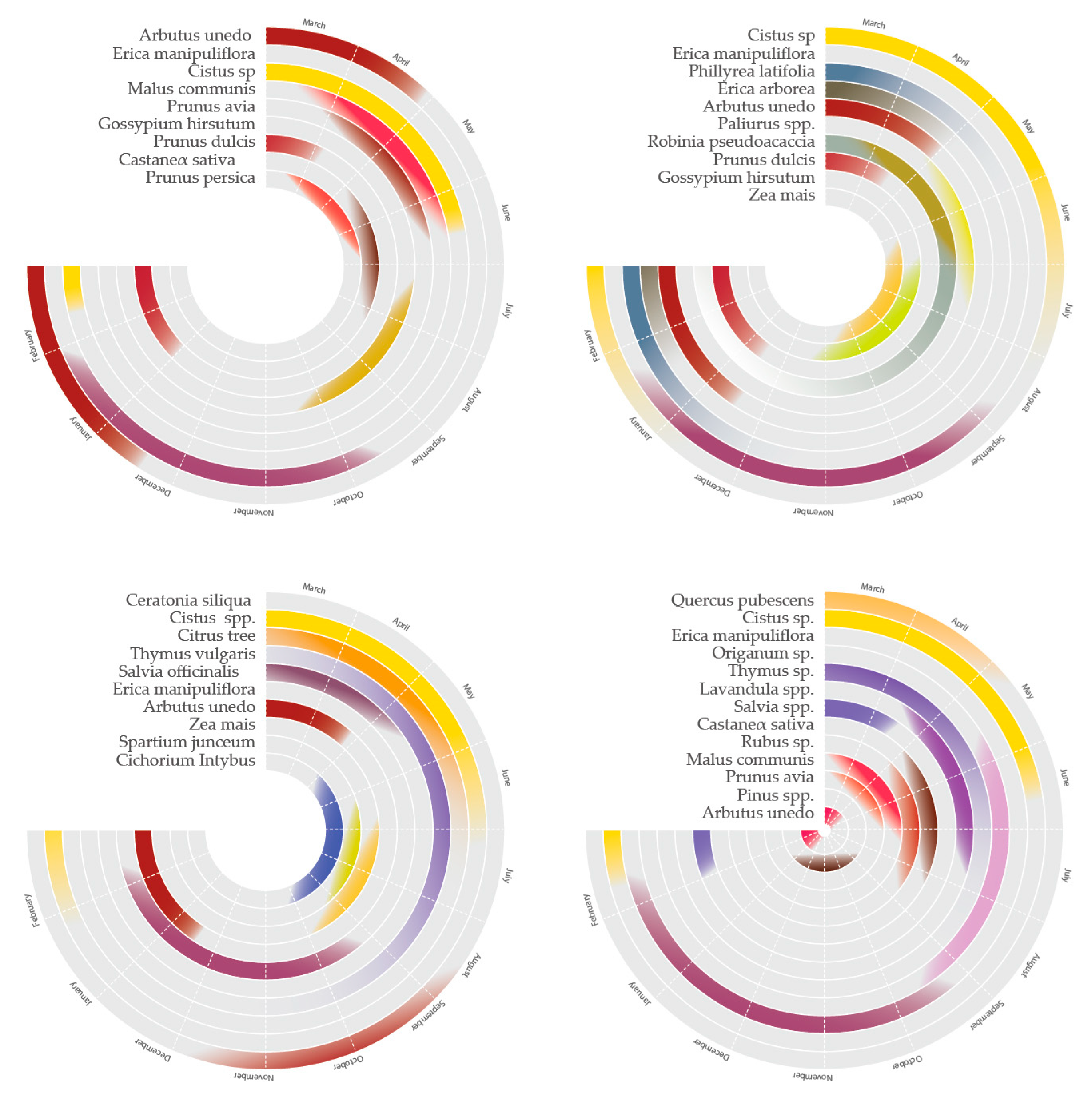
| Significant plant species observed within 6 km suitable for bees (data based on the literature, listed nectar- and pollen-producing species) | Arbutus unedo, Erica manipuliflora, Cistus sp., Malus communis, Prunus avia, Gossypium hirsutum, Prunus dulcis, Castanea sativa, Prunus persica | Cistus sp., Erica manipuliflora, Phillyrea latifolia, Erica arborea, Arbutus unedo, Paliurus spp., Robinia pseudoacaccia, Prunus dulcis, Gossypium hirsutum, Zea mais | Quercus pubescens, Cistus sp., Erica manipuliflora, Origanum sp., Thymus sp., Salvia spp., Castanea sativa, Rubus sp., Malus communis, Prunus avia, Pinus spp., Arbutus unedo | Ceratonia siliqua, Cistus spp., citrus trees, Thymus vulgaris, Salvia officinalis Ericamanipuliflora, Arbutus unedo, Zea mais |
| Point of Sampling (at 3rd Row from Interior Row) | Distance from Beehives (m) | Mean Diameter Every 10 Fruits (mm) |
|---|---|---|
| Point «0»—4th row | 5 | 69.4 |
| Point «A»—8th row | 15 | 69.9 |
| Point «B»—12th row | 29 | 72.4 |
| Point of sampling (at 12rd row from interior row) | ||
| Point «B» [3rd tree] | 72.4 | |
| Point «Β + 20 m» | 71.25 | |
| Point «Β + 40 m» | 73.1 | |
| Point «Β + 60 m» | 71.5 |
| Point of Sampling | Weight (Mean) g | Mean Diameter Every 10 Fruits (mm) |
|---|---|---|
| 1st row—starting point | 228 | 78.4 |
| 1st row—end point | 221 | 76.3 |
| 5th row—starting point | 260 | 81.1 |
| 5th row—end point | 193 | 75.8 |
| 13th row—starting point | 172 | 72.4 |
| 13th row—end point | 203 | 77.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carydi, I.; Koutsianas, A.; Desyllas, M. People, Crops, and Bee Farming: Landscape Models for a Symbiotic Network in Greece. Land 2023, 12, 430. https://doi.org/10.3390/land12020430
Carydi I, Koutsianas A, Desyllas M. People, Crops, and Bee Farming: Landscape Models for a Symbiotic Network in Greece. Land. 2023; 12(2):430. https://doi.org/10.3390/land12020430
Chicago/Turabian StyleCarydi, Io, Athanasios Koutsianas, and Marios Desyllas. 2023. "People, Crops, and Bee Farming: Landscape Models for a Symbiotic Network in Greece" Land 12, no. 2: 430. https://doi.org/10.3390/land12020430