
Landscape and Stand Characteristics Influence on the Bird Assemblage in Nothofagus antarctica Forests of Tierra del Fuego

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
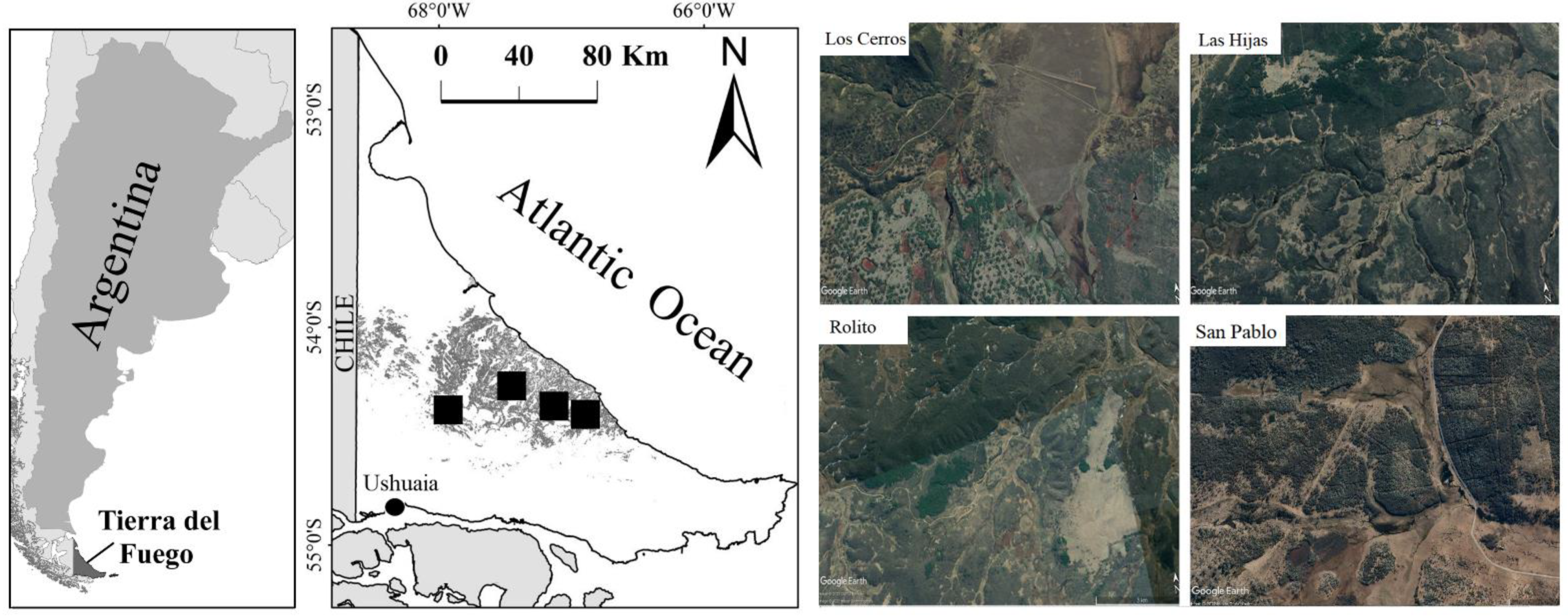
2.1. Study Area
2.2. Sampling Design
2.3. Bird Sampling Methodology
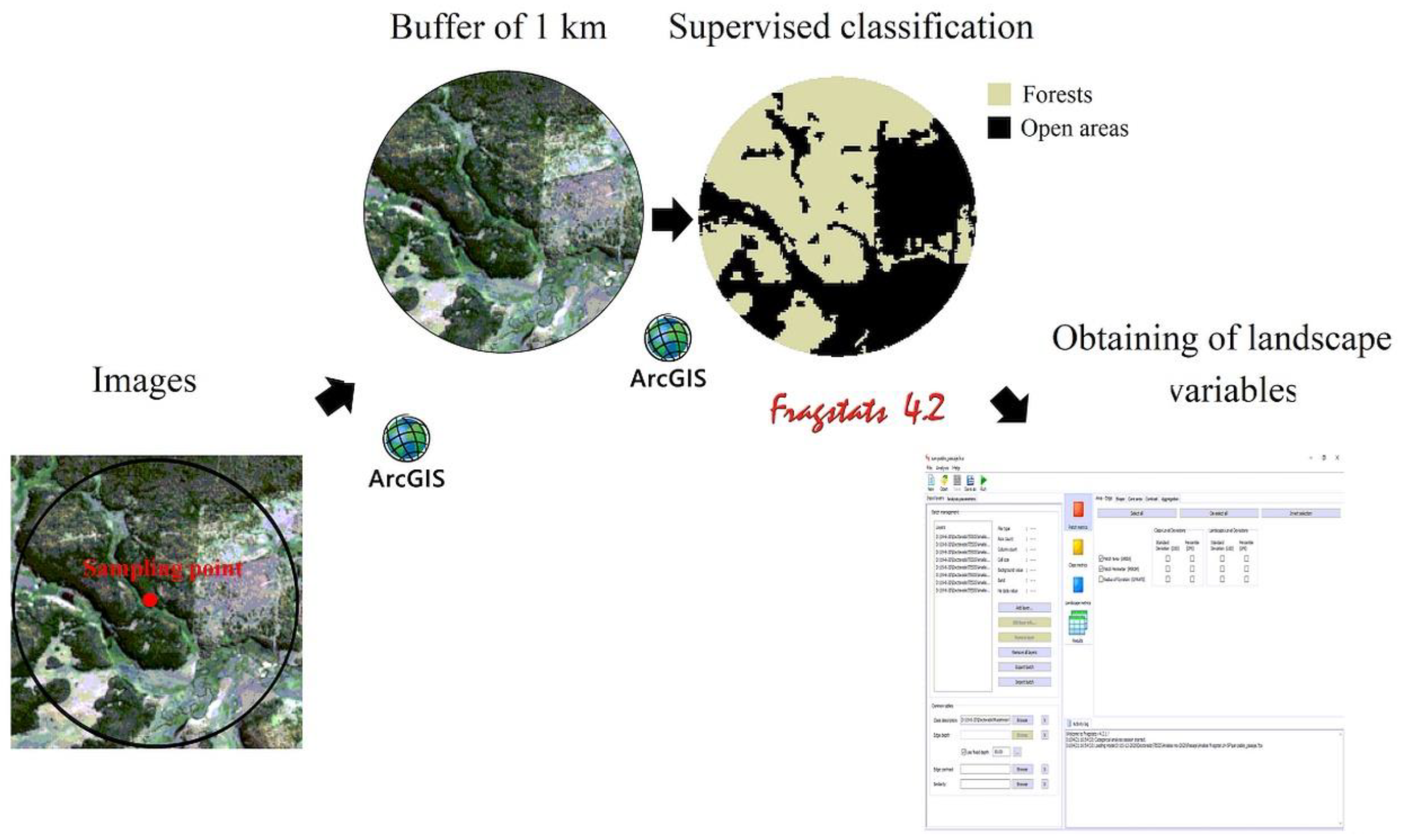
2.4. Landscape Characterization
2.5. Local Stand Characterization
2.6. Data Analysis
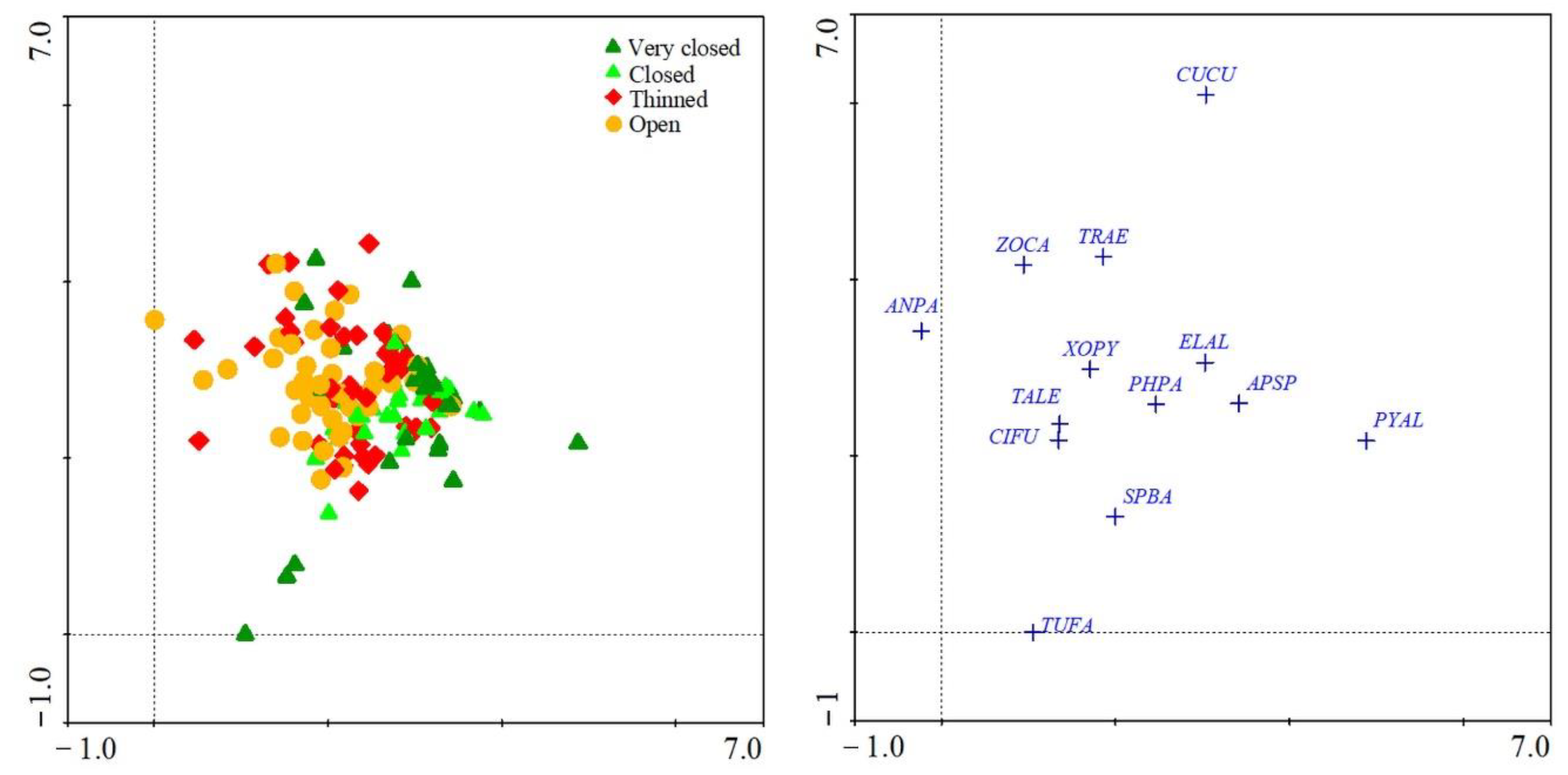
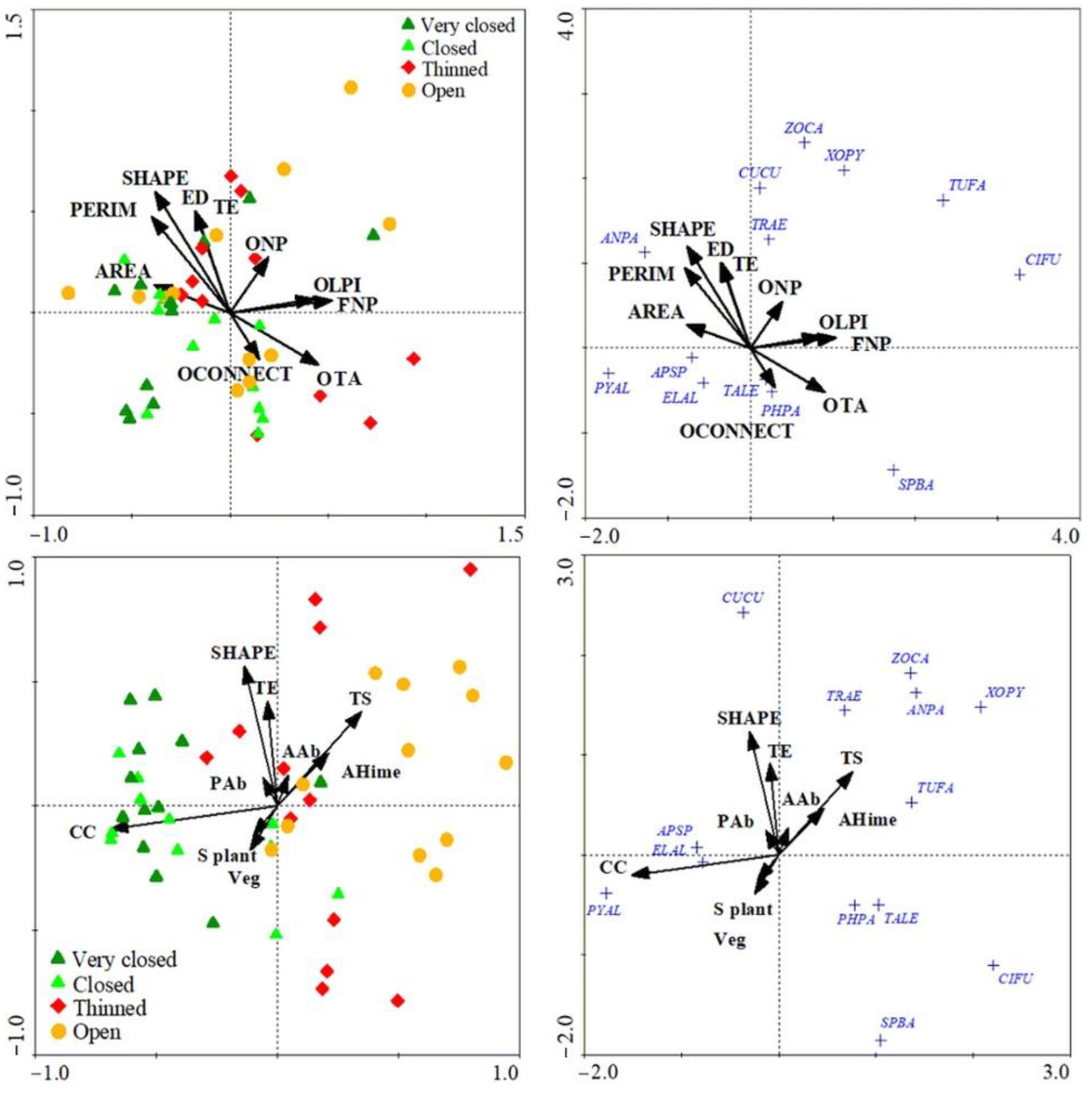
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Characteristics | Habitat Types | ||||
|---|---|---|---|---|---|
| Very Closed (>85% CC) | Closed (65–85% CC) | Thinned (35–65% CC) | Open (<35% CC) | ||
| Habitat structure characteristics | |||||
| Canopy cover | CC (%) | 95.4 ± 7.3 | 82.2 ± 12.0 | 58.5 ± 12.5 | 26.4 ± 9.6 |
| Dominant height | DH (m) | 10.1 ± 2.2 | 11.6 ± 3.0 | 12.4 ± 1.6 | 10.4 ± 136.8 |
| Diameter at breast height | DBH (cm) | 18.6 ± 4.6 | 37.5 ± 10.7 | 41.8 ± 9.6 | 19.0 ± 6.3 |
| Tree density | N (ind ha−1) | 2892.7 ± 1332.2 | 626.3 ± 533.0 | 249.8 ± 136.8 | 416.4 ± 13.1 |
| Basal area | BA (m2 ha−1) | 47.1 ± 16.6 | 34.3 ± 6.6 | 22.3 ± 6.3 | 7.3 ± 9.3 |
| Understory plants cover | U (%) | 69.0 ± 20.9 | 73.4 ± 5.6 | 77.0 ± 19.5 | 87.5 ± 12.3 |
| Debris cover | D (%) | 14.4 ± 9.5 | 14.8 ± 4.0 | 14.9 ± 9.3 | 1.4 ± 7.2 |
| Bryophytes cover | B (%) | 4.0 ± 1.3 | 7.1 ± 4.0 | 3.2 ± 3.8 | 8.4 ± 6.4 |
| Tree saplings cover | TS (%) | 0.3 ± 0.9 | 1.4 ± 2.6 | 5.0 ± 7.2 | 4.0 ± 1.4 |
| Bare soil cover | SO (%) | 12.6 ± 11.9 | 4.8 ± 2.8 | 4.9 ± 3.0 | 2.7 ± 3.8 |
| Food availability characteristics | |||||
| Richness of plants | S plant (n° species) | 6.3 ± 1.7 | 9.1 ± 2.1 | 8.7 ± 1.4 | 6.8 ± 2.9 |
| Plants cover | Veg (%) | 49.6 ± 19.5 | 50.2 ± 7.7 | 49.3 ± 6.4 | 41.0 ± 21.0 |
| Grasses cover | Grasses (%) | 30.7 ± 23.1 | 36.7 ± 11.9 | 36.9 ± 4.7 | 36.9 ± 19.7 |
| Dicots cover | Dicots (%) | 18.9 ± 16.4 | 13.5 ± 9.9 | 12.4 ± 9.0 | 3.6 ± 4.3 |
| Abundance of arthropods captured by attraction traps | AAb (n° ind) | 3202.1 ± 4826.5 | 1845.4 ± 2457.8 | 367.0 ± 320.0 | 3080.3 ± 3657.8 |
| Proportion of Diptera captured by attraction traps | ADip (%) | 93.3 ± 4.4 | 90.0 ± 8.8 | 70.5 ± 17.7 | 87.9 ± 9.9 |
| Proportion of Hymenoptera captured by attraction traps | AHyme (%) | 2.2 ± 2.2 | 5.0 ± 7.1 | 20.5 ± 19.6 | 4.4 ± 6.3 |
| Proportion of Lepidoptera captured by attraction traps | ALep (%) | 3.1 ± 3.3 | 4.0 ± 3.0 | 7.4 ± 9.2 | 7.2 ± 9.2 |
| Proportion of Coleoptera captured by attraction traps | ACole (%) | 1.3 ± 1.6 | 0.9 ± 1.4 | 1.2 ± 1.6 | 0.3 ± 0.3 |
| Abundance of arthropods captured by pitfalls | PAb (n° ind) | 1100.3 ± 1262.0 | 303.8 ± 258.8 | 299.5 ± 223.8 | 536.6 ± 640.5 |
| Proportion of Diptera captured by pitfalls | PDip (%) | 74.2 ± 16.0 | 56.0 ± 14.4 | 45.1 ± 23.2 | 63.2 ± 20.5 |
| Proportion of Hymenoptera captured by pitfalls | PHyme (%) | 6.7 ± 4.2 | 4.1 ± 5.7 | 2.6 ± 3.0 | 4.1 ± 3.2 |
| Proportion of Acarina captured by pitfalls | PAca (%) | 5.1 ± 6.2 | 4.9 ± 4.3 | 10.2 ± 9.1 | 6.9 ± 7.5 |
| Proportion of Coleoptera captured by pitfalls | PCole (%) | 5.0 ± 3.8 | 16.6 ± 9.1 | 14.2 ± 10.7 | 12.2 ± 8.3 |
| Proportion of larvae captured by pitfalls | PLarv (%) | 3.8 ± 3.8 | 4.9 ± 3.6 | 9.0 ± 10.2 | 3.4 ± 3.1 |
References
- Brandolin, P.G.; Blendinger, P.G. Effect of habitat and landscape structure on waterbird abundance in wetlands of central Argentina. Wetl. Ecol. Manag. 2015, 24, 93105. [Google Scholar] [CrossRef]
- Carrara, E.; Arroyo-Rodríguez, V.; Vega-Rivera, J.H.; Schondube, J.E.; de Freitas, S.M.; Fahrig, L. Impact of landscape composition and configuration on forest specialist and generalist bird species in the fragmented Lacandona rainforest, Mexico. Biol. Conserv. 2015, 184, 117–126. [Google Scholar] [CrossRef]
- Dixon, A.P.; Baker, M.E.; Ellis, E.C. Agricultural landscape composition linked with acoustic measures of avian diversity. Land 2020, 9, 145. [Google Scholar] [CrossRef]
- Dunning, J.B.; Danielson, B.J.; Pulliam, H.R. Ecological processes that affect populations in complex landscapes. Oikos 1992, 65, 169–175. [Google Scholar] [CrossRef]
- McGarigal, K.; Marks, B.J. Fragstats: Spatial Pattern Analysis Program for Quantifying Landscape Structure; US Forest Service Pacific Northwest Research Station: Portland, OR, USA, 1995; 122p. [Google Scholar]
- Bonfim, F.C.G.; Dodonov, P.; Cazetta, E. Landscape composition is the major driver of the taxonomic and functional diversity of tropical frugivorous birds. Landsc. Ecol. 2021, 36, 2535–2547. [Google Scholar] [CrossRef]
- Haddad, N.M.; Gonzalez, A.; Brudvig, L.A.; Burt, M.A.; Levey, D.J.; Damschen, E.I. Experimental evidence does not support the habitat amount hypothesis. Ecography 2017, 40, 48–55. [Google Scholar] [CrossRef]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. 2006, 81, 117–142. [Google Scholar] [CrossRef]
- Calamari, N.C.; Zaccagnini, M.E. Respuesta de las aves a la fragmentación del monte nativo entrerriano: Implicancias para la conservación y la agricultura sustentable (Response of birds to the fragmentation of the native forest in Entre Ríos: Implications for conservation and sustainable agriculture). In Agricultura Sustentable en Entre Ríos (Sustainable Agriculture in Entre Ríos); Caviglia, O.P., Paparotti, O.F., Sasal, M.C., Eds.; Ediciones INTA: Buenos Aires, Argentina, 2007; pp. 119–125. [Google Scholar]
- Vala, M.A.; Mitchell, G.W.; Hannah, K.C.; Put, J.; Wilson, S. The effects of landscape composition and configuration on Eastern Whip-poor-will (Caprimulgus vociferous) and Common Nighthawk (Chordeiles minor) occupancy in an agroecosystem. Avian Conserv. Ecol. 2020, 15, 24. [Google Scholar] [CrossRef]
- Baltag, E.S.; Pocora, V.; Bolboaca, L.E.; Ion, C. The Importance of Low-Intensive Agricultural Landscape for Birds of Prey. Land 2021, 10, 252. [Google Scholar] [CrossRef]
- Polis, G.A.; Power, M.E.; Huxel, G.R. Food Webs at the Landscape Level; University of Chicago Press: Chicago, IL, USA, 2004. [Google Scholar]
- Uezu, A.; Metzger, J.P. Vanishing bird species in the Atlantic Forest: Relative importance of landscape configuration, forest structure and species characteristics. Biodivers. Conserv. 2011, 20, 3627–3643. [Google Scholar] [CrossRef]
- Klingbeil, B.T.; Willig, M.R. Matrix composition and landscape heterogeneity structure multiple dimensions of biodiversity in temperate forest birds. Biodivers. Conserv. 2016, 25, 2687–2708. [Google Scholar] [CrossRef]
- Calamari, N.C.; Vilella, F.J.; Sica, Y.V.; Mercuri, P.A. Patch and landscape responses of bird abundance to fragmentation in agroecosystems of east-central Argentina. Avian Conserv. Ecol. 2018, 13, 3. [Google Scholar] [CrossRef]
- Bhakti, T.; Goulart, F.; de Azevedo, C.S.; Antonini, Y. Does scale matter? The influence of three-level spatial scales on forest bird occurrence in a tropical landscape. PLoS ONE 2018, 13, e0198732. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.M.; Campbell Grant, E.H.; Neel, M.C.; Fagan, W.F.; Marra, P.P. Landscape matrix mediates occupancy dynamics of Neotropical avian insectivores. Ecol. Appl. 2011, 21, 1837–1850. [Google Scholar] [CrossRef]
- Cornell, H.V.; Karlson, R.H. Local and regional processes as controls of species richness. In Spatial Ecology: The Role of Space in Population Dynamics and Interspecific Interactions; Tilman, D., Kareiva, P., Eds.; Princeton University Press: Princeton, NY, USA, 1997; pp. 250–268. [Google Scholar]
- Estades, C.F.; Temple, S.A. Deciduous-forest bird communities in a fragmented landscape dominated by exotic pine plantations. Ecol. Appl. 1999, 9, 573–585. [Google Scholar] [CrossRef]
- Mörtberg, U.M. Resident bird species in urban forest remnants, landscape and habitat perspectives. Landsc. Ecol. 2001, 16, 193–203. [Google Scholar] [CrossRef]
- Peri, P.L.; Lencinas, M.V.; Bousson, J.; Lasagne, R.; Soler, R.; Bahamonde, H.; Martínez Pastur, G. Biodiversity and ecological long-term plots in Southern Patagonia to support sustainable land management: The case of PEBANPA network. J. Nat. Conserv. 2016, 34, 51–64. [Google Scholar] [CrossRef]
- Collado, L. Los bosques de Tierra del Fuego. Análisis de su estratificación mediante imágenes satelitales para el inventario forestal de la provincia (The forests of Tierra del Fuego. Analysis of its stratification through satellite images for the forest inventory of the province). Multequina 2001, 10, 15. [Google Scholar]
- Gea-Izquierdo, G.; Martínez Pastur, G.; Cellini, J.M.; Lencinas, M.V. Forty years of silvicultural management in southern Nothofagus pumilio primary forests. For. Ecol. Manag. 2004, 201, 335–347. [Google Scholar] [CrossRef]
- Peri, P.L.; Hansen, N.E.; Bahamonde, H.A.; Lencinas, M.V.; von Müller, A.R.; Ormaechea, S.; Gargaglione, V.; Soler, R.M.; Tejera, L.; Lloyd, C.E.; et al. Silvopastoral systems under native forest in Patagonia, Argentina. In Silvopastoral Systems in Southern South America; Peri, P.L., Dube, F., Varella, A., Eds.; Series: Advances in Agroforestry; Springer: Bern, Switzerland, 2016; Volume 11, pp. 117–168. [Google Scholar] [CrossRef]
- Peri, P.L.; Caballé, G.; Hansen, N.E.; Bahamonde, H.A.; Lencinas, M.V.; Von Müller, A.R.; Ormaechea, S.; Gargaglione, V.; Soler, R.; Sarasola, M.; et al. Silvopastoral systems in Patagonia, Argentina. In Temperate Agroforestry Systems; Gordon, A.M., Newman, S.M., Coleman, B.R.W., Eds.; CABI: Boston, MA, USA, 2017; pp. 252–273. [Google Scholar]
- Perera, A.; Peterson, U.; Martínez Pastur, G.; Iverson, L. Ecosystem Services from Forest Landscapes: Broadscale Considerations; Springer: Cham, Switzerland, 2018; p. 265. [Google Scholar]
- Martínez Pastur, G.; Schlichter, T.; Matteucci, S.D.; Gowda, J.H.; Huertas Herrera, A.; Toro Manríquez, M.; Lencinas, M.V.; Cellini, J.M.; Peri, P.L. Synergies and trade-offs of national conservation policy and agro-forestry management over forest loss in Argentina during the last decade. In Latin America in Times of Global Environmental Change; Lorenzo, C., Ed.; Springer: Cham, Switzerland, 2020; pp. 135–155. [Google Scholar]
- Lencinas, M.V.; Martínez Pastur, G.; Medina, M.; Busso, C. Richness and density of birds in timber Nothofagus pumilio forests and their unproductive associated environments. Biodivers. Conserv. 2005, 14, 2299–2320. [Google Scholar] [CrossRef]
- Deferrari, G.; Camilión, C.; Martínez Pastur, G.; Peri, P.L. Changes in Nothofagus pumilio forest biodiversity during the forest management cycle. 2. Birds. Biodivers. Conserv. 2001, 10, 2093–2108. [Google Scholar] [CrossRef]
- Lencinas, M.V.; Martínez Pastur, G.; Gallo, E.; Cellini, J.M. Alternative silvicultural practices with variable retention improve bird conservation in managed South Patagonian forests. For. Ecol. Manag. 2009, 258, 472–480. [Google Scholar] [CrossRef]
- Lencinas, M.V.; Cellini, J.M.; Benitez, J.; Peri, P.L.; Martínez Pastur, G. Variable retention forestry conserves habitat of bird species in Patagonian Nothofagus pumilio forests. Ann. For. Res. 2018, 61, 147–160. [Google Scholar] [CrossRef]
- Benitez, J.; Barrera, M.D.; Sola, F.J.; Blazina, A.P.; Martínez Pastur, G.J.; Peri, P.L.; Lencinas, M.V. Effects of long-term low intensity silviculture and habitat on birds in Nothofagus antarctica forests of south Patagonia. For. Ecol. Manag. 2022, 516, 120254. [Google Scholar] [CrossRef]
- Ivancich, H.S. Relaciones Entre la Estructura Forestal y el Crecimiento del Bosque de Nothofagus antarctica en Gradientes de edad y Calidad de Sitio. Ph.D. Thesis, Universidad Nacional de La Plata, La Plata, Argentina, 2013. [Google Scholar] [CrossRef]
- Peri, P.L. Sistemas silvopastoriles en bosques de Nothofagus antarctica. Revisión del conocimiento actual en Patagonia Sur, Argentina. (Silvopastoral systems in Nothofagus antarctica forests. Review of current knowledge in South Patagonia, Argentina). Cienc. Investig. For. 2011, 17, 217–242. [Google Scholar] [CrossRef]
- Moore, D. Flora of Tierra del Fuego; Missouri Botanical Garden: London, UK, 1983; 395p. [Google Scholar]
- Martínez Pastur, G.J.; Cellini, J.M.; Lencinas, M.V.; Barrera, M.; Peri, P.L. Environmental variables influencing regeneration of Nothofagus pumilio in a system with combined aggregated and dispersed retention. For. Ecol. Manag. 2011, 261, 178–186. [Google Scholar] [CrossRef]
- Soler, R.; Martínez Pastur, G.; Lencinas, M.V.; Borrelli, L. Differential forage use between large native and domestic herbivores in Southern Patagonian Nothofagus forests. Agrofor. Syst. 2012, 85, 397–409. [Google Scholar] [CrossRef]
- Peri, P.L.; Martínez Pastur, G.; Monelos, L.; Livraghi, E.; Allogia, M.; Christiansen, R.; Sturzenbaum, M.V. Sistemas silvopastoriles en bosques nativos de ñire: Una estrategia para el desarrollo sustentable en la Patagonia Sur (Silvopastoral systems in native ñire forests: A strategy for sustainable development in South Patagonia). In Dinámicas Mundiales, Integración Regional y Patrimonio en Espacios Periféricos (Global Dynamics, Regional Integration and Heritage in Peripheral Spaces), 1st ed.; Zárate, R., Artesi, L., Eds.; Universidad Nacional de la Patagonia Austral: Río Gallegos, Argentina, 2005; pp. 251–259. [Google Scholar]
- A Classification of the Bird Species of South America. Available online: www.museum.lsu.edu/~Remsen/SACCBaseline.htm (accessed on 8 April 2020).
- Humphrey, P.S.; Peterson, D.; Reynolds, R.T.; Humphrey, P.W. Preliminary Smithsonian Manual: Birds of Isla Grande (Tierra del Fuego); Smithsonian Institute Press: Washington, DC, USA; University of Kansas Museum of Natural History: Lawrence, KS, USA, 1970; 441p. [Google Scholar]
- Pizarro, J.C.; Anderson, C.B.; Rozzi, R. Birds as marine-terrestrial linkages in sub-polar archipelagic systems: Avian community composition, function and seasonal dynamics in the Cape Horn Biosphere Reserve (54-55 S), Chile. Polar Biol. 2012, 35, 39–51. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps Computer Software Program Produced by the Authors at the University of Massachusetts, Amherst. Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 20 July 2020).
- Bitterlich, W. Die Winkelzählprobe. Allgemeine Forst und Holzwirtschaftliche Zeitung 1947, 58, 94–96. [Google Scholar]
- Lemmon, P.E. A new instrument for measuring forest overstory density. J. For. 1957, 55, 667668. [Google Scholar]
- Braun-Blanquet, J. Fitosociología: Bases Para el Estudio de las Comunidades Vegetales; Blume: Madrid, Spain, 1979; 820p. [Google Scholar]
- Lencinas, M.V.; Martínez Pastur, G.; Gallo, E.; Cellini, J.M. Alternative silvicultural practices with variable retention to improve understory plant diversity conservation in southern Patagonian forests. For. Ecol. Manag. 2011, 262, 1236–1250. [Google Scholar] [CrossRef]
- Lencinas, M.V.; Martínez Pastur, G.; Gallo, E.; Cellini, J.M. Decreasing negative impacts of harvesting over insect diversity using variable retention silviculture in southern Patagonian forests. J. Insect Conserv. 2014, 18, 479–495. [Google Scholar] [CrossRef]
- Correa, M.N. Flora Patagónica (Patagonian Flora); Colección Científica Instituto Nacional de Tecnología Agropecuaria: Buenos Aires, Argentina, 1969–1998. [Google Scholar]
- Zuloaga, F.O.; Belgrano, M.J.; Zanotti, C.A. Actualización del catálogo de las plantas vasculares del Cono Sur (Updating of the catalog of vascular plants of the Southern Cone). Darwiniana 2019, 7, 208–278. [Google Scholar] [CrossRef]
- Archuby, D.; Marti, L.; Montalti, D.; Soave, G.; Camperi, A.; Arambarri, A.; Darrieu, C.A. Alimentación del cabecita negra austral (Carduelis barbata) durante el otoño (Feeding of the Austral blackhead (Carduelis barbata) during the autumn). Hornero 2007, 22, 65–68. [Google Scholar]
- Brown, C.E.; Anderson, C.B.; Ippi, S.; Sherriffs, M.F.; McGehee, S.; Rozzi, R. Autoecología del Fío-Fío (Elaenia albiceps Lafresnaye &D’Orbigny) en los Bosques Subantárticos de la Reserva de Biosfera Cabo de Hornos, Chile (Autoecology of the Fío-Fío (Elaenia albiceps Lafresnaye & D’Orbigny) in the Subantarctic Forests of the Cape Horn Biosphere Reserve, Chile). An. Inst. Patagon. 2007, 35, 2940. [Google Scholar]
- McGehee, S.M.; Eitniear, J.C. Diet of the patagonian Sierra-finch (Phrygilus patagonicus) on Navarino island, Chile. Ornitol. Neotrop. 2007, 18, 449–452. [Google Scholar]
- Díaz, S. Biología y conservación de la cachaña (Enicognathus ferrugineus) en Argentina (Biology and conservation of cachaña (Enicognathus ferrugineus) in Argentina). Hornero 2012, 27, 17–25. [Google Scholar]
- Muirhead-Thomson, R.S. Trap Responses of Flying Insects; Academic Press: San Diego, CA, USA, 1991. [Google Scholar]
- Cheli, G.H.; Corley, J.C. Efficient sampling of ground-dwelling arthropods using pitfall traps in arid steppes. Neotrop. Entomol. 2010, 39, 912–917. [Google Scholar] [CrossRef]
- Hill, M.O. DECORANA A FORTRAN Program for Detrended Correspondence Analysis and Reciprocal Averaging; Section of Ecology and Systematics, Cornell University: Ithaca, NY, USA, 1979. [Google Scholar]
- Ter Braak, C.J.F. Canonical correspondence analysis: A new eigenvector method for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Ithaca (MicrocomputerPower): New York, NY, USA, 2002; 500p. [Google Scholar]
- Birdlife International. Available online: Birdlife.org (accessed on 14 June 2022).
- Gönc, R.L. 2013. Estudio de la Estructura de las Comunidades de Artrópodos Asociados a Ñirantales (Nothofagus antarctica) del centro-oeste de la Provincia del Chubut, Patagonia Argentina, Sometidos a Diferentes Tipos de Impacto Ambiental (Study of the Structure of the Communities of Arthropods Associated with Ñirantales (Nothofagus antarctica) in the Center-West of the Province of Chubut, Patagonia Argentina, Under Different Types of Environmental Impact). Ph.D. Thesis, Universidad Nacional de la Plata, La Plata, Argentina, 2013. [Google Scholar]
- Rosas, Y.M. Correlación Espacial Entre las Diferentes Estrategias de uso en Patagonia Sur, la Provisión de los Servicios Ecosistémicos y la Conservación de la Biodiversidad (Spatial Correlation between the Different Use Strategies in South Patagonia, the Provision of Ecosystem Services and the Conservation of Biodiversity). Ph.D. Thesis, Universidad Nacional de la Plata, La Plata, Argentina, 2020. [Google Scholar]
- Veblen, T.T.; Donoso, C.; Kitzberger, T.; Rebertus, A.J. Ecology of southern Chilean and Argentinean Nothofagus forests. In Ecology and Biogeography of Nothofagus Forests; Veblen, T.T., Hill, R.S., Read, J., Eds.; Yale University Press: London, UK, 1996; pp. 293–353. [Google Scholar]
- Veblen, T.T.; Kitzberger, T.; Raffaele, E.; Lorenz, D.C. Fire history and vegetation changes in northern Patagonia, Argentina. In Fire and Climatic Change in Temperate Ecosystems of the Western Americas; Veblen, T.T., Baker, W.L., Montenegro, G., Swetnam, T.W., Eds.; Springer: New York, NY, USA, 2003; pp. 265–295. [Google Scholar]
- Vettese, E.S.; Villalba, R.; Orellana Ibáñez, I.A.; Peri, P.L. Tree-growth variations of Nothofagus antarctica related to climate and land use changes in Southern Patagonia, Argentina. In Latin American Dendroecology; Pompa-García, M., Camarero, J.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 331–354. [Google Scholar]
- Bravo, S.P.; Cueto, V.R.; Gorosito, C.A. Migratory timing, rate, routes and wintering areas of White-crested Elaenia (Elaenia albiceps chilensis), a key seed disperser for Patagonian forest regeneration. PLoS ONE 2017, 12, e0170188. [Google Scholar] [CrossRef]
- Csikós, N.; Szilassi, P. Modelling the Impacts of Habitat Changes on the Population Density of Eurasian Skylark (Alauda arvensis) Based on Its Landscape Preferences. Land 2021, 10, 306. [Google Scholar] [CrossRef]
- Fahrig, L. Rethinking patch size and isolation effects: The habitat amount hypothesis. J. Biogeogr. 2013, 40, 1649–1663. [Google Scholar] [CrossRef]
- Capllonch, P. Migraciones de aves en el litoral Argentino (Bird migrations in the Argentine coast). Miscelánea 2004, 12, 363–368. [Google Scholar]
- Bellocq, M.I.; Filloy, J.; Zurita, G.A.; Apellaniz, M.F. Responses in the abundance of generalist birds to environmental gradients: The rufous-collared sparrow (Zonotrichia capensis) in the southern Neotropics. Ecoscience 2011, 18, 354–362. [Google Scholar] [CrossRef]
- Smith, A.C.; Fahrig, L.; Francis, C.M. Landscape size affects the relative importance of habitat amount, habitat fragmentation, and matrix quality on forest birds. Ecography 2011, 34, 103–113. [Google Scholar] [CrossRef]
- Toms, J.D.; Lesperance, M.L. Piecewise regression: A tool for identifying ecological thresholds. Ecology 2003, 84, 2034–2041. [Google Scholar] [CrossRef]
- Muhamad, D.; Okubo, S.; Miyashita, T.; Takeuchi, K. Effects of habitat type, vegetation structure, and proximity to forests on bird species richness in a forest–agricultural landscape of West Java, Indonesia. Agrofor. Syst. 2013, 87, 1247–1260. [Google Scholar] [CrossRef]
- Menon, T.; Sridhar, H.; Shahabuddin, G. Effects of extractive use on forest birds in Western Himalayas: Role of local and landscape factors. For. Ecol. Manag. 2019, 448, 457–465. [Google Scholar] [CrossRef]
- Vergara, P.M.; Schlatter, R.P. Aggregate retention in two Tierra del Fuego Nothofagus forests: Short-term effects on bird abundance. For. Ecol. Manag. 2006, 225, 213–224. [Google Scholar] [CrossRef]
- Stratford, J.A.; Stouffer, P.C. Forest fragmentation alters microhabitat availability for Neotropical terrestrial insectivorous birds. Biol. Conserv. 2015, 188, 109–115. [Google Scholar] [CrossRef]
- Cahill, J.R.A.; Merckx, T.; Van Dyck, H.; Fernandez, M.; Matthysen, E. Lower density of arthropod biomass in small high-Andes Polylepis fragments affects habitat use in insectivorous birds. Ecosphere 2021, 12, e03401. [Google Scholar] [CrossRef]
- Barzan, F.R.; Bellis, L.M.; Dardanelli, S. Livestock grazing constrains bird abundance and species richness: A global meta-analysis. Basic Appl. Ecol. 2021, 56, 289–298. [Google Scholar] [CrossRef]
- Picchio, R.; Mederski, P.S.; Tavankar, F. How and how much, do harvesting activities affect forest soil, regeneration and stands? Curr. For. Rep. 2020, 6, 115–128. [Google Scholar] [CrossRef]




| Type | Variable | Acronym | Description | |
|---|---|---|---|---|
| Forest patch | Area | AREA | Total area (ha). | |
| Perimeter | PERIM | Length of perimeter (m). | ||
| Shape | SHAPE | Perimeter of patch divided by the square root of patch area, adjusted by a constant to fit a standard square. | ||
| Class | Forest | Total area | FTA | Sum of areas of all forest patches, divided by 10,000 (ha). |
| Number of patches | FNP | Number of forest patches. | ||
| Largest patch index | FLPI | Area of largest patch of forest divided by total landscape area, multiplied by 100 (%). | ||
| Connectivity | FCONNECT | Proportion of functional joining among all forest patches (%). | ||
| Open area | Total area | OTA | Sum of areas of all open area patches, divided by 10,000 (ha). | |
| Number of patches | ONP | Number of open area patches. | ||
| Largest patch index | OLPI | Area of largest patch of forest divided by total landscape area, multiplied by 100 (%). | ||
| Connectivity | OCONNECT | Proportion of functional joining among all forest patches (%). | ||
| Landscape | Total edge | TE | Sum of the lengths of all edge segments in the landscape (m). | |
| Edge density | DE | Sum of the lengths of all edge segments in the landscape, divided by the total landscape area and multiplied by 10,000 (m/ha). | ||
| Species | Acronym | Trophic Level | Migratory Status | Mean Density by Habitat Types | ||||
|---|---|---|---|---|---|---|---|---|
| Common Name | Very Closed (>85% CC) | Closed (65–85% CC) | Thinned (35–65% CC) | Open (<35% CC) | ||||
| Phrygilus patagonicus | Patagonia Sierra Finch | PHPA | Omnivore | Resident | 1.7 | 1.3 | 1.3 | 3.1 |
| Zonotrichia capensis | Rufous-collared Sparrow | ZOCA | Omnivore | Migrant | 1.7 | 0.5 | 8.0 | 9.3 |
| Spinus barbatus | Black-chinned Siskin | SPBA | Granivore | Migrant | 4.2 | 9.6 | 20.4 | 6.8 |
| Aphrastura spinicauda * | Thorn-tailed Rayadito | APSP | Insectivore | Resident | 37.8 | 27.7 | 26.0 | 8.5 |
| Cinclodes fuscus | Buff-winged Cinclodes | CIFU | Insectivore | Resident | 0.0 | 0.3 | 1.1 | 1.3 |
| Pygarrhichas albogularis * | White-throated Treerunner | PYAL | Insectivore | Resident | 4.2 | 3.1 | 2.4 | 0.2 |
| Tachycineta leucopyga | Chilean Swallow | TALE | Insectivore | Migrant | 0.0 | 7.8 | 5.8 | 14.1 |
| Curaeus curaeus | Austral Blackbird | CUCU | Omnivore | Resident | 1.3 | 0.0 | 0.9 | 0.5 |
| Troglodytes aedon | House Wren | TRAE | Insectivore | Migrant | 5.1 | 3.6 | 10.6 | 7.8 |
| Turdus falcklandii | Austral Thrush | TUFA | Insectivore | Resident | 2.5 | 0.8 | 1.3 | 2.8 |
| Anairetes parulus | Tufted Tit-Tyrant | ANPA | Insectivore | Resident | 0.4 | 0.0 | 1.3 | 4.5 |
| Elaenia albiceps | White-crested Elaenia | ELAL | Omnivore | Migrant | 31.8 | 17.4 | 14.9 | 11.8 |
| Xolmis pyrope | Fire-eyed Diucon | XOPY | Omnivore | Migrant | 0.0 | 0.0 | 1.5 | 1.8 |
| Type | Variable | Unit | Min | Max | Mean | SD | |
|---|---|---|---|---|---|---|---|
| Forest patch | AREA | ha | 0.9 | 203.9 | 87.1 | 60.6 | |
| PERIM | m | 80.0 | 23,040.0 | 14,262.1 | 7497.4 | ||
| SHAPE | - | 1.0 | 6.9 | 3.9 | 1.4 | ||
| Class | Forest | FTA | ha | 34.8 | 229.3 | 146.8 | 52.0 |
| FNP | - | 5.0 | 76.0 | 26.2 | 17.2 | ||
| FLPI | % | 2.7 | 64.6 | 31.1 | 16.3 | ||
| FCONNECT | % | 3.9 | 40.0 | 12.5 | 8.4 | ||
| Open area | OTA | ha | 86.5 | 281.4 | 169.0 | 52.2 | |
| ONP | - | 5.0 | 57.0 | 17.5 | 13.3 | ||
| OLPI | % | 10.4 | 72.9 | 46.6 | 16.6 | ||
| OCONNECT | % | 4.8 | 40.0 | 17.7 | 9.6 | ||
| Landscape | TE | m | 18,850.0 | 43,860.0 | 29,325.6 | 5430.6 | |
| DE | m ha−1 | 59.6 | 138.6 | 92.9 | 17.2 | ||
| Best Explanatory Variables in CCA | Acronym | Conditional Effects | ||
|---|---|---|---|---|
| LambdaA | p-Value | F-Ratio | ||
| Landscape | SHAPE | 0.14 | 0.002 | 7.88 |
| FNP | 0.07 | 0.006 | 4.09 | |
| OCONNECT | 0.06 | 0.002 | 3.48 | |
| OLPI | 0.04 | 0.004 | 2.56 | |
| PERIM | 0.05 | 0.002 | 2.94 | |
| ONP | 0.05 | 0.006 | 3.19 | |
| AREA | 0.04 | 0.028 | 2.13 | |
| ED | 0.03 | 0.024 | 2.00 | |
| OTA | 0.04 | 0.008 | 2.48 | |
| TE | 0.03 | 0.042 | 2.01 | |
| Landscape and stand | CC | 0.20 | 0.002 | 11.43 |
| SHAPE | 0.14 | 0.002 | 8.69 | |
| PAb | 0.07 | 0.002 | 3.99 | |
| AHyme | 0.06 | 0.002 | 4.24 | |
| TE | 0.07 | 0.002 | 4.12 | |
| S plant | 0.05 | 0.002 | 3.50 | |
| TS | 0.04 | 0.016 | 2.28 | |
| Veg | 0.03 | 0.012 | 2.34 | |
| AAb | 0.03 | 0.038 | 1.88 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benitez, J.; Barrera, M.D.; Rosas, Y.M.; Martínez Pastur, G.J.; Lencinas, M.V. Landscape and Stand Characteristics Influence on the Bird Assemblage in Nothofagus antarctica Forests of Tierra del Fuego. Land 2022, 11, 1332. https://doi.org/10.3390/land11081332
Benitez J, Barrera MD, Rosas YM, Martínez Pastur GJ, Lencinas MV. Landscape and Stand Characteristics Influence on the Bird Assemblage in Nothofagus antarctica Forests of Tierra del Fuego. Land. 2022; 11(8):1332. https://doi.org/10.3390/land11081332
Chicago/Turabian StyleBenitez, Julieta, Marcelo D. Barrera, Yamina M. Rosas, Guillermo J. Martínez Pastur, and María V. Lencinas. 2022. "Landscape and Stand Characteristics Influence on the Bird Assemblage in Nothofagus antarctica Forests of Tierra del Fuego" Land 11, no. 8: 1332. https://doi.org/10.3390/land11081332