
Concept for Genetic Monitoring of Hemiboreal Tree Dynamics in Lithuania



Abstract
:
1. Introduction
2. Genetic Processes of Tree Populations
2.1. Genetic Structure of Tree Species
2.2. Genetic Monitoring System
2.3. Monitoring of Reproductive Behaviour as a Part of Successful Natural Regeneration
2.4. Genetic Effects of Forest Disturbances
3. Lithuania as a Case Study for Europe’s Hemiboreal Forests
4. Hemiboreal Tree Dynamics of the Main Forest Habitat Types
4.1. Tree Regeneration Strategies in Forest Gaps
4.2. Concept of Genetic Monitoring of Hemiboreal Tree Dynamics
5. Ways of Forest Self-Regulation, Natural Regeneration, and Reproduction
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Verzandvoort, S.; Rietra, R.; Alterra, M.H. Pressures on Prime Agricultural Land in Europe. In Proceedings of the Conference “Pressures on Prime Agricultural Land in Europe”, Brussels, Belgium, 19 November 2008; Wageningen UR: Wageningen, The Netherlands, 2009; pp. 1–17. [Google Scholar]
- Key Observed and Projected Climate Change and Impacts for the Main Regions in Europe—European Environment Agency. Available online: https://www.eea.europa.eu/soer/data-and-maps/figures/key-past-and-projected-impacts-and-effects-on-sectors-for-the-main-biogeographic-regions-of-europe-5 (accessed on 24 May 2022).
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate Change Impacts, Adaptive Capacity, and Vulnerability of European Forest Ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.-J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate Change and European Forests: What Do We Know, What Are the Uncertainties, and What Are the Implications for Forest Management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Opgenoorth, L.; Dauphin, B.; Benavides, R.; Heer, K.; Alizoti, P.; Martínez-Sancho, E.; Alía, R.; Ambrosio, O.; Audrey, A.; Auñón, F.; et al. The GenTree Platform: Growth Traits and Tree-Level Environmental Data in 12 European Forest Tree Species. GigaScience 2021, 10, giab010. [Google Scholar] [CrossRef]
- Alberto, F.J.; Aitken, S.N.; Alía, R.; González-Martínez, S.C.; Hänninen, H.; Kremer, A.; Lefèvre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for Evolutionary Responses to Climate Change–Evidence from Tree Populations. Glob. Change Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [PubMed]
- Chuine, I. Why Does Phenology Drive Species Distribution? Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3149–3160. [Google Scholar] [CrossRef] [PubMed]
- Pezzini, F.F.; Ranieri, B.D.; Brandão, D.O.; Fernandes, G.W.; Quesada, M.; Espírito-Santo, M.M.; Jacobi, C.M. Changes in Tree Phenology along Natural Regeneration in a Seasonally Dry Tropical Forest. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2014, 148, 965–974. [Google Scholar] [CrossRef]
- Spielman, D.; Brook, B.W.; Frankham, R. Most Species Are Not Driven to Extinction before Genetic Factors Impact Them. Proc. Natl. Acad. Sci. USA 2004, 101, 15261–15264. [Google Scholar] [CrossRef] [PubMed]
- Kavaliauskas, D.; Fussi, B.; Westergren, M.; Aravanopoulos, F.; Finzgar, D.; Baier, R.; Alizoti, P.; Bozic, G.; Avramidou, E.; Konnert, M.; et al. The Interplay between Forest Management Practices, Genetic Monitoring, and Other Long-Term Monitoring Systems. Forests 2018, 9, 133. [Google Scholar] [CrossRef]
- Fussi, B.; Westergren, M.; Aravanopoulos, F.; Baier, R.; Kavaliauskas, D.; Finzgar, D.; Alizoti, P.; Bozic, G.; Avramidou, E.; Konnert, M.; et al. Forest Genetic Monitoring: An Overview of Concepts and Definitions. Environ. Monit. Assess. 2016, 188, 493. [Google Scholar] [CrossRef]
- ICP Forests. Available online: http://icp-forests.net/ (accessed on 9 May 2022).
- Hoban, S.; Bruford, M.W.; Funk, W.C.; Galbusera, P.; Griffith, M.P.; Grueber, C.E.; Heuertz, M.; Hunter, M.E.; Hvilsom, C.; Stroil, B.K.; et al. Global Commitments to Conserving and Monitoring Genetic Diversity Are Now Necessary and Feasible. BioScience 2021, 71, 964–976. [Google Scholar] [CrossRef] [PubMed]
- Pärli, R.; Lieberherr, E.; Holderegger, R.; Gugerli, F.; Widmer, A.; Fischer, M.C. Developing a Monitoring Program of Genetic Diversity: What Do Stakeholders Say? Conserv. Genet. 2021, 22, 673–684. [Google Scholar] [CrossRef]
- Postolache, D.; Curtu, A.L.; Şofletea, N.; Popescu, F. Conservation and Management of Romanian Forest Genetic Resources in the Context of Climate Change. In Forests of Southeast Europe Under a Changing Climate: Conservation of Genetic Resources; Šijačić-Nikolić, M., Milovanović, J., Nonić, M., Eds.; Advances in Global Change Research; Springer International Publishing: Cham, Switzerland, 2019; pp. 389–399. ISBN 978-3-319-95267-3. [Google Scholar]
- Kraigher, H.; Bajc, M.; Božič, G.; Brus, R.; Jarni, K.; Westergren, M. Forests, Forestry and the Slovenian Forest Genetic Resources Programme. In Forests of Southeast Europe Under a Changing Climate: Conservation of Genetic Resources; Šijačić-Nikolić, M., Milovanović, J., Nonić, M., Eds.; Advances in Global Change Research; Springer International Publishing: Cham, Switzerland, 2019; pp. 29–47. ISBN 978-3-319-95267-3. [Google Scholar]
- Oettel, J.; Lapin, K. Linking Forest Management and Biodiversity Indicators to Strengthen Sustainable Forest Management in Europe. Ecol. Indic. 2021, 122, 107275. [Google Scholar] [CrossRef]
- Lefèvre, F.; Koskela, J.; Hubert, J.; Kraigher, H.; Longauer, R.; Olrik, D.C.; Schüler, S.; Bozzano, M.; Alizoti, P.; Bakys, R.; et al. Dynamic Conservation of Forest Genetic Resources in 33 European Countries. Conserv. Biol. 2013, 27, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Fady, B.; Cottrell, J.; Ackzell, L.; Alía, R.; Muys, B.; Prada, A.; González-Martínez, S.C. Forests and Global Change: What Can Genetics Contribute to the Major Forest Management and Policy Challenges of the Twenty-First Century? Reg. Environ. Change 2016, 16, 927–939. [Google Scholar] [CrossRef]
- Manel, S.; Holderegger, R. Ten Years of Landscape Genetics. Trends Ecol. Evol. 2013, 28, 614–621. [Google Scholar] [CrossRef]
- Angelstam, P. Landscape Analysis as a Tool for the Scientific Management of Biodiversity. Ecol. Bull. 1997, 46, 140–170. [Google Scholar]
- Fomin, V.; Mikhailovich, A.; Zalesov, S.; Popov, A.; Terekhov, G. Development of Ideas within the Framework of the Genetic Approach to the Classification of Forest Types. BALT FOR 2020, 27, 26–39. [Google Scholar] [CrossRef]
- Christensen, N.L.; Peet, R.K. Secondary Forest Succession on the North Carolina Piedmont. In Forest Succession: Concepts and Application; West, D.C., Shugart, H.H., Botkin, D.B., Eds.; Springer Advanced Texts in Life Sciences; Springer: New York, NY, USA, 1981; pp. 230–245. ISBN 978-1-4612-5950-3. [Google Scholar]
- Jõgiste, K.; Frelich, L.E.; Laarmann, D.; Vodde, F.; Baders, E.; Donis, J.; Jansons, A.; Kangur, A.; Korjus, H.; Köster, K.; et al. Imprints of Management History on Hemiboreal Forest Ecosystems in the Baltic States. Ecosphere 2018, 9, e02503. [Google Scholar] [CrossRef]
- Jandl, R.; Spathelf, P.; Bolte, A.; Prescott, C.E. Forest Adaptation to Climate Change—Is Non-Management an Option? Ann. For. Sci. 2019, 76, 48. [Google Scholar] [CrossRef]
- Oliver, C.D. Forest Development in North America Following Major Disturbances. For. Ecol. Manag. 1980, 3, 153–168. [Google Scholar] [CrossRef]
- Berglund, H.; Kuuluvainen, T. Representative Boreal Forest Habitats in Northern Europe, and a Revised Model for Ecosystem Management and Biodiversity Conservation. Ambio 2021, 50, 1003–1017. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.R.; Chen, H.Y.H. Multiple Successional Pathways of Boreal Forest Stands in Central Canada. Ecography 2011, 34, 208–219. [Google Scholar] [CrossRef]
- Borman, M.M.; Pyke, D.A. Successional Theory and the Desired Plant Community Approach. Rangelands 1994, 16, 82–84. [Google Scholar]
- Baliuckas, V. Life History Traits and Broadleaved Tree Genetics. In Acta Universitatis Agriculturae Sueciae; Silvestria, 258; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2002. [Google Scholar]
- Aravanopoulos, F.A. (Phil) Do Silviculture and Forest Management Affect the Genetic Diversity and Structure of Long-Impacted Forest Tree Populations? Forests 2018, 9, 355. [Google Scholar] [CrossRef]
- Stern, K.; Roche, L. Genetics of Forest Ecosystems; Chapman and Hall: London, UK, 1974; ISBN 978-0-387-06095-8. [Google Scholar]
- Roche, L. The Conservation of Forest Gene Resources in Canada. For. Chron. 1971, 47, 215–217. [Google Scholar] [CrossRef]
- Namkoong; Boyle, T.; El-Kassaby, Y.; Palmberg-Lerche, C.; Eriksson, G.; Gregorius, H.-R.; Joly, H.; Kremer, A.; Savolainen, O.; Wickneswari, R.; et al. Criteria and Indicators for Sustainable Forest Management: Assessment and Monitoring of Genetic Variation; Forest Resources Development Service, Forest Resources Division, Forestry Department, Food and Agriculture Organization of the United Nations: Rome, Italy, 2002; p. 29. [Google Scholar]
- Konnert, M.; Maurer, W.; Degen, B.; Kätzel, R. Genetic Monitoring in Forests - Early Warning and Controlling System for Ecosystemic Changes. iForest-Biogeosci. For. 2011, 4, 77. [Google Scholar] [CrossRef]
- Piotti, A.; Leonardi, S.; Heuertz, M.; Buiteveld, J.; Geburek, T.; Gerber, S.; Kramer, K.; Vettori, C.; Vendramin, G.G. Within-Population Genetic Structure in Beech (Fagus Sylvatica L.) Stands Characterized by Different Disturbance Histories: Does Forest Management Simplify Population Substructure? PLoS ONE 2013, 8, e73391. [Google Scholar] [CrossRef]
- Hoban, S.; Arntzen, J.A.; Bruford, M.W.; Godoy, J.A.; Rus Hoelzel, A.; Segelbacher, G.; Vilà, C.; Bertorelle, G. Comparative Evaluation of Potential Indicators and Temporal Sampling Protocols for Monitoring Genetic Erosion. Evol. Appl. 2014, 7, 984–998. [Google Scholar] [CrossRef]
- Charlier, J. Monitoring Gene Level Biodiversity—Aspects and Considerations in the Context of Conservation; Stockholm University: Stockholm, Sweden, 2011. [Google Scholar]
- Ivanova, N.; Fomin, V.; Kusbach, A. Experience of Forest Ecological Classification in Assessment of Vegetation Dynamics. Sustainability 2022, 14, 3384. [Google Scholar] [CrossRef]
- Tripathi, R.S.; Khan, M.L. Regeneration Dynamics of Natural Forests. Proc. Indian Natl. Sci. Academy. Part A Phys. Sci. 2007, 73, 167–196. [Google Scholar]
- Petrokas, R. Forest Climax Phenomenon: An Invariance of Scale. Forests 2020, 11, 56. [Google Scholar] [CrossRef]
- Gorshkov, V.G.; Makarieva, A.M.; Gorshkov, V.V. Revising the Fundamentals of Ecological Knowledge: The Biota–Environment Interaction. Ecol. Complex. 2004, 1, 17–36. [Google Scholar] [CrossRef]
- Hunter, M.L.; Schmiegelow, F.K.A. Wildlife, Forests, and Forestry: Principles of Managing Forests for Biological Diversity, 2nd ed.; Prentice Hall: Boston, MA, USA, 2011; ISBN 978-0-13-501432-5. [Google Scholar]
- McCarthy, J. Gap Dynamics of Forest Trees: A Review with Particular Attention to Boreal Forests. Environ. Rev. 2001, 9, 1–59. [Google Scholar] [CrossRef]
- Harrison, S. 2. Do Taxa Persist as Metapopulations in Evolutionary Time. In Biodiversity Dynamics: Turnover of Populations, Taxa, and Communities; McKinney, M.L., Drake, J.A., Eds.; Columbia University Press: New York, NY, USA, 2001; pp. 19–30. ISBN 978-0-231-50580-2. [Google Scholar]
- Chazdon, R.L. Second Growth: The Promise of Tropical Forest Regeneration in an Age of Deforestation; University of Chicago Press: Chicago, IL, USA, 2014; ISBN 978-0-226-11791-1. [Google Scholar]
- Chazdon, R.L.; Brancalion, P.H.S.; Laestadius, L.; Bennett-Curry, A.; Buckingham, K.; Kumar, C.; Moll-Rocek, J.; Vieira, I.C.G.; Wilson, S.J. When Is a Forest a Forest? Forest Concepts and Definitions in the Era of Forest and Landscape Restoration. Ambio 2016, 45, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Orr, H.A. Fitness and Its Role in Evolutionary Genetics. Nat. Rev. Genet. 2009, 10, 531–539. [Google Scholar] [CrossRef]
- Ehlers, A.; Worm, B.; Reusch, T.B.H. Importance of Genetic Diversity in Eelgrass Zostera Marina for Its Resilience to Global Warming. Mar. Ecol. Prog. Ser. 2008, 355, 1–7. [Google Scholar] [CrossRef]
- Wright, D.; Bishop, J.M.; Matthee, C.A.; von der Heyden, S. Genetic Isolation by Distance Reveals Restricted Dispersal across a Range of Life Histories: Implications for Biodiversity Conservation Planning across Highly Variable Marine Environments. Divers. Distrib. 2015, 21, 698–710. [Google Scholar] [CrossRef]
- Neophytou, C.; Heer, K.; Milesi, P.; Peter, M.; Pyhäjärvi, T.; Westergren, M.; Rellstab, C.; Gugerli, F. Genomics and Adaptation in Forest Ecosystems. Tree Genet. Genomes 2022, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Demetrius, L. Adaptedness and Fitness. Am. Nat. 1977, 111, 1163–1168. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, Migration or Extirpation: Climate Change Outcomes for Tree Populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Skrøppa, T.; Johnsen, Ø. Patterns of Adaptive Genetic Variation in Forest Tree Species; the Reproductive Enviroment as an Evolutionary Force in Picea Abies. In Forest Genetics and Sustainability; Mátyás, C., Ed.; Forestry Sciences; Springer: Dordrecht, Germany, 1999; pp. 49–58. ISBN 978-94-017-1576-8. [Google Scholar]
- Garnier-Géré, P.; Chikhi, L. Population Subdivision, Hardy–Weinberg Equilibrium and the Wahlund Effect. In eLS; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2013; ISBN 978-0-470-01590-2. [Google Scholar]
- Lynch, M.; Ackerman, M.S.; Gout, J.-F.; Long, H.; Sung, W.; Thomas, W.K.; Foster, P.L. Genetic Drift, Selection and the Evolution of the Mutation Rate. Nat. Rev. Genet. 2016, 17, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Schierenbeck, K.A. Population-Level Genetic Variation and Climate Change in a Biodiversity Hotspot. Ann. Bot. 2017, 119, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Leech, S.M.; Almuedo, P.L.; O’Neill, G. Assisted Migration: Adapting Forest Management to a Changing Climate. J. Ecosyst. Manag. 2011, 12. [Google Scholar] [CrossRef]
- Williams, M.I.; Dumroese, R.K. Preparing for Climate Change: Forestry and Assisted Migration. J. For. 2013, 111, 287–297. [Google Scholar] [CrossRef]
- Benito-Garzón, M.; Fernández-Manjarrés, J.F. Testing Scenarios for Assisted Migration of Forest Trees in Europe. New For. 2015, 46, 979–994. [Google Scholar] [CrossRef]
- Aravanopoulos, F.A. Genetic Monitoring in Natural Perennial Plant Populations. Botany 2011, 89, 75–81. [Google Scholar] [CrossRef]
- Hansen, M.M.; Olivieri, I.; Waller, D.M.; Nielsen, E.E.; Allendorf, F.W.; Schwartz, M.K.; Baker, C.S.; Gregovich, D.P.; Jackson, J.A.; Kendall, K.C.; et al. Monitoring Adaptive Genetic Responses to Environmental Change. Mol. Ecology. 2012, 21, 1311–1329. [Google Scholar] [CrossRef] [PubMed]
- Aravanopoulos, F.; Tollefsrud, M.; Graudal, L.; Koskela; Kätzel; Soto; Nagy, L.; Pilipovic, A.; Zhelev, P.; Bozic, G.; et al. Development of Genetic Monitoring Methods for Genetic Conservation Units of Forest Trees in Europe; Bioversity International: Rome, Italy, 2015. [Google Scholar]
- Bajc, M.; Aravanopoulos, F.; Westergren, M.; Fussi, B.; Kavaliauskas, D.; Alizoti, P.; Kiourtsis, F.; Kraigher, H. Manual for Forest Genetic Monitoring; Slovenian Forestry Institute: Ljubljana, Slovenia, 2020. [Google Scholar]
- de Vries, S.M.G.; Alan, M.; Bozzano, M.; Burianek, V.; Collin, E.; Cottrell, J.; Ivankovic, M.; Kelleher, C.T.; Koskela, J.; Rotach, P.; et al. Pan-European Strategy for Genetic Conservation of Forest Trees: And Establishment of a Core Network of Dynamic Conservation Units; European Forest Genetic Resources Programme: Barcelona, Spain, 2015; ISBN 978-92-9255-029-5. [Google Scholar]
- Garzón, M.B.; Robson, T.M.; Hampe, A. ΔTraitSDMs: Species Distribution Models That Account for Local Adaptation and Phenotypic Plasticity. New Phytol. 2019, 222, 1757–1765. [Google Scholar] [CrossRef]
- Frankham, R.; Bradshaw, C.J.A.; Brook, B.W. Genetics in Conservation Management: Revised Recommendations for the 50/500 Rules, Red List Criteria and Population Viability Analyses. Biol. Conserv. 2014, 170, 56–63. [Google Scholar] [CrossRef]
- Hoban, S.; Bruford, M.; D’Urban Jackson, J.; Lopes-Fernandes, M.; Heuertz, M.; Hohenlohe, P.A.; Paz-Vinas, I.; Sjögren-Gulve, P.; Segelbacher, G.; Vernesi, C.; et al. Genetic Diversity Targets and Indicators in the CBD Post-2020 Global Biodiversity Framework Must Be Improved. Biol. Conserv. 2020, 248, 108654. [Google Scholar] [CrossRef]
- Namkoong, G.; Boyle, T.J.B.; Gregorius, H.-R.; Joly, H.; Savolainen, O.; Ratnam, W.; Young, A. Testing Criteria and Indicators for Assessing the Sustainability of Forest Management: Genetic Criteria and Indicators; Centre for International Forestry Research Working Paper; CIFOR: Bogor, Indonesia, 1996. [Google Scholar]
- Aravanopoulos, F.A. (Phil) Conservation and Monitoring of Tree Genetic Resources in Temperate Forests. Curr. Forestry Rep. 2016, 2, 119–129. [Google Scholar] [CrossRef]
- Leroy, G.; Carroll, E.L.; Bruford, M.W.; DeWoody, J.A.; Strand, A.; Waits, L.; Wang, J. Next-Generation Metrics for Monitoring Genetic Erosion within Populations of Conservation Concern. Evol. Appl. 2018, 11, 1066–1083. [Google Scholar] [CrossRef] [PubMed]
- Bradley, N.L.; Leopold, A.C.; Ross, J.; Huffaker, W. Phenological Changes Reflect Climate Change in Wisconsin. Proc. Natl. Acad. Sci. USA 1999, 96, 9701–9704. [Google Scholar] [CrossRef]
- Alizoti, P.G.; Kilimis, K.; Gallios, P. Temporal and Spatial Variation of Flowering among Pinus Nigra Arn. Clones under Changing Climatic Conditions. For. Ecol. Manag. 2010, 259, 786–797. [Google Scholar] [CrossRef]
- Templeton, A.R.; Robertson, R.J.; Brisson, J.; Strasburg, J. Disrupting Evolutionary Processes: The Effect of Habitat Fragmentation on Collared Lizards in the Missouri Ozarks. Proc. Natl. Acad. Sci. USA 2001, 98, 5426–5432. [Google Scholar] [CrossRef]
- Fitter, A.H.; Fitter, R.S.R. Rapid Changes in Flowering Time in British Plants. Science 2002, 296, 1689–1691. [Google Scholar] [CrossRef] [PubMed]
- El-Kassaby, Y.A.; Fashler, A.M.K.; Sziklai, O. Reproductive Phenology and Its Impact on Genetically Improved Seed Production in a Douglas-Fir Seed Orchard. Silvae Genet. 1984, 33, 120–125. [Google Scholar]
- El-Kassaby, Y.A.; Ritland, L.; Fashler, A.M.K.; Devitt, D. The Role of Reproductive Phenology, Parental Balance and Supplemental Mass Pollination in a Sitka Spruce Seed Orchard. Silvae Genet. 1988, 37, 76–82. [Google Scholar]
- Bhumibhamon, S. Studies on Scots Pine Seed Orchards in Finland with Special Emphasis on the Genetic Composition of the Seed. Metsaentutkimuslaitoksen Julk. 1978. [Google Scholar]
- Schauber, E.M.; Kelly, D.; Turchin, P.; Simon, C.; Lee, W.G.; Allen, R.B.; Payton, I.J.; Wilson, P.R.; Cowan, P.E.; Brockie, R.E. Masting by Eighteen New Zealand Plant Species: The Role of Temperature as a Synchronizing Cue. Ecology 2002, 83, 1214–1225. [Google Scholar] [CrossRef]
- Ranta, H.; Hokkanen, T.; Linkosalo, T.; Laukkanen, L.; Bondestam, K.; Oksanen, A. Male Flowering of Birch: Spatial Synchronization, Year-to-Year Variation and Relation of Catkin Numbers and Airborne Pollen Counts. For. Ecol. Manag. 2008, 255, 643–650. [Google Scholar] [CrossRef]
- Newbery, D.M.; Chuyong, G.B.; Zimmermann, L. Mast Fruiting of Large Ectomycorrhizal African Rain Forest Trees: Importance of Dry Season Intensity, and the Resource-Limitation Hypothesis. New Phytol. 2006, 170, 561–579. [Google Scholar] [CrossRef] [PubMed]
- Tuomi, J.; Niemelä, P.; Mannila, R. Resource Allocation on Dwarf Shoots of Birch (Betula Pendula): Reproduction and Leaf Growth. New Phytol. 1982, 91, 483–487. [Google Scholar] [CrossRef]
- Merriam-Webster, I. Merriam-Webster’s Collegiate Dictionary; Merriam-Webster: Springfield, MA, USA, 2014; ISBN 978-0-87779-809-5. [Google Scholar]
- Mund, M.; Kutsch, W.L.; Wirth, C.; Kahl, T.; Knohl, A.; Skomarkova, M.V.; Schulze, E.-D. The Influence of Climate and Fructification on the Inter-Annual Variability of Stem Growth and Net Primary Productivity in an Old-Growth, Mixed Beech Forest. Tree Physiol. 2010, 30, 689–704. [Google Scholar] [CrossRef] [PubMed]
- Selås, V.; Piovesan, G.; Adams, J.M.; Bernabei, M. Climatic Factors Controlling Reproduction and Growth of Norway Spruce in Southern Norway. Can. J. For. Res. 2002, 32, 217–225. [Google Scholar] [CrossRef]
- Müller-Starck, G.; Ziehe, M.; Schubert, R. Genetic Diversity Parameters Associated with Viability Selection, Reproductive Efficiency, and Growth in Forest Tree Species. In Forest Diversity and Function: Temperate and Boreal Systems; Springer: Berlin/Heidelberg, Germany; Scherer-Lorenzen, M., Körner, C., Schulze, E.-D., Eds.; Ecological Studies; Springer: Berlin, Heidelberg, 2005; pp. 87–108. ISBN 978-3-540-26599-3. [Google Scholar]
- Seifert, T.; Müller-Starck, G. Impacts of Fructification on Biomass Production and Correlated Genetic Effects in Norway Spruce (Picea Abies [L.] Karst.). Eur. J. For. Res. 2009, 128, 155–169. [Google Scholar] [CrossRef]
- de Melo, R.B.; Franco, A.C.; Silva, C.O.; Piedade, M.T.F.; Ferreira, C.S. Seed Germination and Seedling Development in Response to Submergence in Tree Species of the Central Amazonian Floodplains. AoB PLANTS 2015, 7, plv041. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, L.; Liu, Z.; Li, Y.; Liu, Q.; Liu, B. Phylogeny, Seed Trait, and Ecological Correlates of Seed Germination at the Community Level in a Degraded Sandy Grassland. Front. Plant Sci. 2016, 7, 1532. [Google Scholar] [CrossRef] [PubMed]
- Dey, D.C.; Knapp, B.O.; Battaglia, M.A.; Deal, R.L.; Hart, J.L.; O’Hara, K.L.; Schweitzer, C.J.; Schuler, T.M. Barriers to Natural Regeneration in Temperate Forests across the USA. New For. 2019, 50, 11–40. [Google Scholar] [CrossRef]
- Koskela, J.; Lefèvre, F.; Schueler, S.; Kraigher, H.; Olrik, D.C.; Hubert, J.; Longauer, R.; Bozzano, M.; Yrjänä, L.; Alizoti, P.; et al. Translating Conservation Genetics into Management: Pan-European Minimum Requirements for Dynamic Conservation Units of Forest Tree Genetic Diversity. Biol. Conserv. 2013, 157, 39–49. [Google Scholar] [CrossRef]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; DellaSala, D.A.; Hutto, R.L.; Lindenmayer, D.B.; Swanson, F.J. The Forgotten Stage of Forest Succession: Early-Successional Ecosystems on Forest Sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar] [CrossRef]
- Shorohova, E.; Kuuluvainen, T.; Kangur, A.; Jõgiste, K. Natural Stand Structures, Disturbance Regimes and Successional Dynamics in the Eurasian Boreal Forests: A Review with Special Reference to Russian Studies. Ann. For. Sci. 2009, 66, 1–20. [Google Scholar] [CrossRef]
- Odum, E.P.; Barrett, G.W. Fundamentals of Ecology, 5th ed.; Thomson Brooks/Cole: Belmont, CA, USA, 2005; ISBN 978-0-534-42066-6. [Google Scholar]
- Angelstam, P.K. Maintaining and Restoring Biodiversity in European Boreal Forests by Developing Natural Disturbance Regimes. J. Veg. Sci. 1998, 9, 593–602. [Google Scholar] [CrossRef]
- Rull, V. Quaternary Palaeoecology and Ecological Theory. Orsis 1990, 5, 91–111. [Google Scholar]
- Russell, G.J. 16. Turnover Dynamics Across Ecological and Geological Scales. In Biodiversity Dynamics; Columbia University Press: New York, NY, USA, 2001; pp. 377–404. ISBN 978-0-231-50580-2. [Google Scholar]
- Larson, J.E.; Funk, J.L. Regeneration: An Overlooked Aspect of Trait-Based Plant Community Assembly Models. J. Ecol. 2016, 104, 1284–1298. [Google Scholar] [CrossRef]
- Ratnam, W.; Rajora, O.P.; Finkeldey, R.; Aravanopoulos, F.; Bouvet, J.-M.; Vaillancourt, R.E.; Kanashiro, M.; Fady, B.; Tomita, M.; Vinson, C. Genetic Effects of Forest Management Practices: Global Synthesis and Perspectives. For. Ecol. Manag. 2014, 333, 52–65. [Google Scholar] [CrossRef]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.-J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive Forest Management in Central Europe: Climate Change Impacts, Strategies and Integrative Concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Wernsdörfer, H.; Caron, H.; Gerber, S.; Cornu, G.; Rossi, V.; Mortier, F.; Gourlet-Fleury, S. Relationships between Demography and Gene Flow and Their Importance for the Conservation of Tree Populations in Tropical Forests under Selective Felling Regimes. Conserv. Genet. 2011, 12, 15–29. [Google Scholar] [CrossRef]
- Huss, J. SILVICULTURE|Natural Stand Regeneration. In Encyclopedia of Forest Sciences; Burley, J., Ed.; Elsevier: Oxford, UK, 2004; pp. 1017–1033. ISBN 978-0-12-145160-8. [Google Scholar]
- Geburek, T.; Müller, F. How Can Silvicultural Management Contribute to Genetic Conservation. In Conservation and Management of Forest Genetic Resources in Europe; Geburek, T., Turok, J., Eds.; Arbora Publishers: Zvolen, Slovakia, 2005; ISBN 8096708813. [Google Scholar]
- Hosius, B.; Leinemann, L.; Konnert, M.; Bergmann, F. Genetic Aspects of Forestry in the Central Europe. Eur. J. For. Res. 2006, 125, 407–417. [Google Scholar] [CrossRef]
- Kerr, G.; Haufe, J. Thinning Practice: A Silvicultural Guide. For. Comm. 2011, 1. [Google Scholar]
- Pope, P.E.; Edlin, H.L.; Chaney, W.R. Forestry—Natural Regeneration|Britannica. Available online: https://www.britannica.com/science/forestry/Natural-regeneration (accessed on 19 April 2022).
- Grebner, D.L.; Bettinger, P.; Siry, J.P.; Boston, K. Chapter 11—Common Forestry Practices. In Introduction to Forestry and Natural Resources, 2nd ed.; Grebner, D.L., Bettinger, P., Siry, J.P., Boston, K., Eds.; Academic Press: San Diego, CA, USA, 2022; pp. 265–294. ISBN 978-0-12-819002-9. [Google Scholar]
- Brazaitis, G.; Marozas, V.; Augutis, D.; Preikša, Ž.; Šaudytė-Manton, S. Lithuanian Forest Habitat Management Recommendations—“Guidelines for the Management of Natural Forest Habitat Types of EC Importance”; Naturalit: Vilnius, Lietuva, 2021. [Google Scholar]
- Vaičys, M.; Mažvila, J. The Influence of Soil Characteristics on Plant Productivity and Ecological Stability. Ekologija 2009, 55, 99–106. [Google Scholar] [CrossRef]
- Buivydaitė, V. Classification of Soils of Lithuania Based on FAO-Unesco Soil Classification System and WRB. In Proceedings of the 17 World Congress of Soil Science, Bangkok, Thailand, 14–20 August 2002; pp. 2189-1–2189-13. [Google Scholar]
- Karazija, S. Forest types of Lithuania; Mokslas: Vilnius, Lithuania, 1988; ISBN 978-5-420-00421-0. [Google Scholar]
- HoustonDurrant, T.; de Rigo, D.; Caudullo, G. Fagus Sylvatica in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., HoustonDurrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; p. e012b90. ISBN 978-92-79-36740-3. [Google Scholar]
- Godvod, K.; Brazaitis, G.; Bačkaitis, J.; Kulbokas, G. The Development and Growth of Larch Stands in Lithuania. J. For. Sci. 2018, 64, 199–206. [Google Scholar] [CrossRef]
- Petrokas, R.; Baliuckas, V.; Manton, M. Successional Categorization of European Hemi-Boreal Forest Tree Species. Plants 2020, 9, 1381. [Google Scholar] [CrossRef] [PubMed]
- Bohn, U.; Gollub, G.; Hettwer, C.; Weber, H.; Neuhäuslová, Z.; Raus, T.; Schlüter, H. Karte Der Natürlichen Vegetation Europas/Map of the Natural Vegetation of Europe-Maßstab/Scale 1:2,500,000; Federal Agency for Nature Conservation: Bonn, Germany, 2000. [Google Scholar]
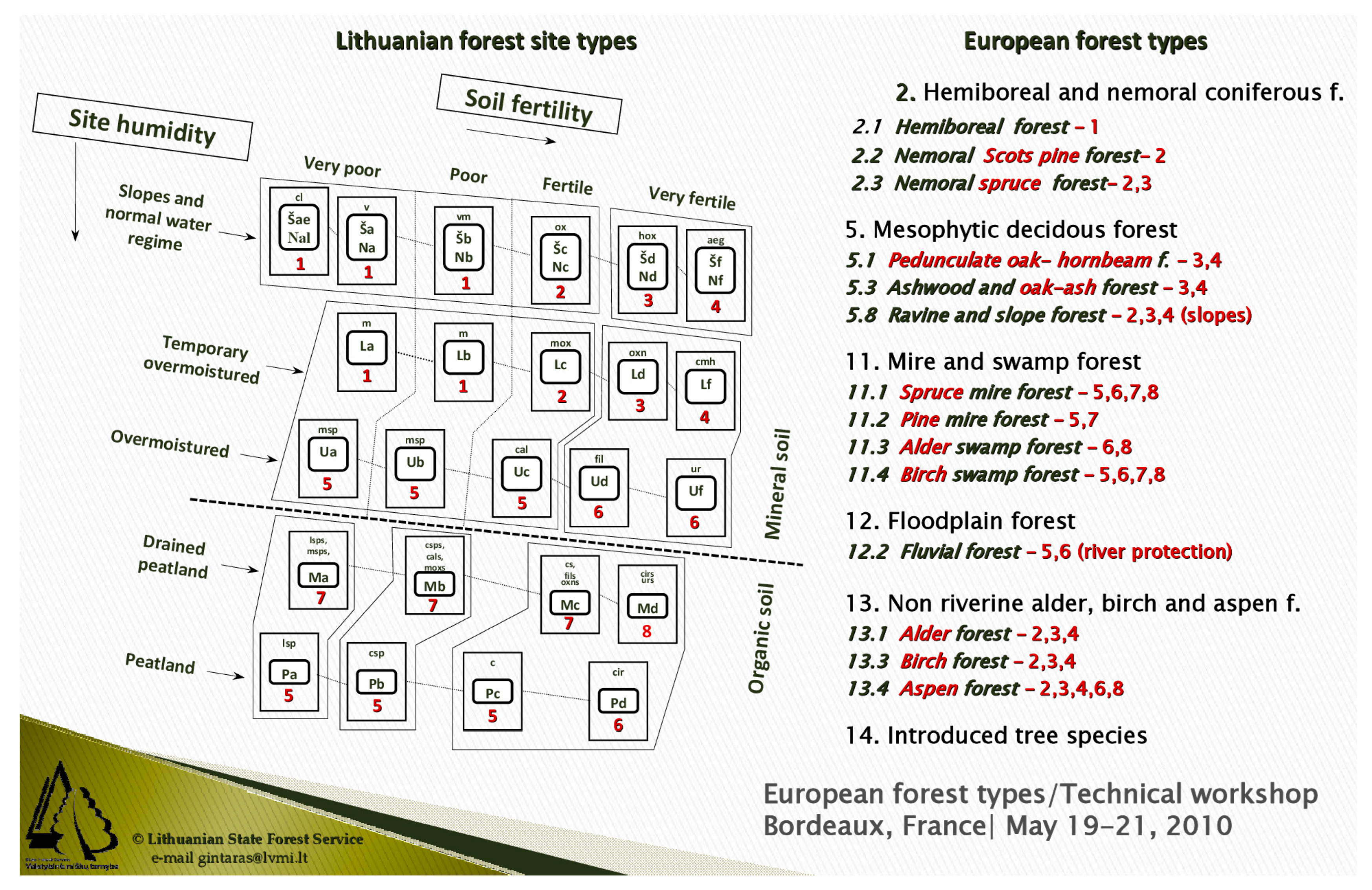
- Kulbokas, G. Experiences from Lithuania-UNECE Technical Workshop on European Forest Types; Unpublished Presentation; Lithuanian State Forest Service: Bordeaux, France, 2010. [Google Scholar]
- Giannetti, F.; Barbati, A.; Mancini, L.D.; Travaglini, D.; Bastrup-Birk, A.; Canullo, R.; Nocentini, S.; Chirici, G. European Forest Types: Toward an Automated Classification. Ann. For. Sci. 2018, 75, 1–14. [Google Scholar] [CrossRef]
- Holbrook, M. Adventures in Complexity: An Essay on Dynamic Open Complex Adaptive Systems, Butterfly Effects, Self-Organizing Order, Coevolution, the Ecological Perspective, Fitness Landscapes, Market Spaces, Emergent Beauty at the Edge of Chaos, and All That Jazz. Acad. Mark. Sci. Rev. 2003, 2003, 1–181. [Google Scholar]
- Fenner, M.W. Seed Ecology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; ISBN 978-94-009-4844-0. [Google Scholar]
- Putz, F.E. SILVICULTURE | Treatments in Tropical Silviculture. In Encyclopedia of Forest Sciences; Burley, J., Ed.; Elsevier: Oxford, UK, 2004; pp. 1039–1044. ISBN 978-0-12-145160-8. [Google Scholar]
- Kuuluvainen, T.; Lindberg, H.; Vanha-Majamaa, I.; Keto-Tokoi, P.; Punttila, P. Low-Level Retention Forestry, Certification, and Biodiversity: Case Finland. Ecol. Processes 2019, 8, 47. [Google Scholar] [CrossRef]
- Raye, J. Fractal Organisation Theory. J. Organ. Transform. Soc. Change 2014, 11, 50–68. [Google Scholar] [CrossRef]
- Shono, K.; Chazdon, R.; Bodin, B.; Wilson, S.; Durst, P. Assisted Natural Regeneration: Harnessing Nature for Restoration. Unasylva 2020, 252, 71–81. [Google Scholar]
- Yamamoto, S. Gap Regeneration of Major Tree Species in Different Forest Types of Japan. Vegetatio 1996, 127, 203–213. [Google Scholar] [CrossRef]
- Ning, Z.; Hong, J.; Yong-yan, J. A Phenology Study on the Common Tree Species of Natural Secondary Forests in Northeast China. Chin. J. Plant Ecol. 1990, 14, 336. [Google Scholar]
- Ulft, L. Regeneration in Natural and Logged Tropical Rain Forest-Modelling Seed Dispersal and Regeneration of Tropical Trees in Guyana; Tropenbos-Guyana Series 12; Tropenbos International: Georgetown, Guyana, 2004; ISBN 90-5113-076-7. [Google Scholar]
- Clark, D.A.; Clark, D.B. Life History Diversity of Canopy and Emergent Trees in a Neotropical Rain Forest. Ecol. Monogr. 1992, 62, 315–344. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Whitmore, T.C. Canopy Gaps and the Two Major Groups of Forest Trees. Ecology 1989, 70, 536–538. [Google Scholar] [CrossRef]
- Franklin, J. Regeneration and Growth of Pioneer and Shade-tolerant Rain Forest Trees in Tonga. N. Z. J. Bot. 2003, 41, 669–684. [Google Scholar] [CrossRef]
- Кoлесникoв, Б.П. Генетический этап в леснoй типoлoгии и егo задачи. Лесoведение 1974, 2, 3–20. [Google Scholar]
- Birks, H. Late-Quaternary Biotic Changes in Terrestrial and Lacustrine Environments, with Particular Reference to North-West Europe. In Handbook of Holocene palaeoecology and palaeohydrology; Berglund, B.E., Ed.; Wiley-Interscience; John Wiley & Sons Ltd.: Chichester, UK, 1986. [Google Scholar]
- Birks, H. Contributions of Quaternary Botany to Modern Ecology and Biogeography. Plant Ecol. Divers. 2019, 12, 189–385. [Google Scholar] [CrossRef]
- Navasaitis, M.; Ozolinčius, R.; Smaliukas, D.; Balevičienė, J.M. Lietuvos Dendroflora: Monografija; Lutute: Kaunas, Lithuania, 2003; ISBN 978-9955-575-35-1. [Google Scholar]
- Luo, C.; Liu, Y.; Shen, Z.; Yang, K.; Wang, X.; Jiang, Y. Modifying Regeneration Strategies Classification to Enhance the Understanding of Dominant Species Growth in Fire-Prone Forest in Southwest China. For. Ecosyst. 2022, 9, 100009. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Guariguata, M.R. Natural Regeneration as a Tool for Large-Scale Forest Restoration in the Tropics: Prospects and Challenges. Available online: https://www.cifor.org/knowledge/publication/6282/ (accessed on 19 April 2022).
- Storfer, A.; Murphy, M.A.; Evans, J.S.; Goldberg, C.S.; Robinson, S.; Spear, S.F.; Dezzani, R.; Delmelle, E.; Vierling, L.; Waits, L.P. Putting the ‘Landscape’ in Landscape Genetics. Heredity 2007, 98, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Forest Strategy. Available online: https://ec.europa.eu/environment/strategy/forest-strategy_en (accessed on 7 April 2022).
- Biodiversity Strategy for 2030. Available online: https://ec.europa.eu/environment/strategy/biodiversity-strategy-2030_en (accessed on 7 April 2022).
- Alia, R.; Aravanopoulos, F.; Fjellstad, K.B.; Bozzano, M.; Fady, B.; Farsakoglou, A.-M.; Gonzáles Martinez, S.C.; Heinze, B.; Kandemir, G.; Koziol, C.; et al. Forest Genetic Resources Strategy for Europe; EUFORGEN Secretariat, European Forest Institute: Barcelona, Spain, 2021; ISBN 978-952-7426-48-7. [Google Scholar]
- Xiong, H.; Choe, Y. Dynamical Pathway Analysis. BMC Syst. Biol. 2008, 2, 9. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Forest Habitat Types (NATURA 2000 Codes) | Main Forest Types, i.e., Field Layer-Canopy Dominants | Forest Site Types * | Soil Types ** |
|---|---|---|---|
| Mixed broadleaved forests (9020 9080 91F0 91E0 | Aegopodio-Quercetum | Nf, Lf | CM, LV |
| Carico-mixtoherbo-Fraxinetum | Lf | CM, LV | |
| Calamagrostido-Betuletum pubescentis | Uc | GL | |
| Filipendulo-Alnetum glutinosae | Ud | GL | |
| Urtico-Alnetum glutinosae | Uf | GL | |
| Carico-irido-Alnetum glutinosae | Pd | HSs-ph-ef | |
| Carico-Betuletum pubescentis | Pc | HSs-ph-mf | |
| Mixed Norway spruce forests (9050 9160 9180 9190 9070) | Oxalido-Piceetum | Nc | CM, LV, PL, AB, AR, FL |
| Myrtillo-oxalido-Piceetum | Lc | CM, LV, PL, AB, AR | |
| Hepatico-oxalido-Quercetum | Nc, Nd | CM, LV, PL, AB, AR, FL | |
| Oxalido-nemoroso-Piceetum | Ld | CM, LV, FL | |
| Scots pine forests (9010 9060 91D0 91T0) | Cladonio-Pinetum | Na | RG, AR |
| Vaccinio-Pinetum | Na, Nb | RG, AR, PZ | |
| Vaccinio-myrtillo-Pinetum | Nb | AR, PZ | |
| Myrtillo-Pinetum | Lb | AR, PL, PZ | |
| Myrtillo-sphagno-Pinetum | Ub | GL | |
| Carico-sphagno-Pinetum | Pb | HSf-s | |
| Ledo-sphagno-Pinetum | Pa | HSf |
| Growth | Establishment | |
|---|---|---|
| Forest | Gaps | |
| Forest | Expansion (A)—competitive stress-tolerators (C-S): Fagus sylvatica, Tilia cordata. Advanced regeneration under shade and grows best in forest stands; average growth rates, especially as juveniles (1). | Occupation (C)—competitive stress-tolerant ruderals (C-S-R): Fraxinus excelsior, Populus tremula, Quercus robur, Ulmus laevis. Regenerates and grows best in gaps, saplings can survive in closed forests; increased juvenile growth potential over groups A or B (3). |
| Gaps | Invasion (B)—competitive ruderals (C-R): Acer platanoides, Carpinus betulus, Picea abies, Ulmus glabra. Regenerates in shade but shows heightened association with gaps as saplings; growth rates are as low as group A but increase with size (2). | Colonization (D)—stress-tolerant ruderals (S-R): Alnus glutinosa, Alnus incana, Betula pendula, Betula pubescens, Larix decidua, Pinus sylvestris. Regenerates after gap formation and achieves optimal growth at all juvenile stages; juveniles have the highest growth potential (4). |
| Regeneration Strategies of Tree Species: C—Colonization, O—Occupation, I—Invasion, E—Expansion | Dynamic Forest Habitat Types (NATURA 2000 Codes)/Field Layer Codes of the Forest Type Series * | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gap Phase Dynamics with Mixed Broadleaved Forests on Rich Sites | Successional Development in Mixed Norway Spruce Forests on Mesic Sites | Multi-Cohort Succession in Scots Pine Forests on Poor Sites | |||||||||||||||||
| (9020 9080 91F0 91E0) | (9050 9160 9180 9190 9070) | (9010 9060 91D0 91T0) | |||||||||||||||||
| aeg * | cmh | cal | fil | ur | cir | c | ox | mox | hox | oxn | cl | v | vm | m | msp | csp | lsp | ||
| Alnus glutinosa | C | x | X | X | X | X | X | x | |||||||||||
| Alnus incana | C | X | x | x | x | x | x | X | X | ||||||||||
| Betula pendula | C | x | x | x | x | x | X | X | X | X | x | x | X | ||||||
| Betula pubescens | C | x | x | X | X | x | x | X | x | X | |||||||||
| Larix decidua | C | X | |||||||||||||||||
| Pinus sylvestris | C | x | X | X | X | X | X | X | X | X | X | ||||||||
| Fraxinus excelsior | O | X | X | x | x | X | |||||||||||||
| Populus tremula | O | x | x | x | x | X | x | X | x | ||||||||||
| Quercus robur | O | X | x | x | X | x | |||||||||||||
| Ulmus laevis | O | X | x | x | |||||||||||||||
| Acer platanoides | I | x | x | ||||||||||||||||
| Carpinus betulus | I | x | X | ||||||||||||||||
| Picea abies | I | x | x | X | X | X | X | x | x | x | |||||||||
| Ulmus glabra | I | X | |||||||||||||||||
| Fagus sylvatica | E | X | |||||||||||||||||
| Tilia cordata | E | X | X | ||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrokas, R.; Kavaliauskas, D. Concept for Genetic Monitoring of Hemiboreal Tree Dynamics in Lithuania. Land 2022, 11, 1249. https://doi.org/10.3390/land11081249
Petrokas R, Kavaliauskas D. Concept for Genetic Monitoring of Hemiboreal Tree Dynamics in Lithuania. Land. 2022; 11(8):1249. https://doi.org/10.3390/land11081249
Chicago/Turabian StylePetrokas, Raimundas, and Darius Kavaliauskas. 2022. "Concept for Genetic Monitoring of Hemiboreal Tree Dynamics in Lithuania" Land 11, no. 8: 1249. https://doi.org/10.3390/land11081249