
Influence of Leguminous Cover Crops on Soil Chemical and Biological Properties in a No-Till Tropical Fruit Orchard


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Location and Characteristics
2.2. Experimental Design
2.3. Field Sampling and Laboratory Analyses
2.3.1. Field Methodology
2.3.2. Soil Physicochemical Properties
2.3.3. Soil Enzyme Activity
2.3.4. Statistical Analyses
3. Results and Discussion
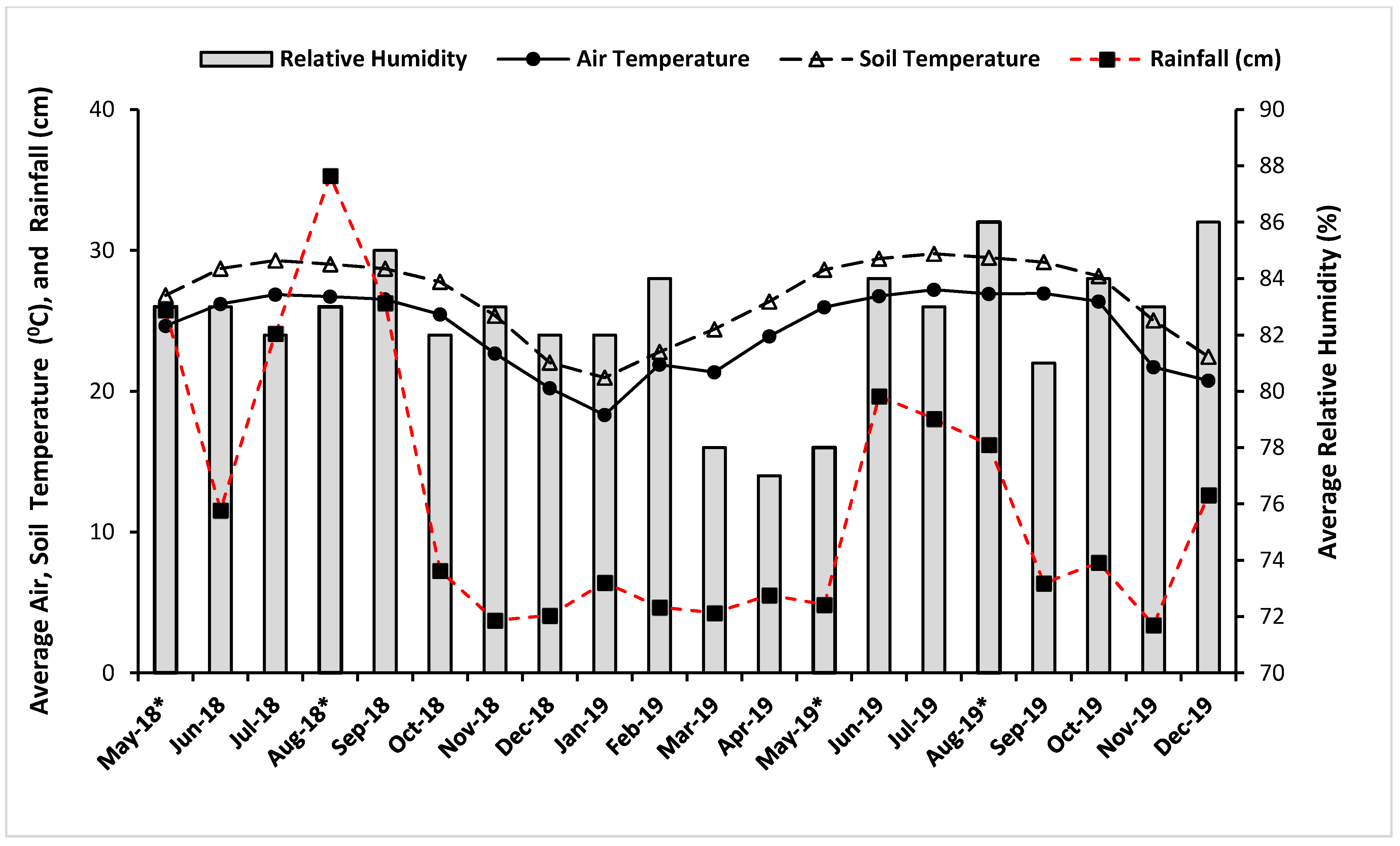
3.1. Background Soil Characteristics, Climatic Conditions, and Cover Crop Contributions
3.2. Effect of Cover Crops on Soil Chemical Properties
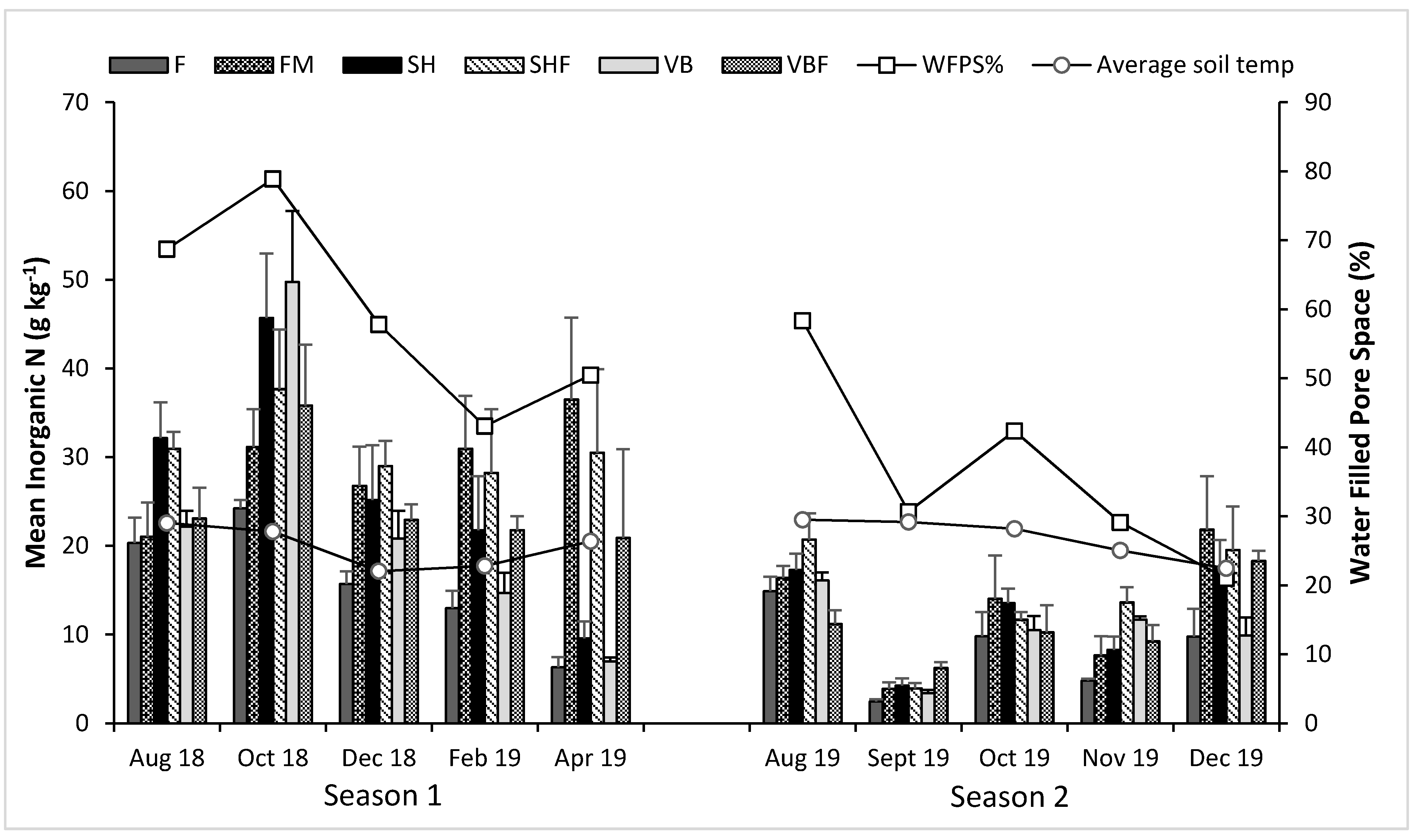
3.3. Soil Inorganic Nitrogen
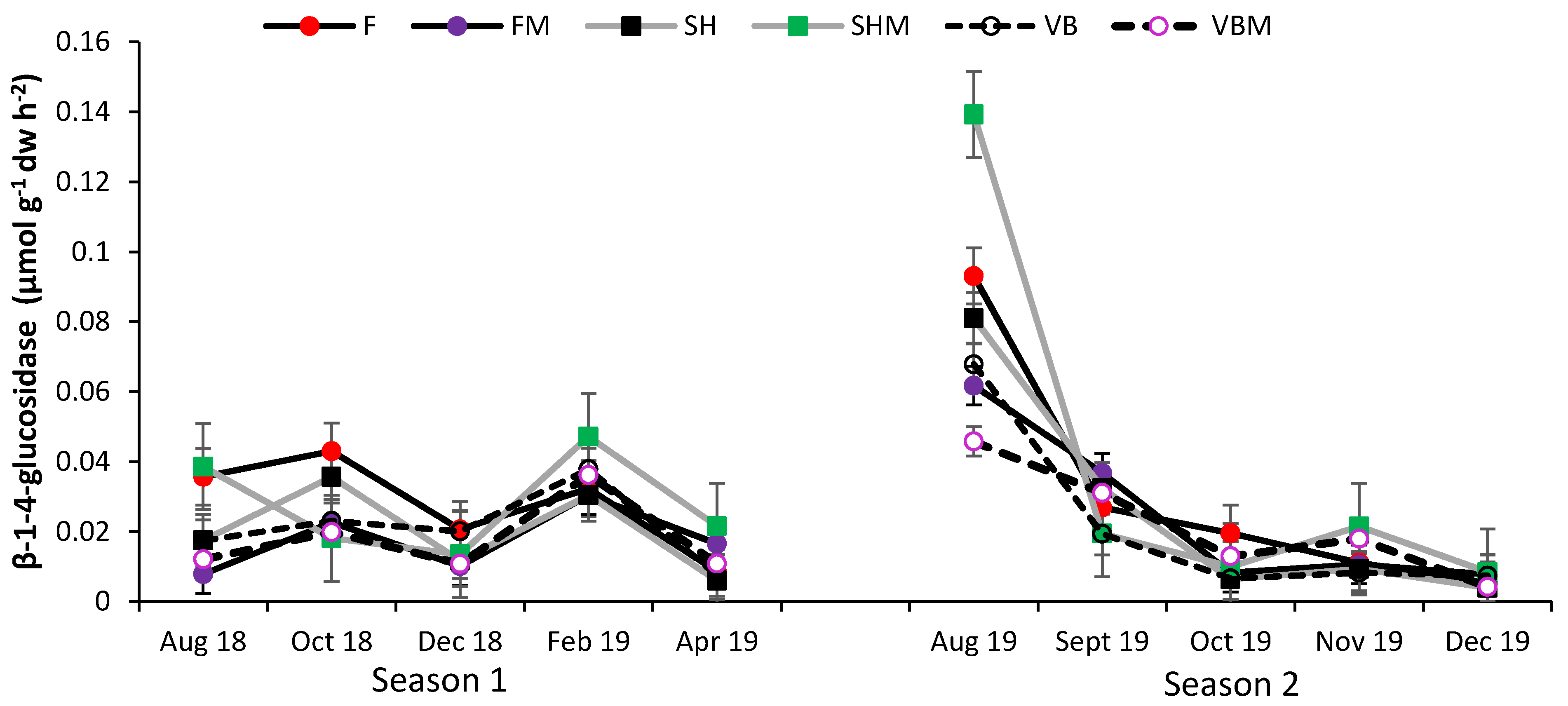
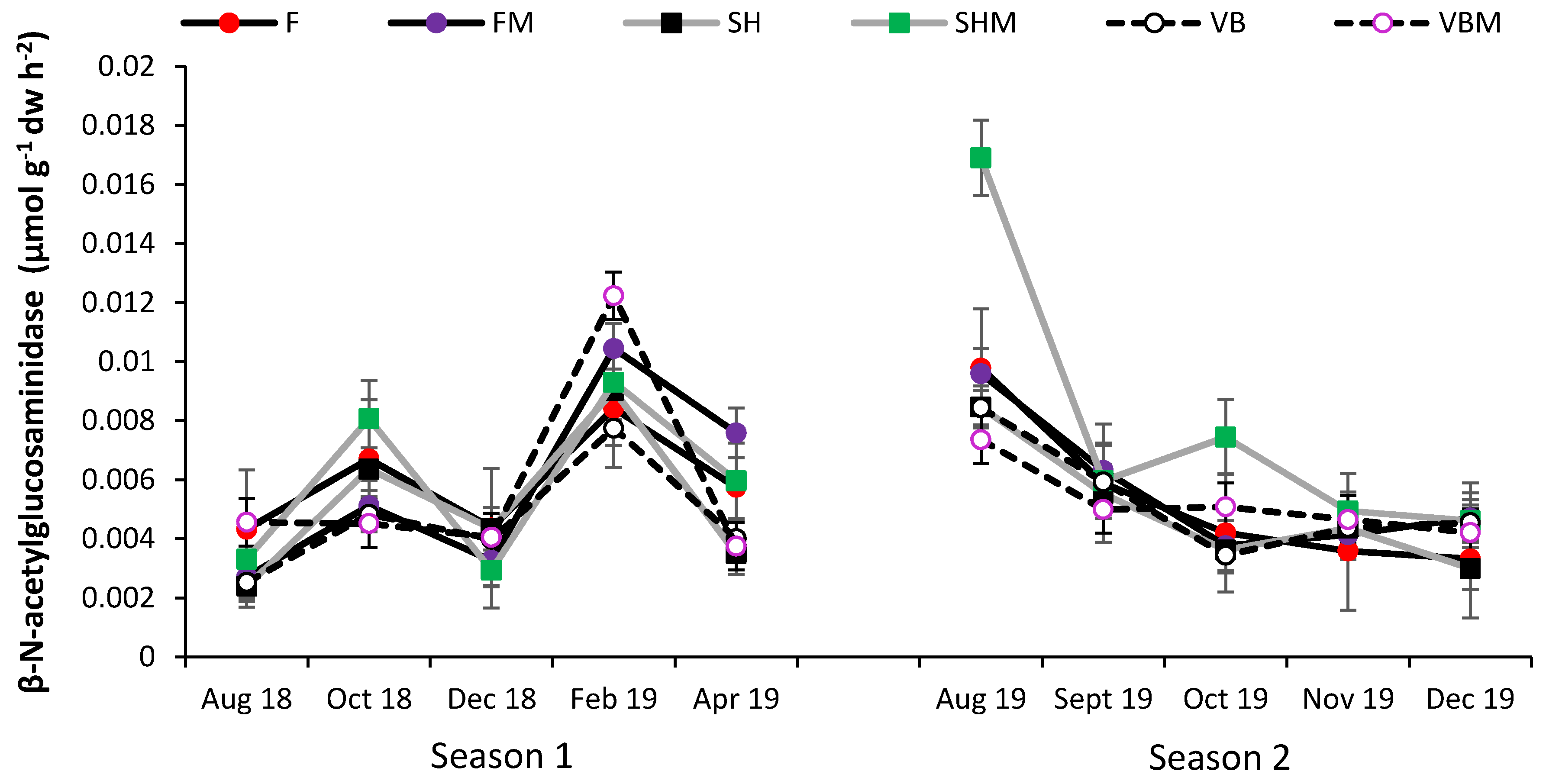
3.4. Soil Enzyme Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Inorganic N (g kg−1) | |||||||
|---|---|---|---|---|---|---|---|
| Season 1 | Overall | F | FM | SH | SHM | VB | VBM |
| August 2018 | 25.48 B | 20.32 c | 21.04 bc | 32.15 a | 30.94 ab | 22.16 bc | 23.08 abc |
| October 2018 | 38.26 A | 24.22 b | 31.15 ab | 45.70 ab | 37.62 ab | 49.74 a | 35.82 ab |
| December 2018 | 23.42 B | 15.71 b | 26.76 ab | 25.18 ab | 29.00 a | 20.83 ab | 22.93 ab |
| February 2019 | 20.90 B C | 12.96 b | 30.94 a | 21.76 ab | 28.23 ab | 14.71 ab | 21.77 ab |
| April 2019 | 17.58 C | 6.32 b | 36.49 a | 9.55 b | 30.50 a | 6.97 b | 20.90 ab |
| Season 2 | |||||||
| August 2019 | 16.30 CD | 14.89 ab | 16.39 ab | 17.28 ab | 20.70 a | 16.10 ab | 11.19 b |
| September 2019 | 4.03 F | 2.45 b | 3.90 b | 4.23 b | 3.94 b | 3.41 b | 6.23 a |
| October 2019 | 11.54 DE | 9.81 a | 14.02 a | 13.56 a | 11.67 a | 10.51 a | 10.24 a |
| November 2019 | 9.25 EF | 4.82 c | 7.65 bc | 8.27 bc | 13.63 a | 11.69 ab | 9.24 abc |
| December 2019 | 16.36 CD | 9.76 a | 21.83 a | 17.43 a | 19.52 a | 9.87 a | 18.30 a |
| SOM a | TC b | TN c | C:N | Inorganic N d | MUFN e | MUFC f | Moisture g | |
|---|---|---|---|---|---|---|---|---|
| pH | −0.326 ** i | −0.165 * | −0.315 ** | 0.299 ** | −0.511 ** | −0.273 ** | −0.171 ** | −0.240 ** |
| SOM | 0.690 ** | 0.764 ** | −0.541 ** | 0.506 ** | 0.081 | 0.130* | 0.294 ** | |
| TC | 0.740 ** | −0.353 ** | 0.404 ** | −0.034 | 0.060 | 0.234 ** | ||
| TN | −0.783 ** | 0.532 ** | 0.055 | 0.124 | 0.410 ** | |||
| C:N | −0.346 ** | −0.101 | −0.141 | −0.268 ** | ||||
| Inorganic N | 0.243 ** | 0.212 ** | 0.613 ** | |||||
| MUFN | 0.430 ** | 0.313 ** | ||||||
| MUFC | 0.214 ** |
References
- Camarotto, C.; Dal Ferro, N.; Piccoli, I.; Polese, R.; Furlan, L.; Chiarini, F.; Morari, F. Conservation agriculture and cover crop practices to regulate water, carbon and nitrogen cycles in the low-lying venetian plain. Catena 2018, 167, 236–249. [Google Scholar] [CrossRef]
- Kassam, A.; Friedrich, T.; Derpsch, R. Global spread of conservation agriculture. Int. J. Environ. Stud. 2019, 76, 29–51. [Google Scholar] [CrossRef]
- Hobbs, P.R.; Sayre, K.; Gupta, R. The role of conservation agriculture in sustainable agriculture. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, B.; Ghimire, R.; VanLeeuwen, D.; Mesbah, A. Cover crop residue amount and quality effects on soil organic carbon mineralization. Sustainability 2017, 9, 2316. [Google Scholar] [CrossRef] [Green Version]
- Horowitz, J.K.; Ebel, R.M.; Ueda, K. “No-Till” Farming Is a Growing Practice; Report No. 1476-2016-120976; USDA: Washington, DC, USA, 2010.
- Vukicevich, E.; Lowery, T.; Bowen, P.; Úrbez-Torres, J.R.; Hart, M. Cover crops to increase soil microbial diversity and mitigate decline in perennial agriculture. A review. Agron. Sustain. Dev. 2016, 36, 48. [Google Scholar] [CrossRef] [Green Version]
- Bayer, C.; Dieckow, J.; Amado, T.J.C.; Eltz, F.L.F.; Vieira, F.C.B. Cover crop effects increasing carbon storage in a subtropical no-till sandy acrisol. Commun. Soil Sci. Plant Anal. 2009, 40, 1499–1511. [Google Scholar] [CrossRef]
- Chen, Q.; Sun, Y.; Shen, C.; Peng, S.; Yi, W.; Li, Z.; Jiang, M. Organic matter turnover rates and CO2 flux from organic matter decomposition of mountain soil profiles in the subtropical area, south China. Catena 2002, 49, 217–229. [Google Scholar] [CrossRef]
- Tang, J.; Qi, Y.; Xu, M.; Misson, L.; Goldstein, A.H. Forest thinning and soil respiration in a ponderosa pine plantation in the sierra nevada. Tree Physiol. 2005, 25, 57–66. [Google Scholar] [CrossRef]
- Curiel Yuste, J.; Baldocchi, D.D.; Gershenson, A.; Goldstein, A.; Misson, L.; Wong, S. Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture. Glob. Chang. Biol. 2007, 13, 2018–2035. [Google Scholar] [CrossRef] [Green Version]
- Abang, A.F.; Kouamé, C.M.; Abang, M.; Hanna, R.; Fotso, A.K. Assessing vegetable farmer knowledge of diseases and insect pests of vegetable and management practices under tropical conditions. Int. J. Veg. Sci. 2014, 20, 240–253. [Google Scholar] [CrossRef] [Green Version]
- Vincent, C.; Schaffer, B.; Rowland, D.L.; Migliaccio, K.W.; Crane, J.H.; Li, Y. Sunn hemp intercrop and mulch increases papaya growth and reduces wind speed and virus damage. Sci. Hortic. 2017, 218, 304–315. [Google Scholar] [CrossRef]
- Ramos, M.G.; Villatoro, M.A.A.; Urquiaga, S.; Alves, B.J.; Boddey, R.M. Quantification of the contribution of biological nitrogen fixation to tropical green manure crops and the residual benefit to a subsequent maize crop using 15N-isotope techniques. J. Biotechnol. 2001, 91, 105–115. [Google Scholar] [CrossRef]
- Perin, A.; Santos, R.H.S.; Urquiaga, S.S.; Cecon, P.R.; Guerra, J.G.M.; Freitas, G.B.d. Sunnhemp and millet as green manure for tropical maize production. Sci. Agric. 2006, 63, 453–459. [Google Scholar] [CrossRef]
- Rigon, J.; Franzluebbers, A.J.; Calonego, J.C. Soil aggregation and potential carbon and nitrogen mineralization with cover crops under tropical no-till. J. Soil Water Conserv. 2020, 75, 601–609. [Google Scholar] [CrossRef]
- Oliveira, F.C.C.; Ferreira, G.W.D.; Souza, J.L.S.; Vieira, M.E.O.; Pedrotti, A. Soil physical properties and soil organic carbon content in northeast brazil: Long-term tillage systems effects. Sci. Agric. 2019, 77. [Google Scholar] [CrossRef] [Green Version]
- Comin, J.J.; Ferreira, L.B.; dos Santos, L.H.; de Paula Koucher, L.; Machado, L.N.; dos Santos, E., Jr.; Mafra, Á.L.; Kurtz, C.; Souza, M.; Brunetto, G.; et al. Carbon and nitrogen contents and aggregation index of soil cultivated with onion for seven years using crop successions and rotations. Soil Tillage Res. 2018, 184, 195–202. [Google Scholar] [CrossRef]
- Calonego, J.C.; Raphael, J.P.; Rigon, J.P.; de Oliveira Neto, L.; Rosolem, C.A. Soil compaction management and soybean yields with cover crops under no-till and occasional chiseling. Eur. J. Agron. 2017, 85, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Nannipieri, P.; Kandeler, E.; Ruggiero, P. Enzyme activities and microbiological and biochemical processes in soil. In Enzymes in the Environment; Marcel Dekker: New York, NY, USA, 2002; pp. 1–33. [Google Scholar]
- Hamido, S.A.; Kpomblekou-A, K. Cover crop and tillage effects on soil enzyme activities following tomato. Soil Tillage Res. 2009, 105, 269–274. [Google Scholar] [CrossRef]
- Willer, H.; Lernoud, J. The World of Organic Agriculture: Statistics and Emerging Trends 2019; Research Institute of Organic Agriculture FiBL: Frick, Switzerland; IFOAM Organics International: Bonn, Germany, 2019. [Google Scholar]
- USDA National Agricultural Statistics Service. Census of Agriculture; USDA National Agricultural Statistics Service: Washington, DC, USA, 2017.
- USDA Agricultural Marketing Service. Organic Integrity Database. 2022. Available online: https://organic.ams.usda.gov/integrity/ (accessed on 16 May 2022).
- Li, Y. Calcareous Soils in Miami Dade County. Fact Sheet 183. Soil and Water Science Department, Florida Coorperative Extension Services, University of Florida. 2001. Available online: Http://Edis.Ifas.Ufl.Edu/Pdffiles/TR/TR00400.Pdf (accessed on 1 October 2007).
- Farhad, W.; Saleem, M.F.; Cheema, M.A.; Hammad, H.M. Effect of poultry manure levels on the productivity of spring maize (Zea mays L.). J. Anim. Plant Sci. 2009, 19, 122–125. [Google Scholar]
- Hochmuth, G.; Hochmuth, R.; Mylavarapu, R. Using composted poultry manure (litter) in mulched vegetable production. EDIS 2009, 2009. [Google Scholar] [CrossRef]
- Nyakatawa, E.Z.; Reddy, K.C.; Sistani, K.R. Tillage, cover cropping, and poultry litter effects on selected soil chemical properties. Soil Tillage Res. 2001, 58, 69–79. [Google Scholar] [CrossRef]
- Reddy, C.K.; Nyakatawa, E.Z.; Reeves, D.W. Tillage and poultry litter application effects on cotton growth and yield. Agron. J. 2004, 96, 1641–1650. [Google Scholar] [CrossRef] [Green Version]
- Núñez-Elisea, R.; Crane, J.H. Selective pruning and crop removal increase early-season fruit production of carambola (Averrhoa carambola L.). Sci. Hortic. 2000, 86, 115–126. [Google Scholar] [CrossRef]
- Ballen, F.H.; Evans, E.; Crane, J.; Singh, A. Sample profitability and cost estimates of producing sweet flavored carambola (Averrhoa carambola) in south florida. EDIS 2020, 2020, 7. [Google Scholar] [CrossRef]
- Crane, J.H.; Wasielewski, J. Tropical Fruit Acerage in Florida. UF/IFAS Extension Miami Dade County. 2018. Available online: https://sfyl.ifas.ufl.edu/media/sfylifasufledu/miami-dade/documents/tropical-fruit/Tropical-Fruit-Acreage.pdf (accessed on 16 May 2022).
- Menocal, O.; Kendra, P.E.; Montgomery, W.S.; Crane, J.H.; Carrillo, D. Vertical distribution and daily flight periodicity of ambrosia beetles (coleoptera: Curculionidae) in florida avocado orchards affected by laurel wilt. J. Econ. Entomol. 2018, 111, 1190–1196. [Google Scholar] [CrossRef] [PubMed]
- Menocal, O.; Kendra, P.E.; Padilla, A.; Chagas, P.C.; Chagas, E.A.; Crane, J.H.; Carrillo, D. Influence of Canopy Cover and Meteorological Factors on the Abundance of Bark and Ambrosia Beetles (Coleoptera: Curculionidae) in Avocado Orchards Affected by Laurel Wilt. Agronomy 2022, 12, 547. [Google Scholar] [CrossRef]
- Carrillo, D.; Duncan, R.E.; Peña, J.E. Ambrosia beetles (coleoptera: Curculionidae: Scolytinae) that breed in avocado wood in florida. Fla. Entomol. 2012, 95, 573–579. [Google Scholar] [CrossRef]
- Evans, E.; Wasielewski, J.; Crane, J.H. Monetary Value of Tropical and Sub-Tropical Fruit Trees in US Dollars, UF/IFAS Extension Miami-Dade County. 2017. Available online: https://sfyl.ifas.ufl.edu/media/sfylifasufledu/miami-dade/documents/disaster-preparation/post-hurricane-and-disaster/TropicalFruitTreeWorthFactsheet2017.pdf (accessed on 16 May 2022).
- USDA Plant Hardiness Zone Map. Agricultural Research Service, U.S. Department of Agriculture. 2012. Available online: https://planthardiness.ars.usda.gov (accessed on 16 May 2022).
- Kwon, H.; Lall, U.; Obeysekera, J. Simulation of daily rainfall scenarios with interannual and multidecadal climate cycles for south florida. Stoch. Environ. Res. Risk Assess. 2009, 23, 879–896. [Google Scholar] [CrossRef]
- Wang, Q.; Klassen, W.; Li, Y.; Codallo, M. Cover crops and organic mulch to improve tomato yields and soil fertility. Agron. J. 2009, 101, 345–351. [Google Scholar] [CrossRef]
- Crane, J.H.; Balerdi, C.F. Preparing for and recovering from hurricane and tropical storm damage to tropical fruit groves in Florida1. EDIS 2006, 2006. [Google Scholar]
- Freidenreich, A.; Dattamudi, S.; Li, Y.C.; Jayachandran, K. Soil Respiration and Carbon Balance Under Cover Crop in a no-Till Tropical Fruit Orchard. Front. Environ. Sci. 2021, 9, 653. [Google Scholar] [CrossRef]
- Crane, J.H. The Carambola (Star Fruit). University of Florida Cooperative Extension Service, Institute of Food and Agriculture Sciences, EDIS 2001. Available online: https://www.doc-developpement-durable.org/file/Culture/Arbres-Fruitiers/FICHES_ARBRES/Carambolier/Carambola%20Star%20Fruit.pdf (accessed on 12 October 2020).
- USEPA—US Environmental Protection Agency. Method 350.1. Determination of Ammonia Nitrogen by Semi-Automated Colorimetry; Environmental Monitoring Systems Laboratory, Office of Research and Development: Cincinnati, OH, USA, 1993.
- USEPA—US Environmental Protection Agency. Method 353.2. Determination of Nitrate Nitrite Nitrogen by Automated Colorimetry; Environmental Monitoring Systems Laboratory, Office of Research and Development: Cincinnati, OH, USA, 1993.
- Sinsabaugh, R.L.; Findlay, S.; Franchini, P.; Fischer, D. Enzymatic analysis of riverine bacterioplankton production. Limnol. Oceanogr. 1997, 42, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Hoppe, H. Use of fluorogenic model substrates for extracellular enzyme activity (EEA) measurement of bacteria. In Handbook of Methods in Aquatic Microbial Ecology; CRC Press: Boca Raton, FL, USA, 1993; pp. 423–431. [Google Scholar]
- Chróst, R.J.; Krambeck, H. Fluorescence correction for measurements of enzyme activity in natural waters using methylumbelliferyl-substrates. Arch. Hydrobiol. 1986, 106, 79–90. [Google Scholar]
- Xiong, Y.; Zeng, H.; Xia, H.; Guo, D. Interactions between leaf litter and soil organic matter on carbon and nitrogen mineralization in six forest litter-soil systems. Plant Soil 2014, 379, 217–229. [Google Scholar] [CrossRef]
- Sheahan, C.M. Plant Guide for Sunn Hemp (Crotalaria Juncea). USDA-Natural Resources Conservation Service, Cape May Plant Materials Center, Cape May, NJ, 8210. 2012. Available online: https://www.nrcs.usda.gov/Internet/FSE_PLANTMATERIALS/publications/njpmcpg11706.pdf (accessed on 16 June 2020).
- Baligar, V.C.; Fageria, N.K. Agronomy and physiology of tropical cover crops. J. Plant Nutr. 2007, 30, 1287–1339. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Klassen, W. Summer cover crops and soil amendments to improve growth and nutrient uptake of okra. HortTechnology 2006, 16, 328–338. [Google Scholar] [CrossRef] [Green Version]
- Wayman, S.; Cogger, C.; Benedict, C.; Collins, D.; Burke, I.; Bary, A. Cover crop effects on light, nitrogen, and weeds in organic reduced tillage. Agroecol. Sustain. Food Syst. 2015, 39, 647–665. [Google Scholar] [CrossRef]
- Romanyà, J.; Casals, P. Biological nitrogen fixation response to soil fertility is species-dependent in annual legumes. J. Soil Sci. Plant Nutr. 2019, 20, 546–556. [Google Scholar] [CrossRef]
- Garcia, R.A.; Li, Y.; Rosolem, C.A. Soil organic matter and physical attributes affected by crop rotation under no-till. Soil Sci. Soc. Am. J. 2013, 77, 1724–1731. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Wang, R.; Cai, J.; Zhang, Y.; Li, H.; Huang, S.; Jiang, Y. Impacts of fertilization practices on pH and the pH buffering capacity of calcareous soil. Soil Sci. Plant Nutr. 2016, 62, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Janzen, H.H.; McGinn, S.M. Volatile loss of nitrogen during decomposition of legume green manure. Soil Biol. Biochem. 1991, 23, 291–297. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Alva, A. Cover crops in mono-and biculture for accumulation of biomass and soil organic carbon. J. Sustain. Agric. 2012, 36, 423–439. [Google Scholar] [CrossRef]
- Eiland, F.; Klamer, M.; Lind, A.M.; Leth, M.; Bååth, E. Influence of initial C/N ratio on chemical and microbial composition during long term composting of straw. Microb. Ecol. 2001, 41, 272–280. [Google Scholar] [CrossRef]
- Leghari, S.J.; Wahocho, N.A.; Laghari, G.M.; HafeezLaghari, A.; MustafaBhabhan, G.; HussainTalpur, K.; Bhutto, T.A.; Wahocho, S.A.; Lashari, A. Role of nitrogen for plant growth and development: A review. Adv. Environ. Biol. 2016, 10, 209–219. [Google Scholar]
- Dias, T.; Dukes, A.; Antunes, P.M. Accounting for soil biotic effects on soil health and crop productivity in the design of crop rotations. J. Sci. Food Agric. 2015, 95, 447–454. [Google Scholar] [CrossRef]
- Rao, R.B.; Li, Y.C. Nitrogen mineralization of cover crop residues in calcareous gravelly soil. Commun. Soil Sci. Plant Anal. 2003, 34, 299–313. [Google Scholar] [CrossRef]
- Linn, D.M.; Doran, J.W. Effect of water-filled pore space on carbon dioxide and nitrous oxide production in tilled and nontilled soils. Soil Sci. Soc. Am. J. 1984, 48, 1267–1272. [Google Scholar] [CrossRef] [Green Version]
- Ball, B.C.; Crichton, I.; Horgan, G.W. Dynamics of upward and downward N2O and CO2 fluxes in ploughed or no-tilled soils in relation to water-filled pore space, compaction and crop presence. Soil Tillage Res. 2008, 101, 20–30. [Google Scholar] [CrossRef]
- Linn, D.M.; Doran, J.W. Aerobic and anaerobic microbial populations in no-till and plowed soils. Soil Sci. Soc. Am. J. 1984, 48, 794–799. [Google Scholar] [CrossRef] [Green Version]
- Clagnan, E.; Thornton, S.F.; Rolfe, S.A.; Wells, N.S.; Knoeller, K.; Murphy, J.; Tuohy, P.; Daly, K.; Healy, M.G.; Ezzati, G.; et al. An integrated assessment of nitrogen source, transformation and fate within an intensive dairy system to inform management change. PLoS ONE 2019, 14, e0219479. [Google Scholar] [CrossRef]
- Liu, X.J.; Mosier, A.R.; Halvorson, A.D.; Reule, C.A.; Zhang, F.S. Dinitrogen and N2O emissions in arable soils: Effect of tillage, N source and soil moisture. Soil Biol. Biochem. 2007, 39, 2362–2370. [Google Scholar] [CrossRef]
- Dobbie, K.E.; Smith, K.A. The effects of temperature, water-filled pore space and land use on N2O emissions from an imperfectly drained gleysol. Eur. J. Soil Sci. 2001, 52, 667–673. [Google Scholar] [CrossRef]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.; et al. Complete nitrification by nitrospira bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Carey, C.J.; Dove, N.C.; Beman, J.M.; Hart, S.C.; Aronson, E.L. Meta-analysis reveals ammonia-oxidizing bacteria respond more strongly to nitrogen addition than ammonia-oxidizing archaea. Soil Biol. Biochem. 2016, 99, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Zhou, G.; Rees, R.M.; Cao, W. Green manuring inhibits nitrification in a typical paddy soil by changing the contributions of ammonia-oxidizing archaea and bacteria. Appl. Soil Ecol. 2020, 156, 103698. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, L.; Zhu, Y.; Zhang, J.; He, J. Abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea communities of an alkaline sandy loam. Environ. Microbiol. 2008, 10, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Norton, J.M. Microbial control of nitrate concentrations in an agricultural soil treated with dairy waste compost or ammonium fertilizer. Soil Biol. Biochem. 2000, 32, 1453–1457. [Google Scholar] [CrossRef]
- Yin, Z.; Bi, X.; Xu, C. Ammonia-oxidizing archaea (AOA) play with ammonia-oxidizing bacteria (AOB) in nitrogen removal from wastewater. Archaea 2018, 2018, 8429145. [Google Scholar] [CrossRef] [Green Version]
- Hungria, M.; Kaschuk, G. Regulation of N2 fixation and NO3−/NH4 assimilation in nodulated and N-fertilized phaseolus vulgaris L. exposed to high temperature stress. Environ. Exp. Bot. 2014, 98, 32–39. [Google Scholar] [CrossRef]
- Enrico, J.M.; Piccinetti, C.F.; Barraco, M.R.; Agosti, M.B.; Eclesia, R.P.; Salvagiotti, F. Biological nitrogen fixation in field pea and vetch: Response to inoculation and residual effect on maize in the pampean region. Eur. J. Agron. 2020, 115, 126016. [Google Scholar] [CrossRef]
- Nezomba, H.; Tauro, T.P.; Mtambanengwe, F.; Mapfumo, P. Nitrogen fixation and biomass productivity of indigenous legumes for fertility restoration of abandoned soils in smallholder farming systems. S. Afr. J. Plant Soil 2008, 25, 161–171. [Google Scholar] [CrossRef]
- Allen, O.N.; Allen, E.K. The Leguminosae: A Source Book of Characteristics, Uses, and Nodulation; University of Wisconsin Press: Madison, WI, USA, 1981. [Google Scholar]
- Li, T.; Gao, J.; Bai, L.; Wang, Y.; Huang, J.; Kumar, M.; Zeng, X. Influence of green manure and rice straw management on soil organic carbon, enzyme activities, and rice yield in red paddy soil. Soil Tillage Res. 2019, 195, 104428. [Google Scholar] [CrossRef]
- Chung, H.; Zak, D.R.; Reich, P.B.; Ellsworth, D.S. Plant species richness, elevated CO2, and atmospheric nitrogen deposition alter soil microbial community composition and function. Global Change Biology. 2007, 13, 980–989. [Google Scholar] [CrossRef] [Green Version]
- Maltais-Landry, G.; Scow, K.; Brennan, E. Soil phosphorus mobilization in the rhizosphere of cover crops has little effect on phosphorus cycling in california agricultural soils. Soil Biol. Biochem. 2014, 78, 255–262. [Google Scholar] [CrossRef]
- Henry, S.; Texier, S.; Hallet, S.; Bru, D.; Dambreville, C.; Chèneby, D.; Bizouard, F.; Germon, J.C.; Philippot, L. Disentangling the rhizosphere effect on nitrate reducers and denitrifiers: Insight into the role of root exudates. Environ. Microbiol. 2008, 10, 3082–3092. [Google Scholar] [CrossRef]





| Season 1 | Season 2 | |||||
|---|---|---|---|---|---|---|
| Treatment | Biomass (Mg ha−1) | N (kg ha−1) | Weed Biomass (kg ha−1) | Biomass (Mg ha−1) | N (kg ha−1) | Weed Biomass (kg ha−1) |
| F | - | - | 9475 a | - | - | 6496 a |
| FM | - | - | 7802 a | - | - | 6531 a |
| SH | 8.8 a | 177 ab | 1778 b | 9.4 a | 213 a | 1930 b |
| SHM | 10.7a | 238 a | 2322 b | 5.4 b | 135 b | 3302 b |
| VB | 4.5 b | 88 c | 3928 b | 2.7 c | 84 bc | 2742 b |
| VBM | 5.0 b | 136 bc | 2384 b | 1.9 c | 65 c | 3156 b |
| pH | SOM% | TC (g kg−1) | TN (g kg−1) | C:N (mol:mol) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | Season 1 | Season 2 | Season 1 | Season 2 | Season 1 | Season 2 | Season 1 | Season 2 | Season 1 | Season 2 |
| F | 8.03 a | 8.08 a | 15.00 bc | 13.99 b | 158.63 c | 159.81 c | 7.08 b | 7.10 bc | 26.55 a | 26.88 ab |
| FM | 7.75 b | 7.83 b | 17.86 a | 16.45 a | 183.63 a | 181.37 a | 9.17 a | 8.31 a | 23.38 a | 24.93 b |
| SH | 7.98 ab | 8.03 ab | 17.32 ab | 15.64 a | 175.32 ab | 169.67 abc | 8.95 a | 7.86 ab | 24.27 a | 25.18 ab |
| SHM | 7.85 ab | 7.99 ab | 17.45 a | 15.48 ab | 188.05 a | 170.79 abc | 9.25 a | 7.41 abc | 24.55 a | 27.37 ab |
| VB | 7.95 ab | 7.93 ab | 16.31 abc | 14.14 b | 178.11 ab | 173.89 ab | 7.70 ab | 6.55 c | 26.52 a | 28.10 a |
| VBM | 7.87 ab | 7.87 ab | 14.70 c | 15.48 ab | 166.42 bc | 168.94 bc | 7.89 ab | 6.98 bc | 25.38 a | 24.91 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freidenreich, A.; Dattamudi, S.; Li, Y.; Jayachandran, K. Influence of Leguminous Cover Crops on Soil Chemical and Biological Properties in a No-Till Tropical Fruit Orchard. Land 2022, 11, 932. https://doi.org/10.3390/land11060932
Freidenreich A, Dattamudi S, Li Y, Jayachandran K. Influence of Leguminous Cover Crops on Soil Chemical and Biological Properties in a No-Till Tropical Fruit Orchard. Land. 2022; 11(6):932. https://doi.org/10.3390/land11060932
Chicago/Turabian StyleFreidenreich, Ariel, Sanku Dattamudi, Yuncong Li, and Krishnaswamy Jayachandran. 2022. "Influence of Leguminous Cover Crops on Soil Chemical and Biological Properties in a No-Till Tropical Fruit Orchard" Land 11, no. 6: 932. https://doi.org/10.3390/land11060932