
Net Ecosystem Exchange of Carbon Dioxide in Rice-Spring Wheat System of Northwestern Indo-Gangetic Plains

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Site Description
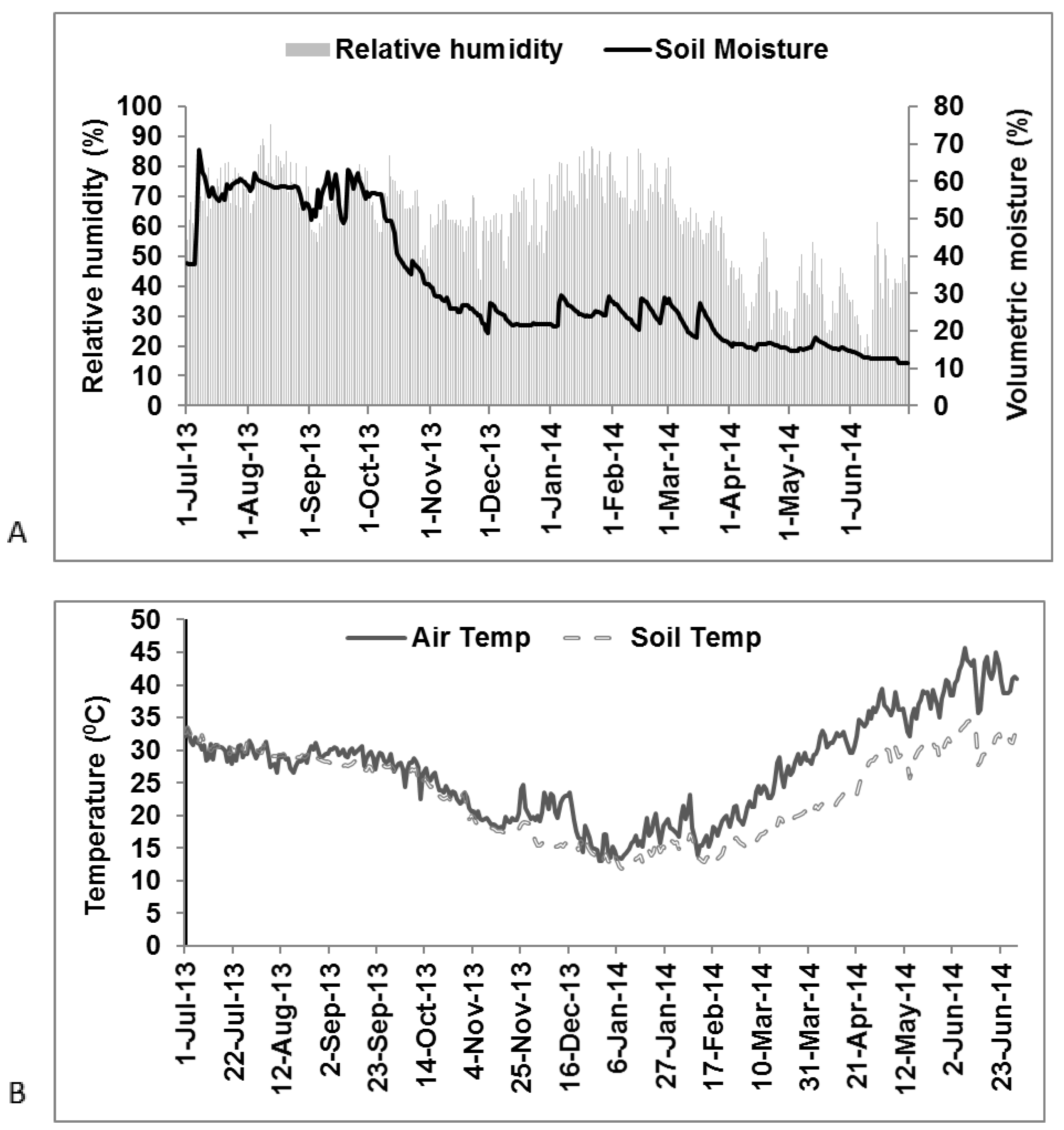
2.2. Climatic Description
2.3. Soil Properties
2.4. Crop Management
2.5. Eddy Covariance Measurements
2.6. Auxiliary Measurements
2.7. Quality Control of Flux Data
2.8. Partitioning NEE into GPP and RE
3. Results
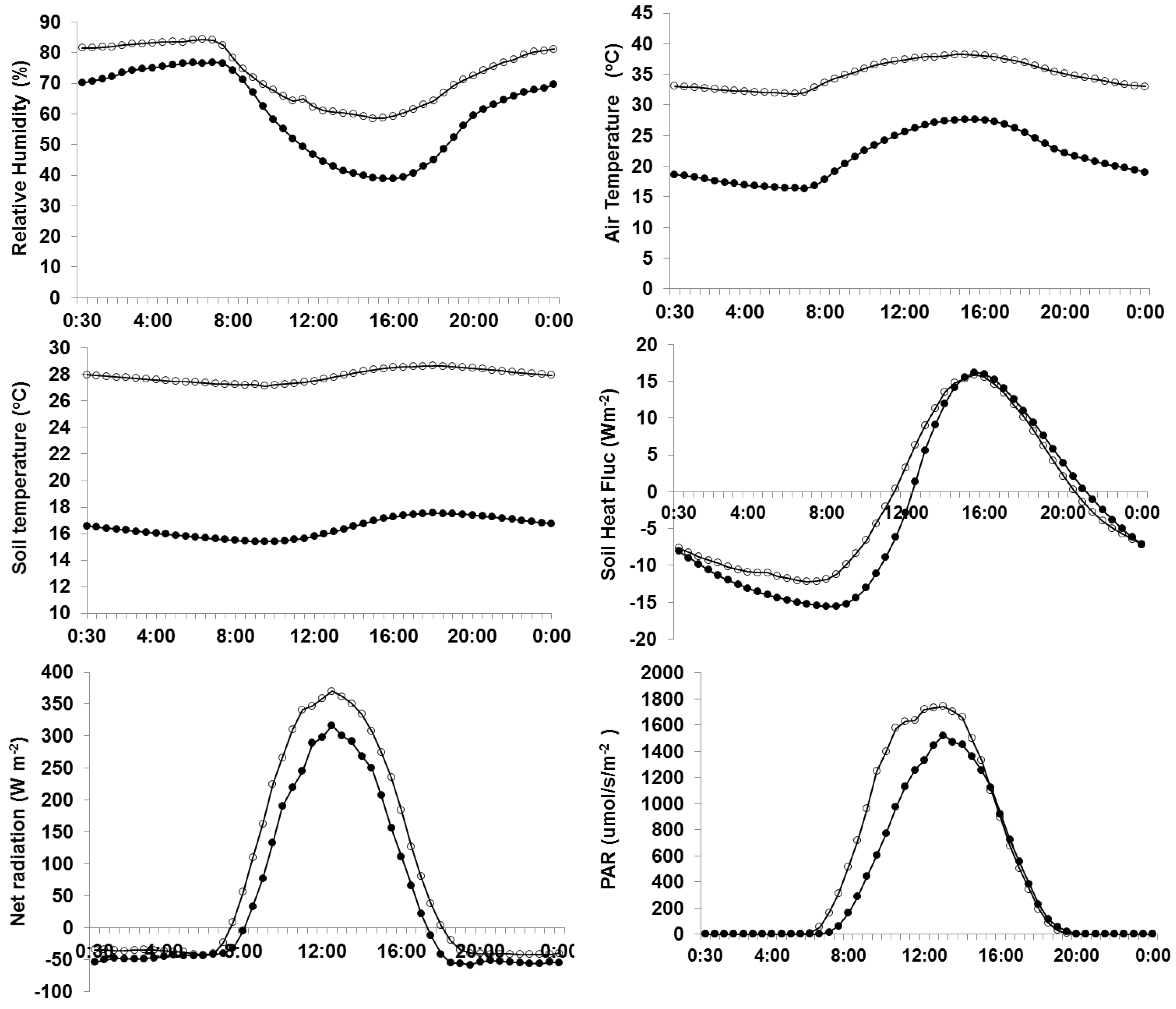
3.1. Climatic Variability during Rice and Wheat Growing Season
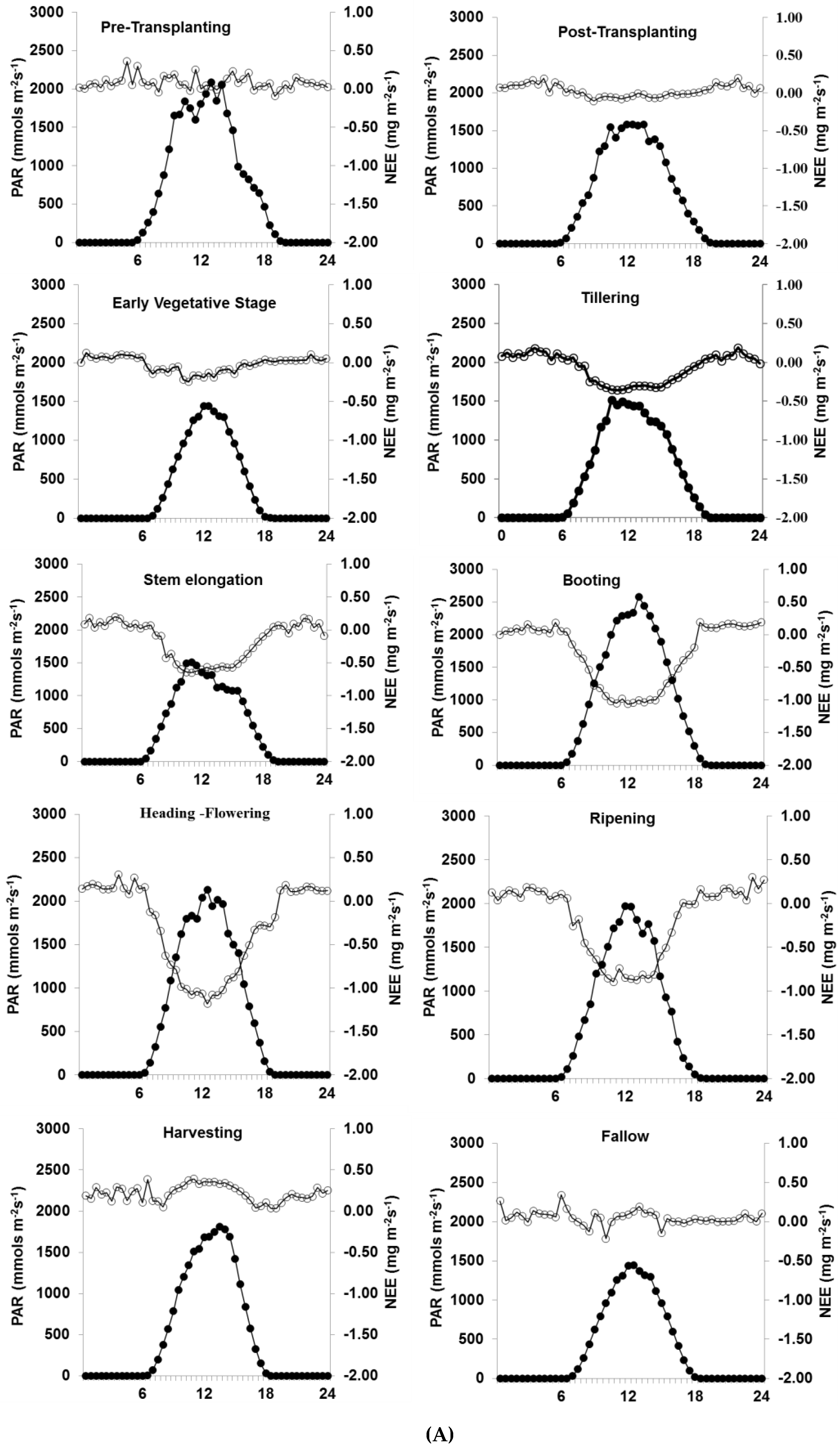
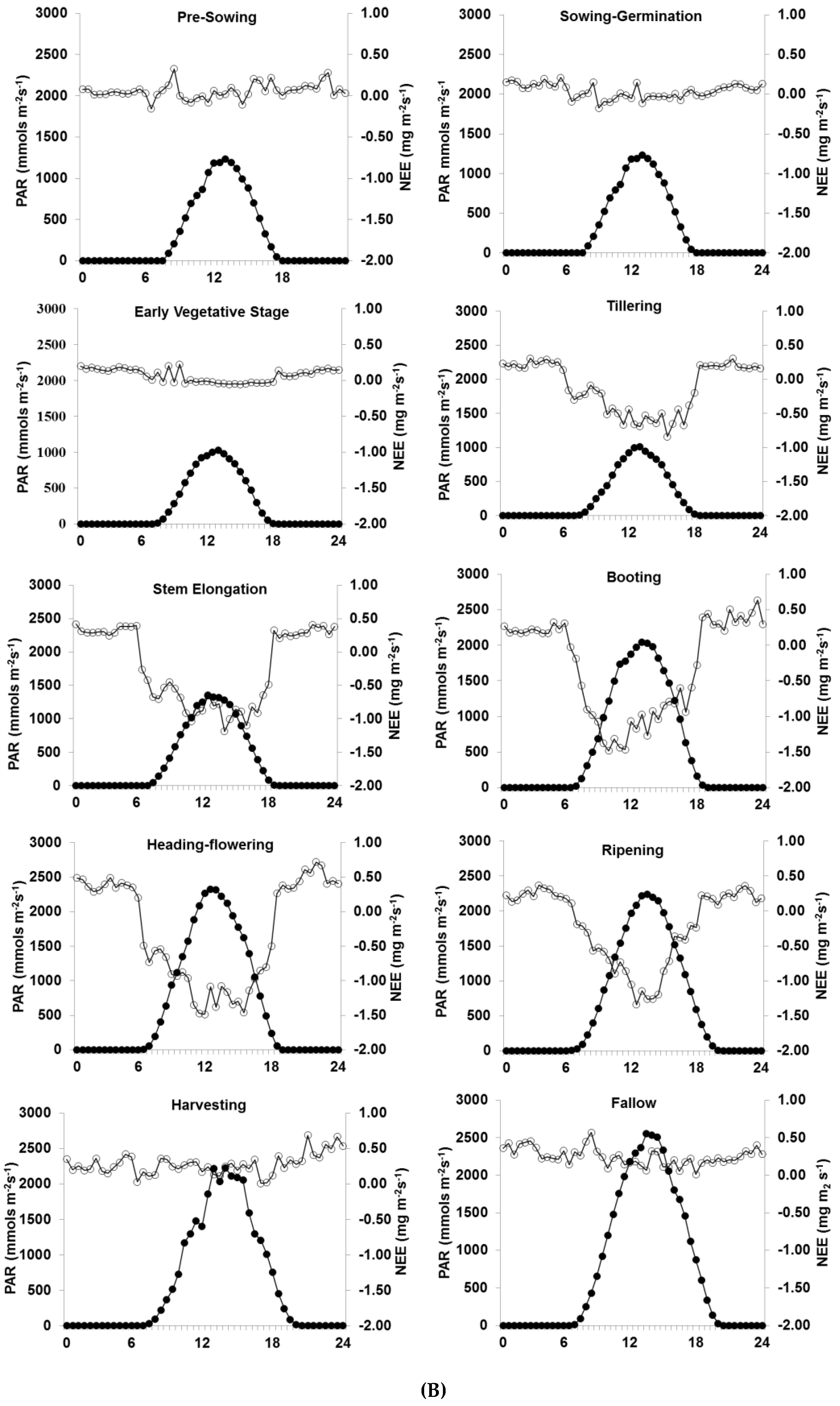
3.2. Diurnal Variation in NEE
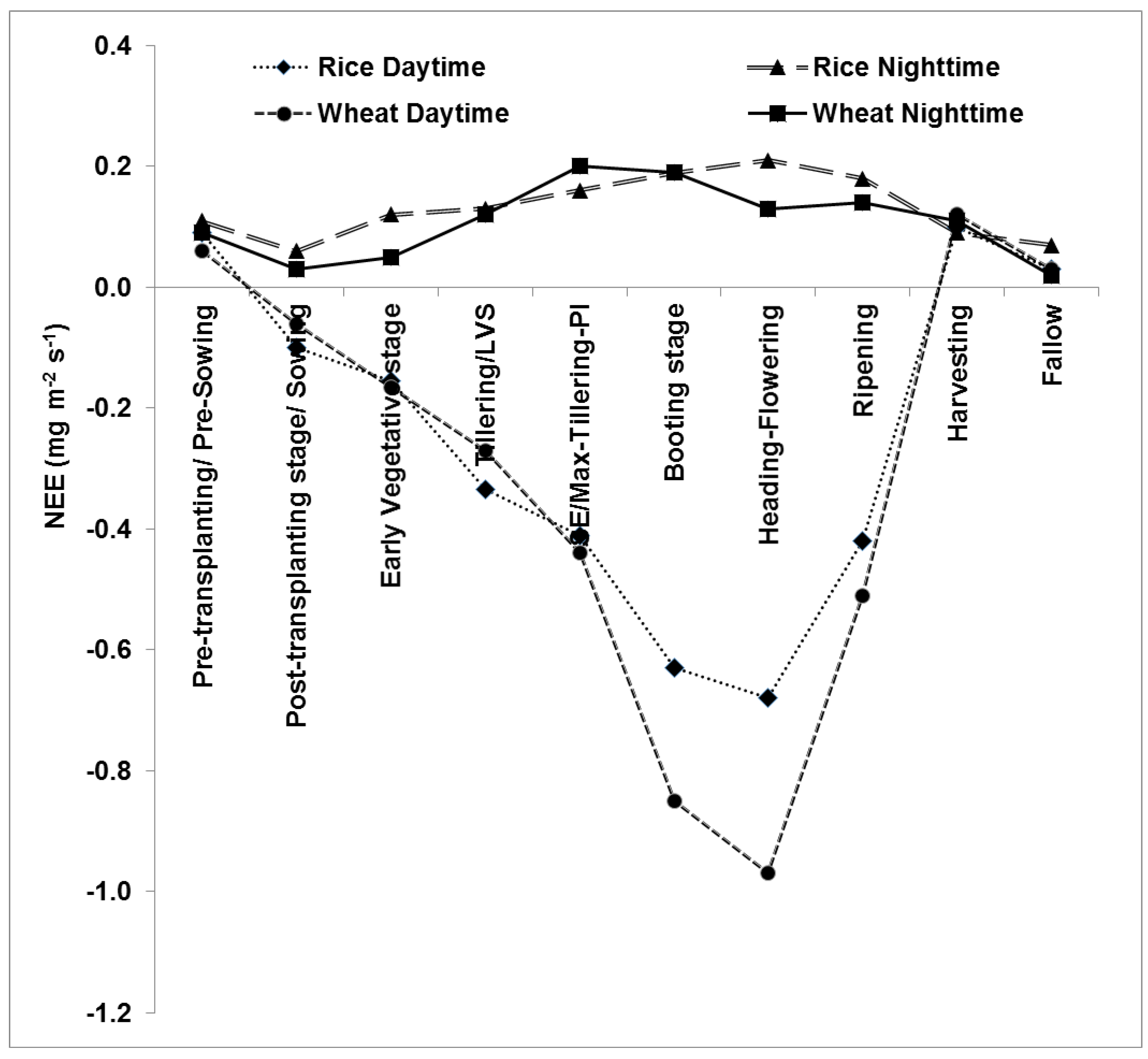
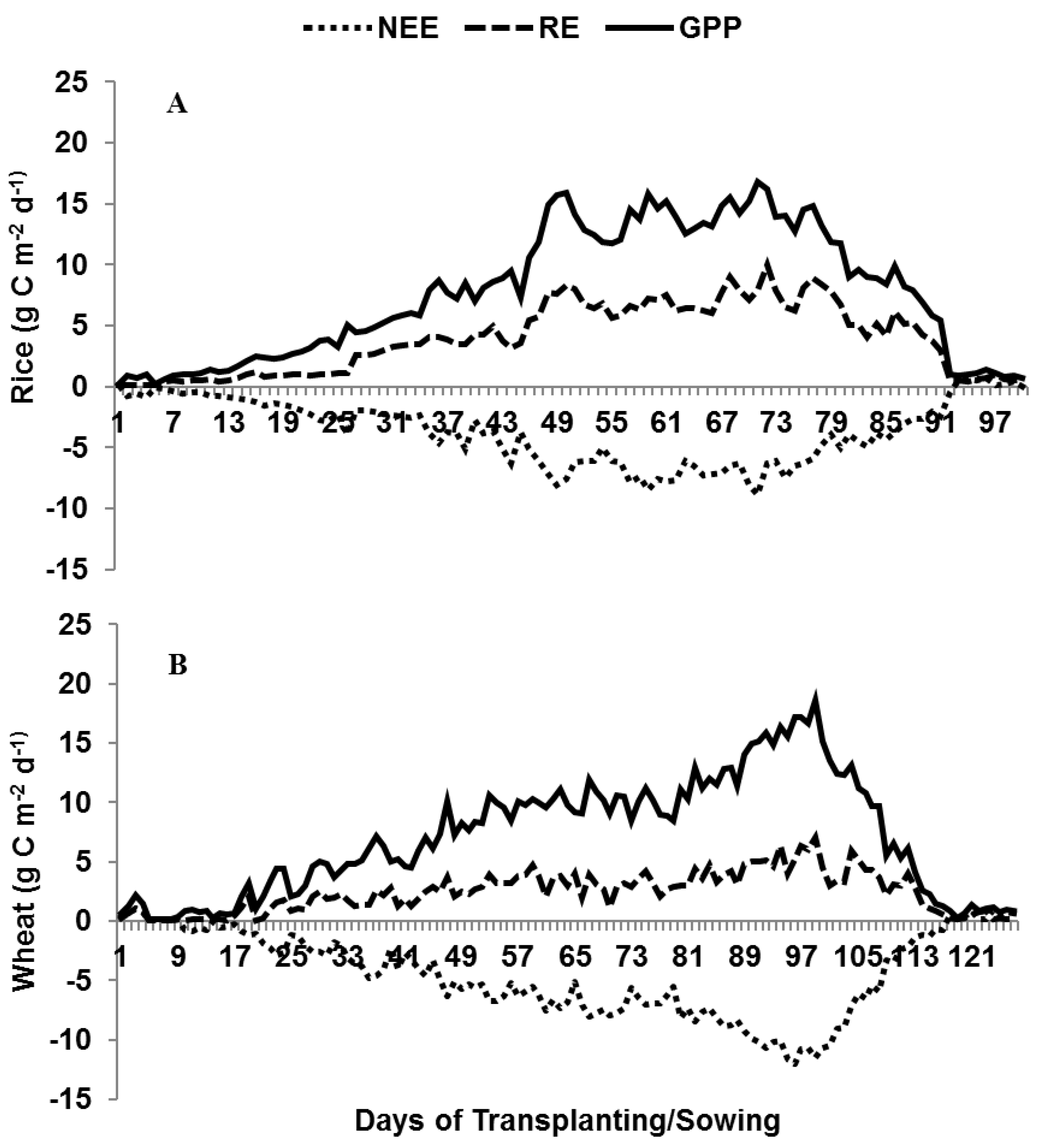
3.3. Seasonal Variation in Daily NEE
3.4. RE and GPP at Different Crop Growth Stages
3.5. Environmental Variables in Rice and Wheat
4. Discussion
4.1. Diurnal and Seasonal Variation in NEE
4.2. Effect of Crop Growth, LAI and Air Temperature on GPP
4.3. Effect of NR, PAR, SM, RH and VPD on GPP
4.4. Effect of Crop Stage, Environmental Variables on RE
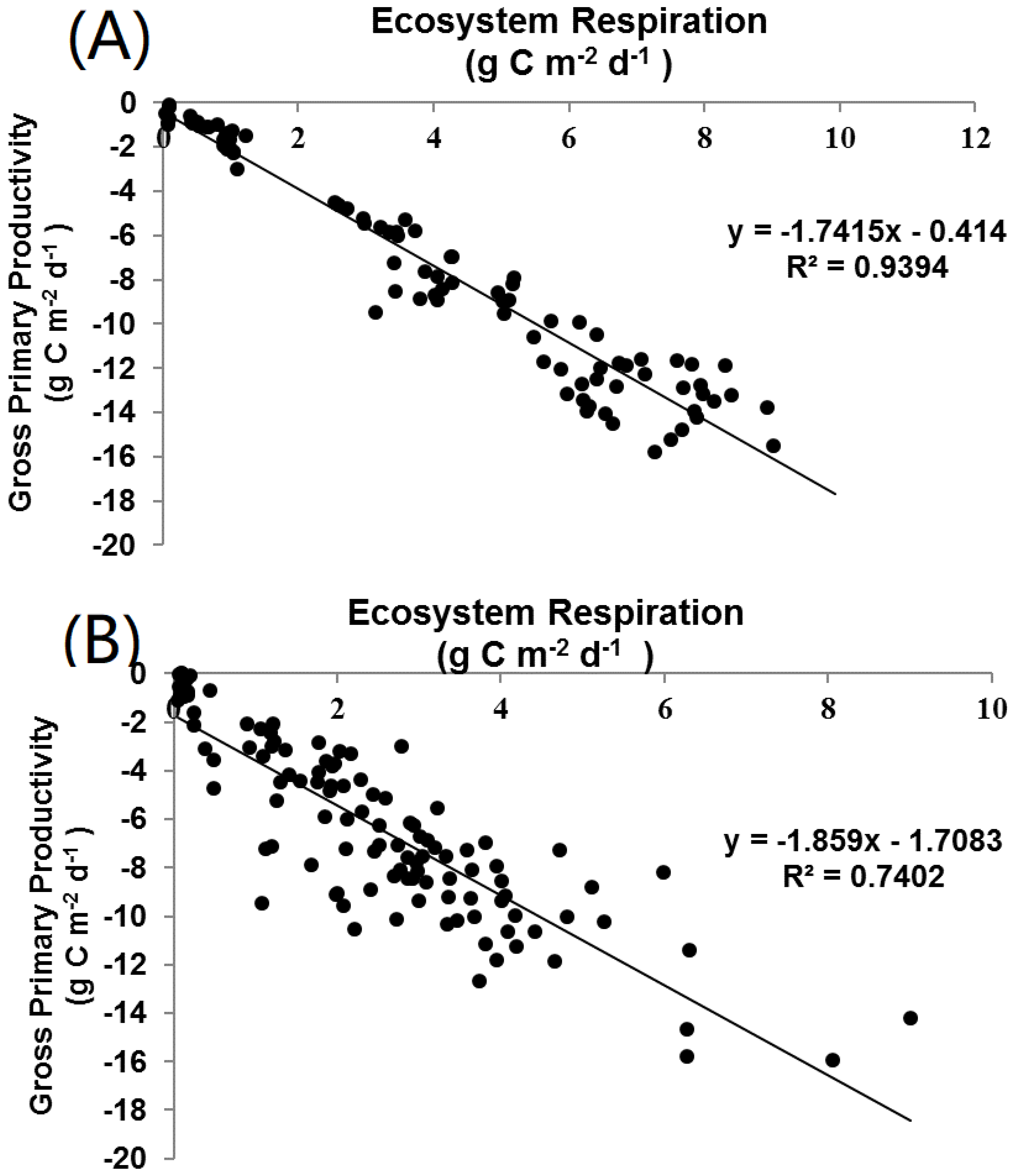
4.5. Relation between GPP and RE
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bhatia, A.; Kumar, A.; Kumar, V.; Jain, N. Low carbon option for sustainable agriculture. Ind. Farm. 2013, 63, 18–22. [Google Scholar]
- IPCC, Climate Change. The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; p. 1535. [Google Scholar]
- Kumar, A.; Bhatia, A.; Fagodiya, R.K.; Malyan, S.K.; Meena, B.L. Eddy Covariance Flux Tower: A Promising Technique for Greenhouse Gases Measurement. Adv. Plants Agric. Res. 2017, 7, 00263. [Google Scholar]
- Bhattacharyya, R.; Bhatia, A.; Das, T.K.; Lata, S.; Kumar, A.; Tomer, R.; Singh, G. Aggregate-associated N and global warming potential of conservation agriculture-based cropping of maize-wheat system in the north-western Indo-Gangetic Plains. Soil Till. Res. 2018, 182, 66–77. [Google Scholar] [CrossRef]
- Kumar, A.; Tomer, R.; Bhatia, A.; Jain, N.; Pathak, H. Greenhouse Gas Mitigation in Indian Agriculture in Agro-Technologies for Adaptation to Climate Change; Pathak, H., Chakrabarti, B., Eds.; CESCRA, IARI: New Delhi, India, 2015. [Google Scholar]
- Kumar, A.; Sharma, M.P.; Taxak, A.K. Effect of Vegetation Communities and Altitudes on the Soil Organic Carbon Stock in Kotli Bhel-1A Catchment, India. CLEAN Soil Air Water 2017, 45, 1600650. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, M.P. Carbon stock estimation in the catchment of Kotli Bhel 1A hydroelectric reservoir, Uttarakhand, India. Ecotoxicol. Environ. Saf. 2016, 134, 365–369. [Google Scholar] [CrossRef]
- Kumar, A.; Yang, T.; Sharma, M.P. Greenhouse gas measurement from Chinese freshwater bodies: A review. J. Clean. Prod. 2019, 233, 368–378. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, M.P.; Yang, T. Estimation of carbon stock for greenhouse gas emissions from hydropower reservoirs. Stoch. Environ. Res. Risk Assess. 2018. [Google Scholar] [CrossRef]
- Sándor, R.; Ehrhardt, F.; Grace, P.; Recous, S.; Smith, P.; Snow, V.; Soussana, J.-F.; Basso, B.; Bhatia, A.; Brilli, L.; et al. Ensemble modelling of carbon fluxes in grasslands and croplands. Field Crop Res. 2020, 252, 10779. [Google Scholar] [CrossRef]
- Ladha, J.K.; Pathak, H.; Tirol-Padre, A.; Dawe, D. Productivity trends in intensive rice–wheat cropping systems in Asia. In Productivity and Sustainability of Rice–Wheat Systems: Issues and Impacts; ASA Spec. Publ. 65; ASA, CSSA, and SSSA: Madison, WI, USA, 2003; pp. 45–76. [Google Scholar]
- Chen, C.; Dan, L.I.; Zhiqiu, G.; Tang, J.; Xiaofeng, G.; Linlin, W.; Bingcheng, W. Seasonal and interannual variations of carbon exchange over a rice–wheat rotation system on the north china plain. Advan. Atmos. Sci. 2015, 32, 1365–1380. [Google Scholar] [CrossRef]
- Gupta, D.K.; Bhatia, A.; Kumar, A.; Chakrabarti, B.; Jain, N.; Pathak, H. Global warming potential of rice–wheat cropping system of the IGP. Ind. J. Agric Sci. 2015, 85, 807–816. [Google Scholar]
- Bhatia, A.; Pathak, H.; Aggarwal, P.K.; Jain, N. Trade-off between productivity enhancement and global warming potential of rice and wheat in India. Nutr. Cycl. Agroecosys. 2010, 86, 413–424. [Google Scholar] [CrossRef]
- Gupta, D.K.; Bhatia, A.; Das, T.K.; Singh, P.; Kumar, A.; Jain, N.; Pathak, H. Economic analysis of different greenhouse gas mitigation technologies in rice–wheat cropping system of the IGP. Curr. Sci. 2016, 110, 867–874. [Google Scholar]
- Gupta, D.K.; Bhatia, A.; Kumar, A.; Das, T.K.; Jain, N.; Tomar, R.; Malyan, S.K.; Fagodiya, R.K.; Dubey, R.; Pathak, H. Mitigation of greenhouse gas emission from rice–wheat system of the IGP: Through tillage, irrigation and fertilizer management. Agric. Ecosyst. Environ. 2016, 230, 1–9. [Google Scholar] [CrossRef]
- Sage, R.F.; Kubien, D.S. The temperature response of C3 and C4 photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef]
- Kramer, K.; Leinonen, I.; Bartelink, H.H.; Berbigier, P.; Borghetti, M.; Bernhofer, C.; Cienciala, E.; Dolman, A.J.; Froer, O.; Gracia, C.A.; et al. Evaluation of six process-based forest growth models using eddy-covariance measurements of CO2 and H2O fluxes at six forest sites in Europe. Global Chang. Biol. 2002, 8, 213–230. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Neogi, S.; Roy, K.S.; Dash, P.K.; Tripathi, R.; Rao, K.S. Net ecosystem CO2 exchange and carbon cycling in tropical lowland flooded-rice ecosystem. Nutr. Cycl. Agroecosys. 2013, 95, 133–144. [Google Scholar] [CrossRef]
- Alberto, M.C.R.; Hirano, T.; Miyata, A.; Wassmann, R.; Kumar, A.; Padre, A.; Amante, M. Influence of climate variability on seasonal and inter-annual variations of ecosystem CO2 exchange in flooded and non-flooded rice fields in the Philippines. Field Crops Res. 2012, 134, 80–94. [Google Scholar] [CrossRef]
- Tseng, K.H.; Tsai, J.L.; Alagesan, A.; Tsuang, B.J.; Yao, M.H.; Kuo, P.H. Determination of methane and CO2 fluxes during the rice maturity period in Taiwan by combining profile and eddy covariance measurements. Agric. For. Meteor. 2010, 150, 852–859. [Google Scholar] [CrossRef]
- Hobbs, P.R.; Gupta, R.K. Resource-Conserving Technologies for Wheat in Rice Wheat Systems; NRG Web Publication; CIMMYT: México-Veracruz, Mexico, 2001. [Google Scholar]
- Bhatia, A.; Agarwal, P.K.; Jain, N.; Pathak, H. Greenhouse gas emission from rice and wheat-growing areas in India: Spatial analysis and upscaling. Greenhouse Gas Sci. Technol. 2012, 2, 115–125. [Google Scholar] [CrossRef]
- Saito, M.; Miyata, A.; Nagai, H.; Yamada, T. Seasonal variation of CO2 exchange in rice paddy field in Japan. Agric. For. Meteor. 2005, 135, 93–109. [Google Scholar] [CrossRef]
- Webb, E.K.; Pearman, G.I.; Leuning, R. Correction of flux measurement for density effects due to heat and water vapour transfer. J. Roy Meteor. Soc. 1980, 106, 85–100. [Google Scholar] [CrossRef]
- Kaimal, J.C.; Finnigan, J.J. Atmospheric Boundary Layer Flows: Their Structure and Measurement; Oxford University Press: New York, NY, USA, 1994; Volume 289. [Google Scholar]
- Van Dijk, A.; Kohsiek, W.; de Bruin, H.A.R. The Principles of Surface Flux Physics: Theory, Practice and Description of the ECPACK Library; Meteorology and Air Quality Group, Wageningen University: Wageningen, The Netherlands, 2004. [Google Scholar]
- Fan, S.M.; Wofsy, S.C.; Bakwin, P.S. Atmosphere biosphere exchange of CO2 and O3 in the central Amazon forest. J. Geophys. Res. 1990, 95, 851–864. [Google Scholar]
- Foken, T.; Wichura, B. Tools for quality assessment of surface-based flux measurements. Agric. For. Meteor. 1996, 78, 83–105. [Google Scholar] [CrossRef]
- Zhang, X.; Fan, C.; Ma, Y. Two approaches for net ecosystem carbon budgets and soil carbon sequestration in a rice-wheat rotation system in China. Nutr. Cycl. Agroecosys. 2014, 100, 301–313. [Google Scholar] [CrossRef]
- Hirata, R.; Hirano, T.; Saigusa, N.; Fujinuma, Y.; Inukai, K.; Kitamori, Y.; Takahashi, Y.; Yamamoto, S. Seasonal and interannual variations in CO2 exchange of a temperate larch forest. Agric. For. Meteor. 2007, 147, 110–124. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology; The Benjamin/Cummings Publishing Co., Inc.: Redwood City, CA, USA, 1991; pp. 434–436. [Google Scholar]
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Aubinet, M.; Bernhofer, C.; Burba, G.; Ceulemans, R.; Clement, R.; Dolman, H. Gap-filling strategies for defensible annual sums of net ecosystem exchange. Agric. For. Meteor. 2001, 107, 43–69. [Google Scholar] [CrossRef] [Green Version]
- Alberto, M.C.R.; Wassmann, R.; Hirano, T.; Miyata, A.; Kumar, A.; Padre, A.; Amante, M. CO2/heat fluxes in rice fields: Comparative assessment of flooded and non-flooded fields in the Philippines. Agric. For. Meteor. 2009, 149, 1737–1750. [Google Scholar] [CrossRef]
- Das, D.K.; Mukherjee, J.; Vashisth, A.; Sehgal, V.K.; Singh, J. Agro-climatic Manual of Indian Agricultural Research Institute New Delhi. Division of Agricultural Physics; ICAR: New Delhi, India, 2015. [Google Scholar]
- Miyata, A.; Leuning, R.; Denmead, O.W.; Kim, J.; Harazano, Y. CO2 and methane fluxes from an intermittently-flooded paddy field. Agr. For. Meteor. 2000, 102, 287–303. [Google Scholar] [CrossRef]
- Gao, Z.; Bian, L.; Zhou, X. Measurements of turbulent transfer in the near-surface layer over a rice paddy in China. J. Geophys. Res. 2003, 108, 4387. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, R.; Xu, Z.; Qiao, C.; Jiang, G. Diurnal and Seasonal Variations in Carbon Dioxide Exchange in Ecosystems in the Zhangye Oasis Area, Northwest China. PLoS ONE 2015, 10, e0120660. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.S.; Heilman, J.L.; McInnes, K.J.; Wilson, L.T.; Medley, J.C.; Wu, G.W.; Cobos, D.R. Diel and seasonal variation in CO2 flux of irrigated-rice. Agric. For. Meteor. 2001, 108, 15–27. [Google Scholar] [CrossRef]
- Patel, N.R.; Dadhwal, V.K.; Saha, S.K. Measurement and Scaling of Carbon Dioxide Exchanges in Wheat Using Flux-Tower and Remote Sensing. J. Ind. Soc. Remote Sens. 2011, 39, 383–391. [Google Scholar] [CrossRef]
- Baker, J.M.; Griffis, T.J. Examining strategies to improve the carbon balance of corn/soybean agriculture using eddy covariance and mass balance techniques. Agric. Forest Meteorol. 2005, 128, 163–177. [Google Scholar] [CrossRef]
- Li, J.; Yu, Q.; Sun, X.M.; Tong, X.J.; Ren, C.Y.; Wang, J.; Liu, E.; Zhu, Z.; Yu, G. Carbon dioxide exchange and the mechanism of environmental control in a farmland ecosystem in North China plain. Sci. China Ser. D 2006, 49, 226–240. [Google Scholar] [CrossRef]
- Schmidt, M.; Reichenaua, T.G.; Fienerc, P.; Schneider, K. The carbon budget of a winter wheat field: An eddy covariance analysis of seasonal and inter-annual variability. Agric. For. Meteor. 2012, 165, 114–126. [Google Scholar] [CrossRef]
- Moureaux, C.; Debacq, A.; Bodson, B.; Heinesch, B.; Aubinet, M. Annual net ecosystem carbon exchange by a sugar beet crop. Agric. For. Meteorol. 2006, 139, 25–39. [Google Scholar] [CrossRef]
- Amthor, J.S. The McCree—de Wit—Penning de Vries—Thornley respiration paradigms: 30 years later. Ann. Bot. 2000, 86, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Béziat, P.; Ceschia, E.; Dedieu, G. Carbon balance of a three crop succession over two cropland sites in South West France. Agric. For. Meteor. 2009, 149, 1628–1645. [Google Scholar] [CrossRef] [Green Version]
- Gitelsona, A.A.; Penga, Y.; Arkebauer, T.J.; Suyker, A.E. Productivity absorbed photosynthetically active radiation, and light use efficiency in crops: Implications for remote sensing of crop primary production. J. Plant. Physiol. 2015, 177, 100–109. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Li, Z.; Xie, F. Environmental influences on light response parameters of net carbon exchange in two rotation croplands on the North China Plain. Sci. Rep. 2019, 9, 18702. [Google Scholar] [CrossRef]
- Shirke, P.A.; Pathre, U.V. Influence of leaf-to-air vapour pressure deficit (VPD) on the biochemistry and physiology of photosynthesis in Prosopis juliflora. J. Exp. Bot. 2004, 55, 2111–2120. [Google Scholar] [CrossRef]
- Baldocchi, D. A comparative-study of mass and energy-exchange rates over a closed C3 and an open C4 crop. Agric. For. Meteor. 1994, 67, 291–321. [Google Scholar] [CrossRef]
- Mielnik, P.C.; Dugas, W.A. Soil CO2 flux in a tall grass prairie. Soil Biol. Biochem. 2000, 32, 221–228. [Google Scholar] [CrossRef]
- Mowjood, M.I.M.; Ishiguro, K.; Kasubuchi, T. Effect of convection in ponded water on the thermal regime of a paddy field. Soil Sci. 1997, 162, 583–587. [Google Scholar] [CrossRef]
- Long, S.P. Modification of the response of photosynthetic productivity to rising temperature by atmospheric CO2 concentration: Has its importance been underestimated? Plant Cell Environ. 1991, 14, 729–739. [Google Scholar] [CrossRef]
- Atkin, O.K.; Edwards, E.J.; Loveys, B.R. Response of root-respiration to changes in temperature and its relevance to global warming. New. Phytol. 2008, 147, 141–154. [Google Scholar] [CrossRef]
- Zhang, Q.; Lei, H.; Yang, D.; Xiong, L.; Liu, P.; Fang, B. Decadal variation in CO2 fluxes and its budget in a wheat and maize rotation cropland over the North China Plain. Biogeosciences 2020, 17, 2245–2262. [Google Scholar] [CrossRef] [Green Version]
- Gifford, R.M. Whole plant respiration and photosynthesis of wheat under increased CO2 concentration and temperature: Long-term vs. short-term distinctions for modeling. Glob. Chang. Biol. 1995, 1, 385–396. [Google Scholar] [CrossRef]
- Rogelj, J.; Forster, P.M.; Kriegler, E.; Smith, C.J.; Séférian, R. Estimating and tracking the remaining carbon budget for stringent climate targets. Nature 2019, 571, 335–342. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop/Stage | Crop/Stage Duration (Days) | Rate (g C m−2 d−1) | Cumulative (g C m−2) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Wheat | Rice | Wheat | Rice | Wheat | Rice | Wheat | Rice | Wheat | Rice | Wheat | Rice | Wheat | Rice | |
| NEE | RE | GPP | NEE | RE | GPP | |||||||||
| Pre-sowing/Pre-transplanting | 4 | 4 | 0.69 | 0.88 | 0.7 | 0.89 | 0.01 | 0.01 | 2.76 | 3.52 | 2.8 | 3.56 | 0.04 | 0.04 |
| Sowing-germination/Post-Transplanting | 9 | 6 | −0.27 | −0.21 | 0.12 | 0.11 | 0.39 | 0.32 | −2.43 | −1.26 | 1.08 | 0.66 | 3.51 | 1.92 |
| Early Vegetative Stage | 11 | 14 | −1.28 | −0.31 | 0.79 | 0.42 | 2.07 | 0.73 | −14.08 | −4.34 | 8.69 | 5.88 | 22.77 | 10.22 |
| Tillering | 27 | 11 | −3.6 | −2.25 | 2.01 | 0.99 | 5.61 | 3.24 | −97.2 | −24.75 | 54.27 | 10.89 | 151.47 | 35.64 |
| Stem Elongation | 28 | 19 | −6.63 | −3.41 | 3.21 | 3.53 | 9.84 | 6.94 | −185.64 | −64.79 | 89.88 | 67.07 | 275.52 | 131.86 |
| Booting | 10 | 13 | −8.1 | −6.54 | 3.77 | 6.71 | 11.87 | 13.25 | −64.8 | −85.02 | 30.16 | 87.23 | 94.96 | 172.25 |
| Heading | 15 | 18 | −10.43 | −7.12 | 5.23 | 7.3 | 15.66 | 14.42 | −125.16 | −128.16 | 62.76 | 131.44 | 187.92 | 259.56 |
| Ripening | 15 | 17 | −4.34 | −3.4 | 2.85 | 4.92 | 7.18 | 8.32 | −86.8 | −57.8 | 57.05 | 83.64 | 143.6 | 141.44 |
| Harvest | 2 | 2 | 0.46 | 0.35 | 1.04 | 0.86 | 0.58 | 0.51 | 0.92 | 0.7 | 2.08 | 1.72 | 1.16 | 1.02 |
| Average/Total (Crop period) | 115 | 98 | −5.01 | −3.74 | 2.64 | 3.22 | 7.65 | 7.68 | −576 | −368 | 304 | 387 | 880 | 753 |
| Fallow (after) | 91 | 61 | 0.4 | 0.28 | 0.87 | 1.02 | 0.51 | 0.74 | 32.76 | 17.08 | 79.17 | 62.22 | 46.41 | 45.14 |
| Environmental Variables | Ecosystem Respiration (RE) | Environmental Variables | Gross Primary Productivity (GPP) | ||
|---|---|---|---|---|---|
| Rice (N = 4712) | Wheat (N = 5472) | Rice (N = 4712) | Wheat (N = 5472) | ||
| AT | 0.060 ns | 0.294 ** | AT | 0.129 * | 0.443 ** |
| ST | 0.543 ** | 0.010 ns | PAR | 0.225 ** | 0.348 ** |
| SM | −0.088 ns | −0.322 ** | VPD | −0.315 ** | −0.147 ns |
| SHF | 0.084 ns | 0.111 ns | NR | 0.248 * | 0.099 ns |
| RH | 0.140 ns | −0.121 ns | SM | 0.172 ns | 0.471 ** |
| GPP | 0.917 ** | 0.698 ** | |||
| Growth Stage | Rice | Wheat | ||||
|---|---|---|---|---|---|---|
| α (mg CO2 µmolephoton−1) | Pmax (mg CO2 m−2s−1) | R2 | α (mg CO2 µmole photon−1) | Pmax (mg CO2 m−2s−1) | R2 | |
| EVS | 0.0007 | 0.347 | 0.67 | 0.0008 | 0.461 | 0.61 |
| LVS/Tillering | 0.0012 | 0.642 | 0.84 | 0.0014 | 0.707 | 0.69 |
| SE/MT-PI | 0.0016 | 1.334 | 0.70 | 0.0017 | 1.469 | 0.73 |
| Booting | 0.0022 | 2.412 | 0.82 | 0.0022 | 2.203 | 0.89 |
| Heading | 0.0023 | 1.909 | 0.91 | 0.0026 | 2.841 | 0.70 |
| Ripening | 0.0018 | 0.938 | 0.76 | 0.0020 | 1.318 | 0.88 |
| Location | Climate | NEE | GPP | RE | RE/GPP | Reference | |
|---|---|---|---|---|---|---|---|
| Rice | International Rice Research Institute, Philippines | Tropical | −258 | −778 | 521 | 0.67 | [34] |
| Mase paddy site, Japan | Tropical | −396 | −1140 | 743 | 0.65 | [24] | |
| Central Rice Research Institute, Cuttak, India | Sub-tropical monsoon | −448 | −811 | 363 | 0.44 | [19] | |
| North China Plain, China | Subtropical semi-humid monsoon | −583 −512 −451 | −1220 −1135 −859 | 637 623 459 | 0.52 0.54 0.53 | [12] | |
| Indian Agricultural Research Institute, New Delhi, India | Sub-tropical, semi-arid | −368 | −753 | 387 | 0.51 | Present study | |
| Wheat | Selhausen test site, Germany (First Year) | Temperate | −627 | −1304 | 676 | 0.51 | [43] |
| Selhausen test site, Germany (Second Year) | Temperate | −537 | −1067 | 529 | 0.49 | [43] | |
| North China Plain, China | Subtropical semi-humid monsoon climate | −438 −431 | −987 −966 | 552 538 | 0.56 0.56 | [12] | |
| Indian Agricultural Research Institute, New Delhi, India | Subtropical semi-arid | −576 | −888 | 304 | 0.34 | Present study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, A.; Bhatia, A.; Sehgal, V.K.; Tomer, R.; Jain, N.; Pathak, H. Net Ecosystem Exchange of Carbon Dioxide in Rice-Spring Wheat System of Northwestern Indo-Gangetic Plains. Land 2021, 10, 701. https://doi.org/10.3390/land10070701
Kumar A, Bhatia A, Sehgal VK, Tomer R, Jain N, Pathak H. Net Ecosystem Exchange of Carbon Dioxide in Rice-Spring Wheat System of Northwestern Indo-Gangetic Plains. Land. 2021; 10(7):701. https://doi.org/10.3390/land10070701
Chicago/Turabian StyleKumar, Amit, Arti Bhatia, Vinay Kumar Sehgal, Ritu Tomer, Niveta Jain, and Himanshu Pathak. 2021. "Net Ecosystem Exchange of Carbon Dioxide in Rice-Spring Wheat System of Northwestern Indo-Gangetic Plains" Land 10, no. 7: 701. https://doi.org/10.3390/land10070701