

Reconstruction of the Late Holocene Environment of Central Yakutia Based on Fossil Invertebrates and Plants from a Buried Lake at the Vilyuy River Valley

 , ,
, ,
Abstract
:1. Introduction
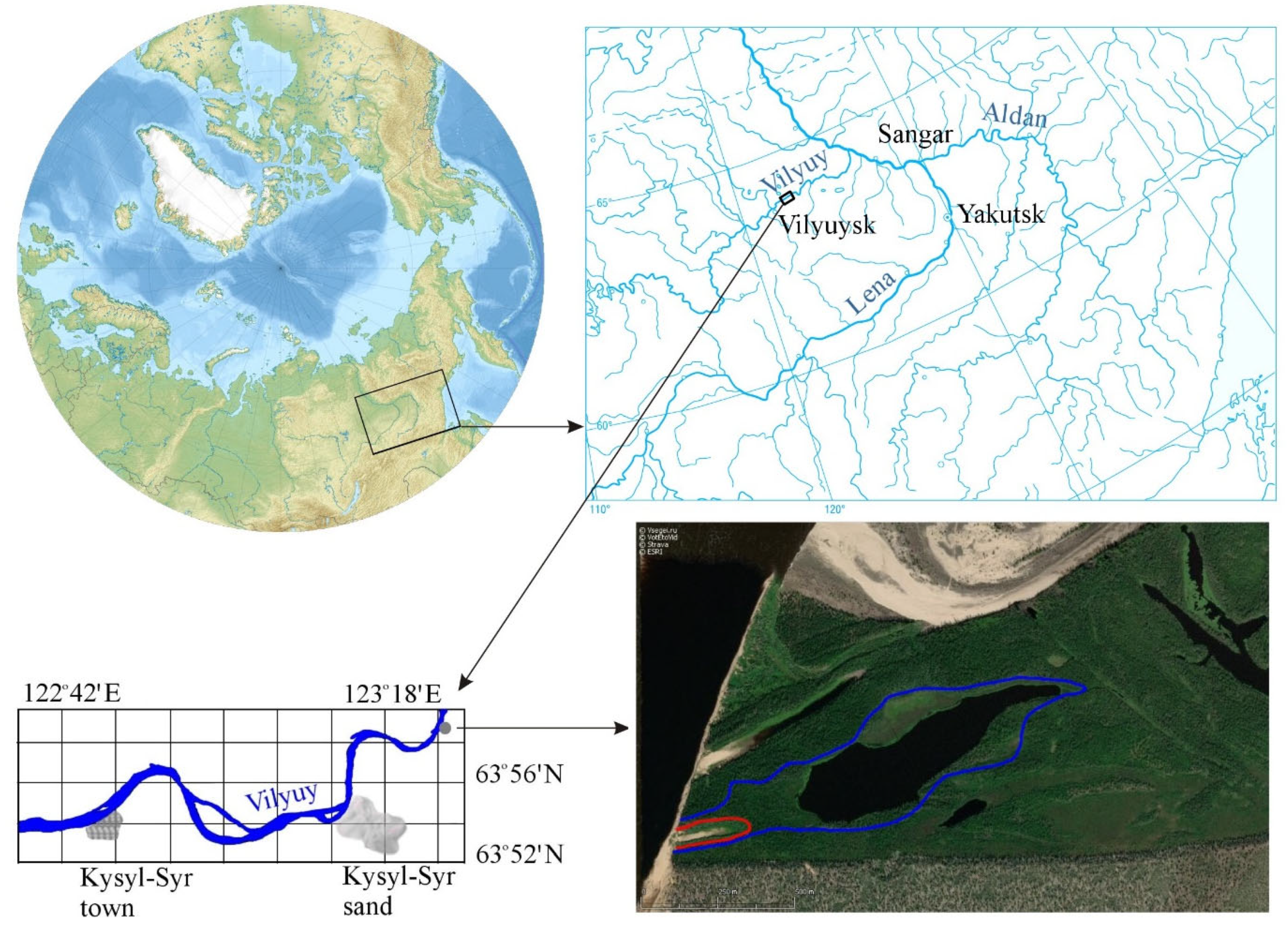
2. Regional Setting
3. Materials and Methods
4. Results
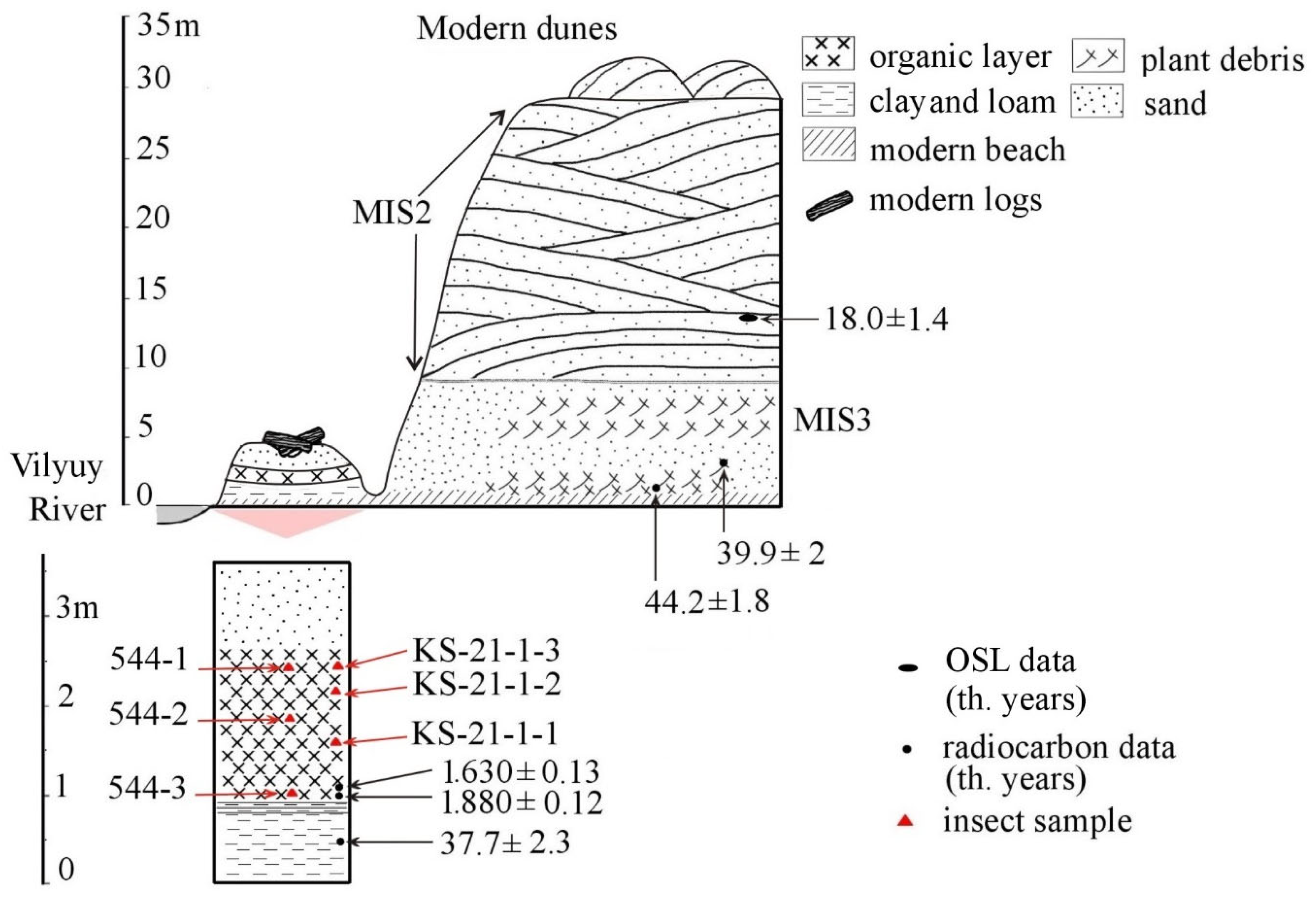
4.1. Section Stratigraphy
- Layer at 0–75 cm: bluish-grey gleied loam with reddish ferruginous spots and plant inclusions, solid, bedding is unclear.
- Layer at 75–85 cm: laminated loam deposit with alternating light and dark laminae, several mm thick.
- Layer at 85–90 cm: brown loosely packed organics with admixture of sand and silt.
- Layer at 90–160 cm: dark brown and reddish-brown consolidated gyttja-like organic material with fragments of grasses, wood, and spruce cones.
- Layer at 160–360 cm: modern alluvial sand with logs in the central part of the section.
4.2. Macrofossils
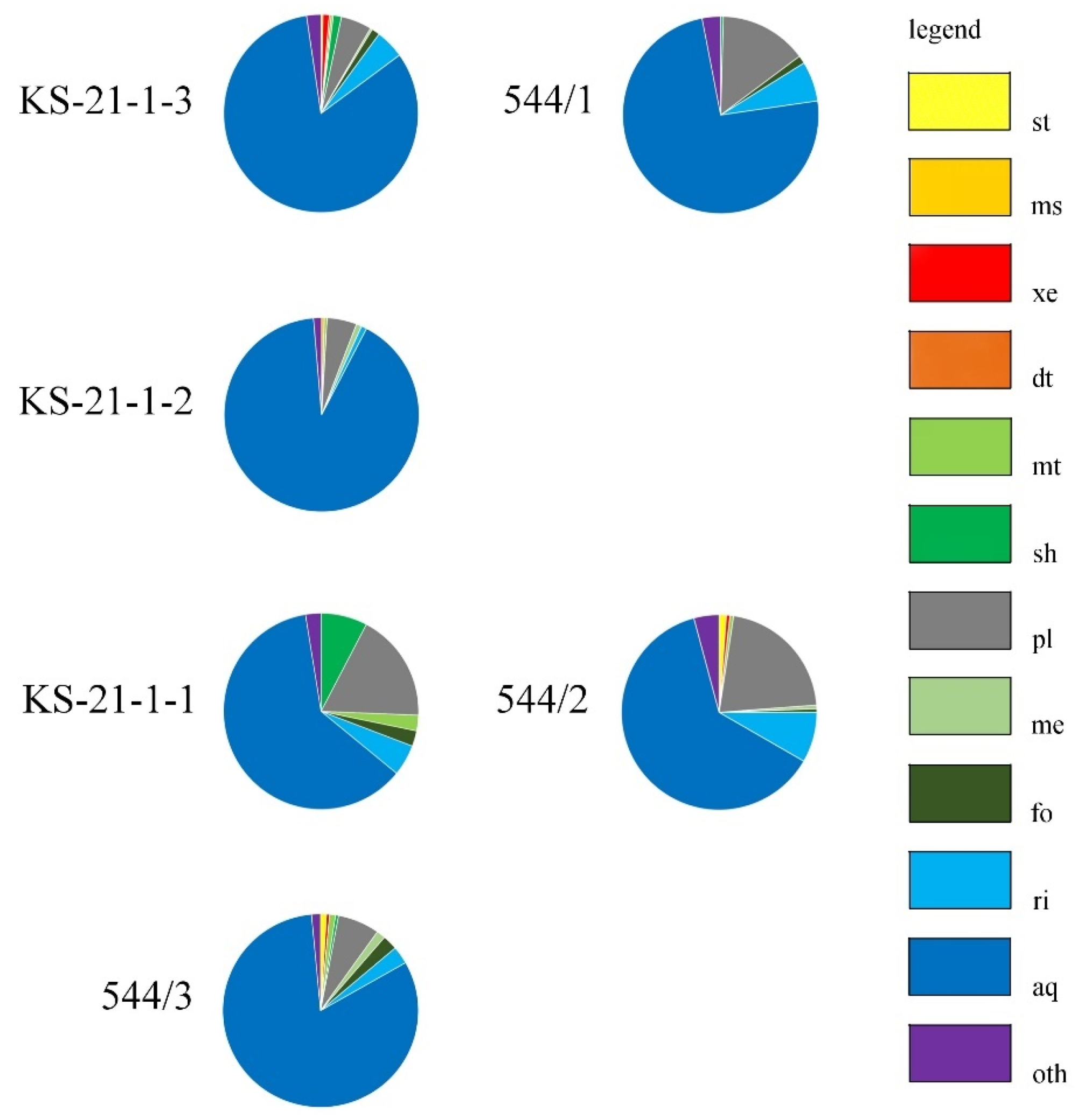
4.2.1. Insects and Other Invertebrates
4.2.2. Plants
4.2.3. Charred Plant Remains
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clements, F.E. Plant Succession: An Analysis of the Development of Vegetation; Carnegie Institution of Washington: Washington, DC, USA, 1916. [Google Scholar]
- Horne, A.J.; Goldman, C.R. Limnology, 2nd ed.; McGraw-Hill: New York, NY, USA, 1994. [Google Scholar]
- Odum, E.P.; Barrett, G.W. Fundamentals of Ecology, 5th ed.; Thomson Brooks/Cole: Belmont, CA, USA, 2005. [Google Scholar]
- Wetzel, R.G. Limnology, Lake And River Ecosystem, 3rd ed.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Rydin, H.; Jeglum, J.K. The Biology of Peatlands; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Speller, J.; Forbes, V. On the role of peat bogs as components of Indigenous cultural landscapes in Northern North America. Arct. Antarct. Alp. Res. 2022, 54, 96–110. [Google Scholar] [CrossRef]
- Cohen, A.S. Paleolimnology: The History and Evolution of Lake Systems; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Douglas, M.S.V. Paleolimnology. Overview of Paleolimnology. In Encyclopedia of Quaternary Science, 2nd ed.; Elias, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 259–270. [Google Scholar]
- Telka, A.M.; Brooks, G.R. Macrofossil Reports from Core MUS1, Muskrat Bay, lower French River Area, and Core DPN2, Depensier Lake, North Bay, Ontario; Open File 5901; Geological Survey of Canada: Ottawa, ON, Canada, 2008; 26p. [Google Scholar]
- Elias, S.A. Quaternary Insects and Their Environments; Smithsonian Institution Press: Washington, DC, USA, 1994. [Google Scholar]
- Elias, S. Coleoptera and Trichoptera. In Tracking Environmental Change Using Lake Sediments: Biological and Statistical Techniques; Smol, J.P., Last, W.M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 67–80. [Google Scholar]
- Nazarov, V.I. Reconstruction of the Belarussian Landscapes by Palaeoentomological Data. (Anthropogen); Trudy of the Palaeontological Institute: Moscow, Nauka, 1984; Volume 205, 96p. (In Russian) [Google Scholar]
- Lavoie, C.; Elias, S.A.; Payette, S. Holocene fossil beetles from a treeline peatland in subarctic Quebec. Can. J. Zool. 1997, 75, 227–236. [Google Scholar] [CrossRef]
- Olsson, F.; Lemdahl, G. A continuous Holocene beetle record from the site Stavsakra, southern Sweden: Implications for the last 10600 years of forest and land use history. J. Quat. Sci. 2009, 24, 612–626. [Google Scholar] [CrossRef]
- Whitehouse, N.J.; Langdon, P.G.; Bustin, R.; Galsworthy, S. Fossil insects and ecosystem dynamics in wetlands: Implications for biodiversity and conservation. Biodivers. Conserv. 2008, 17, 2055–2078. [Google Scholar] [CrossRef]
- Kiselev, S.V.; Nazarov, V.I. Late Cenozoic Insects of Northern Eurasia. Pleiades Publishing, Ltd.: New York, NY, USA. Paleontol. J. Suppl. 2009, 43, 723–850. [Google Scholar] [CrossRef]
- Zhang, T.; Elias, S.A. Holocene palaeoenvironmental reconstruction based on fossil beetle faunas from the Southern Altai region, north-west China. J. Quat. Sci. 2019, 34, 593–602. [Google Scholar] [CrossRef]
- Kiselev, S.V. Late Cenozoic Coleoptera of North-East Siberia; Nauka: Moscow, Russia, 1981. (In Russian) [Google Scholar]
- Berman, D.I.; Alfimov, A.V.; Mazhitova, G.G.; Grishkan, I.B.; Yurtsev, B.A. Cold Steppe in North-Eastern Asia; Dal’nauka: Vladivostok, Russia, 2001; 183p. (In Russian) [Google Scholar]
- Berman, D.; Alfimov, A.; Kuzmina, S. Invertebrates of the relict steppe ecosystems of Beringia, and the reconstruction of Pleistocene landscapes. Quat. Sci. Rev. 2011, 30, 2200–2219. [Google Scholar] [CrossRef]
- Ashastina, K.; Kuzmina, S.; Rudaya, N.; Troeva, E.; Schoch, W.H.; Romermann, C.; Reinecke, J.; Otte, V.; Savvinov, G.; Wesche, K.; et al. Woodlands and steppes: Pleistocene vegetation in Yakutia’s most continental part recorded in the Batagay permafrost sequence. Quat. Sci. Rev. 2018, 196, 38–61. [Google Scholar] [CrossRef]
- Kuzmina, S.A. Quaternary Insects and Environment of the Northeastern Asia. Pleiades Publishing, Ltd.: New York, NY, USA. Paleontol. J. Suppl. 2015, 49, 679–867. [Google Scholar] [CrossRef]
- Smirnova, N.S. (Ed.) Scientific-Applied Reference Book on Climate of the USSR; Yakutskaya ASSR; Gidrometeoizdat: Leningrad, Russia, 1989; Issue 24; 608p. (In Russian) [Google Scholar]
- Karavaev, M.N. Synopsis of the Flora of Yakutia; Academy of Sciences of the USSR Publisher: Moscow-Leningrad, Russia, 1958; 189p. (In Russian) [Google Scholar]
- Zakharova, V.I.; Cherosov, M.M.; Troeva, E.I.; Gogoleva, P.A. Steppes. In The Far North: Plant Biodiversity and Ecology of Yakutia; Elena, I., Troeva, A.P., Isaev, M.M., Karpov, C.N.S., Eds.; Series Plant and Vegetation; Springer Science & Business Media: Dordrecht, The Netherlands, 2010; Volume 3, pp. 193–198. [Google Scholar]
- Galanin, A.A. Late Quaternary sand covers of Central Yakutia (Eastern Siberia): Structure, facies composition and paleoenvironment significance. Krioshera Zemli 2021, 25, 3–34. (In Russian) [Google Scholar] [CrossRef]
- Galanin, A.A.; Pavlova, M.R. Late Quaternary dune formations (D’olkuminskaya Series) in Central Yakutia (Part 2). Krioshera Zemli 2019, 23, 3–16. (In Russian) [Google Scholar] [CrossRef]
- Galanin, A.A.; Pavlova, M.R.; Shaposhnikov, G.I.; Lytkin, V.M. Tukulans: Sand deserts of Yakutia. Priroda 2016, 11, 44–55. (In Russian) [Google Scholar]
- Reimer, P.J.; Austin, W.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Ramsey, C.B.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Bigelow, N.H.; Zazula, G.D.; Atkinson, D.E. Plant macrofossil records. Arctic North America. In Encyclopedia of Quaternary Science; Elias, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 2434–2450. [Google Scholar]
- Kienast, F. Plant macrofossil records. Arctic Eurasia. In The Encyclopedia of Quaternary Science; Elias, S.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 3, pp. 733–745. [Google Scholar]
- Kienast, F.; Wetterich, S.; Kuzmina, S.; Schirrmeister, L.; Andreev, A.; Tarasov, P.; Nazarova, L.; Kossler, A.; Frolova, L.; Kunitsky, V. Paleontological records indicate the occurrence of open woodlands in a dry inland climate at the present-day Arctic coast in western Beringia during the Last Interglacial. Quat. Sci. Rev. 2011, 30, 2134–2159. [Google Scholar] [CrossRef] [Green Version]
- Sher, A.V.; Kuzmina, S. Beetle records/Late Pleistocene of Northern Asia. In Encyclopedia of Quaternary Science; Elias, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 1, pp. 246–267. [Google Scholar]
- Kuzmina, S.A. Macroentomology Analysis: Methods, Opportunities, and Examples of Reconstructions of Paleoclimatic and Paleoenvironmental Conditions in the Quaternary of the Northeastern Siberia. Contemp. Probl. Ecol. 2017, 10, 336–349. [Google Scholar] [CrossRef]
- Carr, R. Limnebius crinifer Rey new to Britain, with a revised key to the British Limnebius species (Coleoptera: Hydraenidae). Entomol. Gaz. 1984, 35, 99–102. [Google Scholar]
- Mansell, L.J.; Whitehouse, N.J.; Gearey, B.R.; Barratt, P.; Roe, H.M. Holocene floodplain palaeoecology of the Humberhead Levels; implications for regional wetland development. Quat. Int. 2014, 341, 91–109. [Google Scholar] [CrossRef]
- Sprick, P. Suitability of an insect group for the habitat directive of the EU: The weevil subfamily Bagoinae (Col., Curculionidae). Contributions to the ecology of phytophagous beetles VII. Snudebiller 2001, 2, 7–40. [Google Scholar]
- Lukin, E.I. Leeches of Fresh and Brackish Water Bodies (Fauna of Leeches of the USSR); Nauka: Leningrad, Russia, 1976; 284p. (In Russian) [Google Scholar]
- Hann, B.J. Cladocera. In Methods in Quaternary Ecology; Warner, B.G., Ed.; Geoscience Canada Reprint Ser. 5; Geological Association of Canada: Ottawa, ON, Canada, 1990; pp. 81–91. [Google Scholar]
- Kotov, A.A.; Kuzmina, S.A.; Frolova, L.A.; Zharov, A.A.; Neretina, A.N.; Smirnov, N.N. Ephippia of the Daphniidae (Branchiopoda: Cladocera) in Late Caenozoic deposits: Untapped source of information for palaeoenvironment reconstructions in the Northern Holarctic. Invertebr. Zool. 2019, 16, 183–199. [Google Scholar] [CrossRef]
- Rogers, D.C.; Zharov, A.A.; Neretina, A.N.; Kuzmina, S.A.; Kotov, A.A. A Review of Recently Discovered Remains of the Pleistocene Branchiopods (Anostraca, Notostraca) from NE Siberia and Arctic Canada. Water 2021, 13, 183–199. [Google Scholar] [CrossRef]
- Tremel, B.; Frey, S.E.; Yan, N.D.; Somers, K.M.; Pawson, T.W. Habitat specificity of littoral Chydoridae (Crustacea, Branchiopoda, Anomopoda) in Plastic Lake, Ontario, Canada. Hydrobiologia 2000, 432, 195–205. [Google Scholar] [CrossRef]
- Kuzmina, S.A. A Record of Phylactolaemata from Late Cenozoic Continental Deposits of Northern Eurasia and North America. Paleontol. J. 2022, 56, 852–861. [Google Scholar] [CrossRef]
- Vinogradov, A.V. Phylactolaemata and Bryozoa of Continental Water Bodies of Eurasia; Faunistics, ecology, zoogeography and evolution of the Phylactolaemata and Bryozoa of continental water bodies of Eurasia. General bryozoology of continental water bodies of Eurasial; Lambert Academic Publishing (LAP): Saarbrucken, Germany, 2011; Volumes 1, 2. (In Russian) [Google Scholar]
- Mackie, G.L. Biology of Freshwater Corbiculid and Sphaeriid Clams of North America; Ohio Biological Survey Bulletin New Series; Ohio Biological Survey: Columbus, OH, USA, 2007; Volume 15, 436p. [Google Scholar]
- Sharapova, T.A.; Trylis, V.V.; Ivanov, S.N.; Ilyushina, V.V. Composition and distribution of sponges (Porifera) in continental waters of western Siberia. Contemp. Probl. Ecol. 2014, 7, 543–550. [Google Scholar] [CrossRef]
- Frost, T.M.; De Nagy, G.S.; Gilbert, J.J. Population Dynamics and Standing Biomass of the Freshwater Sponge Spongilla lacustris. Ecology 1982, 63, 1203–1210. [Google Scholar] [CrossRef]
- Harrison, F.W. Freshwater Sponges. In Methods in Quaternary Ecology; Warner, B.G., Ed.; Geoscience Canada Reprint, Ser. 5; Geological Association of Canada: Ottawa, ON, Canada, 1990; pp. 75–80. [Google Scholar]
- Harrison, F.W.; Warner, B.G. Fossil freshwater sponges (Porifera: Spongillidae) from Western Canada: An overlooked group of Quaternary paleoecological indicators. Trans. Am. Microsc. Soc. 1986, 105, 110–120. [Google Scholar] [CrossRef]
- Sazhnev, A.S.; Ivicheva, K.N.; Komarova, A.S.; Philippov, D.A. A review of aquatic, semi-aquatic and amphibiotic beetles (Insecta: Coleoptera) of Vologodskaya Oblast, Russia. Eurasian Entomolog. J. 2019, 18, 60–74. (In Russian) [Google Scholar] [CrossRef] [Green Version]
- Sazhnev, A.S.; Philippov, D.A. About aquatic and amphibiotic beetles (Insecta: Coleoptera) of boggy water of Vologodskaya Oblast. Rep. Inst. Biol. Inn. Water RAS 2017, 79, 194–199. [Google Scholar] [CrossRef]
- Bienkowski, A.O. Aquatic Leaf Beetles (Coleoptera: Chrysomelidae: Donaciinae); Publisher Mukhamedov G.V.: Livny, Russia, 2014; 380p. (In Russian) [Google Scholar]
- Anderson, R.S. Weevils (Coleoptera: Curculionoidea, Excluding Scolytinae and Platypodinae) of the Yukon. In Insects of the Yukon; Danks, H.V., Downes, J.A., Eds.; Biological Survey of Canada (Terrestrial Arthropods): Ottawa, ON, Canada, 1997; pp. 523–562. [Google Scholar]
- Volovnik, S.V. On the distribution and ecology of some Lixine species (Coleoptera, Curculionidae). V. genus Lixus subgenera Lixus, F.; Ortholixus Reitt., Compsolixus Reitt., Callistrolixus Reitt. Entomol. Rev. 2012, 93, 583–590. (In Russian) [Google Scholar] [CrossRef]
- Meshcheryakova, E.N.; Berman, D.I. Cold hardiness and geographic distribution of earthworms (Oligochaeta, Lumbricidae, Moniligastridae). Entomolog. Rev. 2014, 94, 486–497. [Google Scholar] [CrossRef]
- Ubugunova, V.I.; Boeskorov, V.S.; Savinov, G.N. Distribution of earthworms in soils of Yakutia. Eurasian Soil Sci. 2007, 40, 59–63. [Google Scholar] [CrossRef]
- Ashburner, K.; McAllister, H.A. The Genus Betula: A Taxonomic Revision of Birches (A Botanical Magazine Monograph); Royal Botanic Gardens: Kew, UK, 2013; 431p. [Google Scholar]
- Sokolov, S.Y. (Ed.) Trees and Shrubs of the USSR. Vol. 2 Angiospermous; Komarov Botanical Institute AS USSR: Moscow-Leningrad, Russia, 1951; 612p. (In Russian) [Google Scholar]
- Danilova, N.S. (Ed.) Red Book of the Republic Sakha (Yakutia). Vol. 1. Rare and Endangered Species of Plants and Fungi; Reart: Moscow, Russia, 2017; 412p. (In Russian) [Google Scholar]
- Danilova, N.S.; Ivanova, N.S.; Borisova, S.Z. Ecological and biological features and structure of Dracocephalum ruyschiana biocenosis populations in Central Yakutia. Sci. Educ. 2012, 3, 33–38. (In Russian) [Google Scholar]
- Dulepova, N.A.; Korolyuk, A.Y. Obligate fraction of psammophyte flora of Siberia. Flora Asiat. Russ. 2012, 2, 101–107. (In Russian) [Google Scholar]
- PonPonomarenko, E.; Tomson, P.; Ershova, E.; Bakumenko, V. A multi-proxy analysis of sandy soils in historical slash-and-burn sites: Karula case study. Quat. Int. 2018, 516, 190–206. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Tree Rings and Environment-Dendrochronology; Haupt: Bern, Switzerland, 1996. [Google Scholar]
- Ponomarenko, E.V.; Ershova, E.G.; Stashenkov, D.A.; Ponomarenko, D.S.; Kochkina, A.F. Tracing land use history using a combination of soil charcoal and soil pollen analysis: An example from colluvial deposits of the Middle Volga Region. J. Archaeol. Sci. Rep. 2020, 31, 102269. [Google Scholar] [CrossRef]
- Payette, S. Late-Holocene development of subarctic ombrotrophic peatlands: Allogenic and autogenic succession. Ecology 1988, 69, 516–531. [Google Scholar] [CrossRef]
- Beaulieu-Audy, V.; Garneau, M.; Richard, P.J.H.; Asnong, H. Holocene palaeoecological reconstruction of three boreal peatlands in the La Grande Rivière region, Québec, Canada. Holocene 2009, 19, 459–476. [Google Scholar] [CrossRef]
- Ryberg, E.E.; Valiranta, M.; Martinez-Cortizas, A.; Ehrlen, J.; Sjostrom, J.K.; Kylander, M.E. Postglacial peatland vegetation succession in Store Mosse bog, south-central Sweden: An exploration of factors driving species change. Boreas 2022, 51, 651–666. [Google Scholar] [CrossRef]
- Reyes, A.V.; Zazula, G.D.; Kuzina, S.; Ager, T.A.; Froese, D.G. Identification of last interglacial deposits in eastern Beringia: A cautionary note from the Palisades, interior Alaska. J. Quat. Sci. 2011, 26, 345–352. [Google Scholar] [CrossRef]
- Kuznetsova, T.V.; Kuzmina, S.A.; Kunitsky, V.V.; Schirrmeister, L.; Sher, A.V. The fauna of alas sequences in the Ice Complex area: The case of Mamontovy Bysagasa northwest exposure, Bykovsky Peninsula. In Proceedings of the Fifth Workshop on Russian-German Cooperation: Laptev Sea System 2000, Saint Petersburg, Russia, 26–28 November 1999; Terra Nostra. Schriften der Alfred-Wegener-Stiftung: Koln, Germany, 1999; pp. 49–50. [Google Scholar]
- Head, M.E. Formal subdivision of the Quaternary System/Period: Present status and future directions. Quat. Int. 2019, 500, 32–51. [Google Scholar] [CrossRef]
- Walker, M.; Head, M.J.; Berkelhammer, M.; Björck, S.; Cheng, H.; Cwynar, L.; Fisher, D.; Gkinis, V.; Long, A.; Lowe, J.; et al. Formal ratification of the subdivision of the Holocene Series/Epoch (Quaternary System/Period): Two new Global Boundary Stratotype Sections and Points (GSSPs) and three new stages/subseries. Episodes 2018, 41, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Sernander, R. On the evidence of post-glacial changes of climate furnished by the peat mosses of northern Europe. Geol. Föreningens I Stockh. Förhaldlinger 1908, 30, 365–478. [Google Scholar] [CrossRef]
- McCormick, M.; Büntgen, U.; Cane, M.; Cook, E.R.; Harper, K.; Huybers, P.; Litt, T.; Manning, S.W.; Mayewski, P.A.; More, A.F.; et al. Climate change during and after the Roman Empire: Reconstructing the past from scientific and historical evidence. J. Interdiscip. Hist. 2012, 43, 169–220. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Surge, D.; Mithen, S. Seasonal temperature variability of the Neoglacial (3300–2500 BP) and Roman Warm Period (2500–1600 BP) reconstructed from oxygen isotope ratios of limpet shells (Patella vulgata), Northwest Scotland. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 317–318, 104–113. [Google Scholar] [CrossRef]
- Nazarova, L.; Lüpfert, H.; Subetto, D.; Pestryakova, L.; Diekmann, B. Holocene climate conditions in central Yakutia (Eastern Siberia) inferred from sediment composition and fossil chironomids of Lake Temje. Quat. Int. 2013, 290–291, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Pestryakova, L.; Herzschuh, U.; Wetterich, S.; Ulrich, M. Present-day variability and Holocene dynamics of permafrost-affected lakes in central Yakutia (Eastern Siberia) inferred from diatom records. Quat. Sci. Rev. 2012, 51, 56–70. [Google Scholar] [CrossRef]
- Velichko, A.A.; Andreev, A.A.; Klimanov, V.A. Climate and vegetation dynamics in the tundra and forest zone during the Late Glacial and Holocene. Quat. Int. 1997, 41/42, 71–96. [Google Scholar] [CrossRef]
- Andreev, A.; Tarasov, P. Northern Asia. In The Encyclopedia of Quaternary Science, 2nd ed.; Elias, S.A., Mock, C.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 4, pp. 164–172. [Google Scholar] [CrossRef]
- Glückler, R.; Geng, R.; Grimm, L.; Baisheva, I.; Herzschuh, U.; Stoof-Leichsenring, K.R.; Kruse, S.; Andreev, A.; Pestryakova, L.; Dietze, E. Holocene wildfire and vegetation dynamics in Central Yakutia, Siberia, reconstructed from lake-sediment proxies. EGUsphere 2022. preprint. [Google Scholar] [CrossRef]
- Andreev, A.A.; Klimanov, V.A.; Sulerzhitsky, L.D. Vegetation and climate history of central Yakutia during the Holocene and Late Pleistocene. Bot. Zhurnal 2002, 87, 86–98. (In Russian) [Google Scholar]
- Pavlova, M.R.; Rudaya, N.A.; Galanin, A.A.; Shaposhnikov, G.I. The structure and evolution of dune massifs in the Vilyuy River basin in the late Quaternary (a case study of Makhatta and Kysyl-Syr tukulans). Contemp. Probl. Ecol. 2017, 24, 473–486. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lab. No | Depth (m) | Radiocarbon Ages BP | Calibrated Age BP IntCal20 | Material | Reference |
|---|---|---|---|---|---|
| MPI-156 | 3.1 | 37,745 ± 2300 | 43,190 ± 2810 | shrub remains | Figure 3 in this study |
| MPI-227 | 2.75 | 1860 ± 120 | 1780 ± 150 | wood | Figure 3 in this study |
| MPI-162 | 2.7 | 1630 ± 130 | 1530 ± 130 | shrub remains | Figure 3 in this study |
| MPI-36 | 30 | 39,900 ± 2000 | 44,540 ± 2240 | plant debris | Figure 3 [27] |
| MPI-38 | 33 | 44,200 ± 1800 | 47,730 ± 2280 | plant debris | Figure 3 [27] |
| MPI-43 | 0.1–0.2 | 3320 ± 90 | 3560 ± 110 | moss, sedge | Figure 9 [27] |
| MPI-48 | 1.2–1.3 | 3990 ± 90 | 4470 ± 150 | peat | Figure 9 [27] |
| MPI-52 | 1.3–1.4 | 5840 ± 100 | 6650 ± 120 | peat with wood | Figure 9 [27] |
| MPI-57 | 1.6–1.7 | 8250 ± 100 | 9230 ± 130 | gyttja | Figure 9 [27] |
| OSL | |||||
| RLQG 2265-035 | 23 | 18,000 ± 1400 | quartz sand | Figure 3 [27] | |
| Taxa | Eco | 544/3 | KS-21-1-1 | KS-21-1-2 | 544/2 | KS-21-1-3 | 544/1 |
|---|---|---|---|---|---|---|---|
| lower | middle | top | |||||
| Arthropoda, Insecta, Coleoptera | |||||||
| Gyrinidae | |||||||
| Gyrinus natator L. | aq | 2 | 0 | 5 | 0 | 3 | 1 |
| G. opacus Sahlb. | aq | 1 | 0 | 0 | 0 | 0 | 0 |
| Gyrinus sp. | aq | 0 | 2 | 0 | 1 | 0 | 0 |
| Carabidae | |||||||
| Miscodera arctica (Payk) | me | 1 | 0 | 0 | 0 | 0 | 0 |
| Elaphrus splendidus F.-W. | ri | 0 | 0 | 0 | 0 | 0 | 1 |
| Bembidion (Notaphus) sp. | ri | 1 | 0 | 0 | 0 | 0 | 0 |
| B. (Peryphus) sp. | ri | 0 | 0 | 0 | 1 | 1 | 0 |
| B. (Diplocampa) transparens (Gebl.) | ri | 1 | 0 | 0 | 0 | 0 | 0 |
| B. (Trepanedoris) doris Pan. | ri | 0 | 0 | 0 | 0 | 0 | 1 |
| Bembidion sp. | ri | 0 | 0 | 0 | 1 | 1 | 0 |
| Dicheirotrichus mannerheimi Sahlb. | xe | 1 | 0 | 0 | 1 | 0 | 0 |
| Cymindis sp. | xe | 1 | 0 | 0 | 0 | 1 | 0 |
| Agonum (Europhilus) consimile Gyll. | ri | 0 | 0 | 0 | 4 | 0 | 0 |
| A. (Europhilus) sexpunctatum L. | ri | 1 | 0 | 0 | 0 | 0 | 0 |
| A. (Olisares) duftschmidi J.Sch. | ri | 0 | 0 | 0 | 0 | 0 | 3 |
| A. (Agonothorax) exaratum Schalb. | ri | 0 | 0 | 0 | 0 | 0 | 2 |
| Agonum sp. | ri | 0 | 1 | 0 | 0 | 4 | 0 |
| Poecilus (Derus) nearcticus Lth. | dt | 0 | 0 | 0 | 0 | 1 | 0 |
| Pterostichus (Cryobius) brevicornis (Kby.) | mt | 0 | 0 | 0 | 0 | 0 | 3 |
| P. (Cryobius) ventricosus Esch. | mt | 0 | 0 | 0 | 0 | 1 | 0 |
| Pterostichus (Cryobius) spp. | mt | 2 | 0 | 3 | 1 | 0 | 0 |
| Curtonotus torridus Pz. | xe | 0 | 0 | 0 | 0 | 1 | 0 |
| Carabidae gen. indet. | oth | 0 | 0 | 0 | 0 | 1 | 0 |
| Dytiscidae | |||||||
| Agabus sp. | aq | 0 | 0 | 0 | 1 | 1 | 2 |
| Colymbetes sp. | aq | 1 | 1 | 1 | 1 | 1 | 2 |
| Dytiscidae gen. indet. | aq | 1 | 0 | 0 | 1 | 3 | 1 |
| Hydrophilidae | |||||||
| Cymbiodyta marginella (F.) | ri | 0 | 0 | 0 | 0 | 1 | 1 |
| Cercyon sp. | pl | 0 | 1 | 0 | 0 | 0 | 0 |
| Hydraenidae | |||||||
| Limnebius sp. | aq | 1 | 0 | 0 | 0 | 0 | 0 |
| Staphylinidae | |||||||
| Eucnecosum tenue (LeC.) | pl | 2 | 2 | 0 | 1 | 0 | 0 |
| Olophrum latum Maekl. | pl | 0 | 0 | 0 | 0 | 1 | 0 |
| O. rotundicolle Sahlb. | pl | 1 | 0 | 0 | 5 | 1 | 7 |
| Olophrum sp. | pl | 0 | 0 | 1 | 0 | 0 | 0 |
| Tachyporus sp. | pl | 0 | 0 | 0 | 1 | 0 | 0 |
| Stenus sp. | ri | 0 | 0 | 0 | 1 | 0 | 1 |
| Lathrobium brunnipes (F.) | pl | 0 | 0 | 0 | 0 | 1 | 0 |
| L. longulum Grav. | pl | 0 | 0 | 1 | 0 | 0 | 0 |
| Lathrobium sp. | pl | 1 | 0 | 0 | 0 | 0 | 0 |
| Gymmusa sp. | pl | 1 | 0 | 0 | 0 | 0 | 0 |
| Aleocharinae gen. indet | pl | 0 | 0 | 1 | 2 | 0 | 0 |
| Philonthus sp. | pl | 0 | 0 | 0 | 1 | 0 | 0 |
| Staphylinidae gen. indet. | pl | 0 | 0 | 0 | 0 | 0 | 2 |
| Scirtidae | |||||||
| Cyphon variabilis Thunb | ri | 0 | 0 | 0 | 1 | 1 | 1 |
| Byrrhidae | |||||||
| Cytilus sericeus Forst. | me | 0 | 0 | 1 | 0 | 1 | 0 |
| Porcinolus murinus (F.) | st | 1 | 0 | 0 | 2 | 0 | 0 |
| Morychus sp. | xe | 0 | 0 | 0 | 0 | 1 | 0 |
| Simplocaria semistriata F. | me | 1 | 0 | 0 | 0 | 0 | 0 |
| Heteroceridae | |||||||
| Heterocerus fenestratus (Thunb.) | ri | 1 | 0 | 0 | 1 | 0 | 0 |
| Elateridae | |||||||
| Oedostethus femoralis LeC. | ri | 0 | 0 | 0 | 0 | 2 | 0 |
| Elateridae gen. indet. | ri? | 1 | 0 | 0 | 0 | 0 | 0 |
| Bostrichidae | |||||||
| Stephanopachys substriatus Payk. | fo | 0 | 0 | 0 | 0 | 1 | 0 |
| Anthicidae | |||||||
| Anthicus ater Pz. | me | 0 | 0 | 0 | 1 | 0 | 0 |
| Chrysomelidae | |||||||
| Donacia splendens Jac. | ri | 0 | 0 | 0 | 3 | 4 | 6 |
| Donacia sp. | ri | 0 | 1 | 2 | 2 | 0 | 0 |
| Phaedon concinnus Steph. | me | 0 | 0 | 0 | 0 | 0 | 1 |
| Gonioctena affinis Gyll. | sh | 0 | 1 | 0 | 0 | 0 | 0 |
| Aphthona violacea (Koch) | fo | 1 | 0 | 0 | 0 | 0 | 0 |
| Aphthona sp. | fo? | 0 | 0 | 1 | 0 | 0 | 0 |
| Chaetocnema sp. | me | 0 | 1 | 0 | 0 | 0 | 0 |
| Galerucella grisescens Joann. | fo | 0 | 0 | 0 | 0 | 1 | 0 |
| Curculionidae | |||||||
| Tourotaris bimaculata (F.) | ri | 0 | 0 | 0 | 1 | 0 | 0 |
| Bagous limosus Gyll. | aq | 0 | 0 | 0 | 0 | 1 | 1 |
| Ceutorhynchus sp. | xe | 0 | 0 | 0 | 0 | 1 | 0 |
| Phyllobius kolymensis Kor. et Egorov | ms | 0 | 0 | 1 | 0 | 0 | 0 |
| P. virideaeris Laich. | me | 1 | 0 | 0 | 0 | 0 | 0 |
| Hypera viciae (Gyll.) | me | 0 | 0 | 0 | 0 | 1 | 0 |
| Lixus paraplecticus L. | ri | 1 | 0 | 0 | 0 | 0 | 1 |
| Cleonini gen. indet. | ms | 0 | 0 | 0 | 0 | 1 | 0 |
| Lepyrus canadensis Csy. | sh | 0 | 0 | 0 | 0 | 1 | 0 |
| Lepyrus sp. | sh | 0 | 2 | 0 | 0 | 1 | 1 |
| Pissodes sp. | fo | 0 | 0 | 0 | 0 | 1 | 0 |
| Dorytomus sp. | sh | 0 | 0 | 0 | 0 | 1 | 0 |
| Rhynchaenus sp. | sh | 0 | 0 | 0 | 0 | 1 | 0 |
| Tachyerges stigma Germ. | sh | 0 | 0 | 1 | 0 | 0 | 0 |
| Isochnus flagellum Erics. | sh | 1 | 0 | 0 | 0 | 0 | 0 |
| Curculionidae gen. indet. | oth | 0 | 0 | 0 | 2 | 0 | 0 |
| Arthropoda, Insecta, Hymenoptera | |||||||
| Symphyta, Tenthredinidae | |||||||
| Tenthredinidae gen. indet. (cocoon) | oth | 0 | 0 | 1 | 0 | 0 | 0 |
| Aculeata, Formicidae | |||||||
| Formica sp. | fo | 0 | 0 | 0 | 1 | 0 | 1 |
| Formica sp.? | fo | 1 | 0 | 0 | 0 | 0 | 1 |
| Camponotus herculeanus L. | fo | 3 | 1 | 0 | 0 | 2 | 0 |
| Parasitica, Ichneumonidae | |||||||
| Ichneumonidae gen. indet. (cocoon) | oth | 0 | 0 | 0 | 0 | 0 | 1 |
| Parasitica gen. indet. | oth | 3 | 0 | 0 | 0 | 0 | 0 |
| Arthropoda, Insecta, Neuroptera | |||||||
| Chrysopidae | |||||||
| Chrysopidae gen. indet. (cocoon) | oth | 0 | 0 | 2 | 0 | 2 | 3 |
| Arthropoda, Insecta, Diptera | |||||||
| Chironomidae | |||||||
| Chironomidae gen. indet. (larvae) | aq | 0 | 1 | 0 | 1 | 0 | 1 |
| Diptera gen. indet. (puparia) | oth | 0 | 0 | 0 | 5 | 3 | 3 |
| Arthropoda, Insecta, Trichoptera | |||||||
| Brachycentridae | |||||||
| Brachycentrus subnubilus Curtis | aq | 3 | 0 | 0 | 4 | 0 | 7 |
| Micrasema gelidum McLachlan? | aq | 0 | 0 | 1 | 1 | 0 | 0 |
| Arachnida, Oribatida | |||||||
| Oribatida gen. indet. | oth | 0 | 1 | 0 | 0 | 0 | 0 |
| Arthropoda, Crustacea | |||||||
| Branchiopoda, Cladocera, Anomopoda | |||||||
| Daphniidae | |||||||
| Daphnia (Daphnia) sp. (ephippium) | aq | 74 | 14 | 130 | 10 | 4 | 22 |
| Simocephalus sp. (ephippium) | aq | 0 | 0 | 0 | 0 | 0 | 1 |
| Ceriodaphnia sp. (ephippium) | aq | 0 | 0 | 2 | 0 | 0 | 0 |
| Chydoridae | |||||||
| Alona affinis Leydig? | aq | 0 | 0 | 0 | 13 | 0 | 0 |
| Amphipoda, Gammaridae | |||||||
| Gammarus sp.? | aq | 0 | 1 | 0 | 0 | 0 | 0 |
| Annelida, Clitellata, Haplotaxida | |||||||
| Lumbricidae | |||||||
| Eisenia nordenskioldi (Eisen) | pl | 0 | 0 | 0 | 26 | 1 | 24 |
| Lumbricidae cocoons | pl | 9 | 4 | 9 | 0 | 11 | 0 |
| Annelida, Clitellata, Hirudinea, Arhynchobdellida | |||||||
| Erpobdellidae | |||||||
| Erpobdella octoculata (L)? | aq | 66 | 4 | 57 | 64 | 163 | 86 |
| Haemopidae | |||||||
| Haemopis sanguisuga (L)? | aq | 12 | 0 | 1 | 5 | 19 | 6 |
| Platyhelminthes, Rhabditophora, Tricladida | |||||||
| Planariidae | |||||||
| Planarian eggs | aq | 0 | 0 | 0 | 0 | 1 | 0 |
| Bryozoa, Phylactolaemata | |||||||
| Cristatellidae | |||||||
| Cristatella mucedo Cuvier | aq | 6 | 1 | 4 | 3 | 8 | 15 |
| Mollusca, Bivalvia | |||||||
| Sphaeriidae | |||||||
| Pisidium casertanum (Poli) | aq | 0 | 0 | 0 | 0 | 2 | 0 |
| Porifera, Demospongiae, Spongillida | |||||||
| Spongillidae | |||||||
| Spongillidae gen. indet. | aq | 0 | 0 | 0 | 0 | 37 | 24 |
| sum | 204 | 39 | 225 | 170 | 296 | 233 | |
| Taxa | Group | KS-21-1-1 + 2 | KS-21-1-3 | 544-1 | 544-2 | 544-3 |
|---|---|---|---|---|---|---|
| Pinaceae | ||||||
| Larix gmelinii (Rupr.) Kuzen | t | 1n | 3c, 8s, 1ss, 1n | 8s, 1n | 11s, 1ss, 1n | 1c, 1s |
| Picea obovata Ledeb. | t | 1c, 8s, 3ss, 3n | 28s | 36s, 1n | 12s, 2n | 11s |
| Pinus sylvestris L. | t | 1s | 5s | 1s | ||
| Betulaceae | ||||||
| Betula (s. str.) sect. Betula sp. | t | 5s | 1s | 1s | 1s | 6s |
| B. platyphylla Sukaczev | t | 1ss | 1ss | 1ss | ||
| B. fruticosa Pall. | sh | 1ss | ||||
| B. cf. fruticosa Pall. | sh | 1s | 1s | 1s | ||
| B. (s. str.) sect. Apterocaryon sp. | sh | 1s | 2s | |||
| Alnus hirsuta (Spach) Turcz. ex Rupr. | sh | 4s | 1s, 1b | 2s | 1s, 4ss | 1s |
| Rosaceae | ||||||
| Rubus cf. matsumuranus H. Lev. & Van. | sh | 1s | 1s | |||
| Potentilla sp. | oth | 2s | ||||
| Comarum palustre L. | hm | 3s | 22s | 4s | ||
| Ericaceae | ||||||
| Arctostaphylos uva-ursi (L.) Spreng. | psh | 2s | 1s | 5s | 2s | |
| Vaccinium uliginosum L. | psh | 1s | ||||
| Nymphaeaceae | ||||||
| Nuphar pumila (Timm) DC | h* | 2s | 16s | 1s | 2s | |
| Nymphaea tetragona Georgi | h* | 14s | 3s | 2s | 2s | 6s |
| Typhaceae | ||||||
| Sparganium emersum Rehmann | h | 1s | ||||
| S. hyperboreum Laest. | h | 1s | ||||
| Potamogetonaceae | ||||||
| Potamogeton perfoliatus L. | h | 48s | 470s | 6s | 21s | 37s |
| P. filiformis Pers. | h | 3s | 3s | |||
| P. natans L. | h | 1s | ||||
| Cyperaceae | ||||||
| Schoenoplectus lacustris (L.) Palla | h | 1s | 1s | |||
| Carex spp. | hm | 7s, 3ps | 7s, 5ps | 19s, 8ps | 7s, 7ps | 2ps |
| Eriophorum angustifolium Honck. | hm | 1s | ||||
| Ranunculaceae | ||||||
| Ranunculus repens L. | m | 1s | ||||
| R. monophyllus Ovcz. | m | 2s | 2s | |||
| Batrachium sp. | h | 1s | 1s | |||
| Haloragaceae | ||||||
| Myriophyllum verticillatum L. | h | 1s | 1s | |||
| Polygonaceae | ||||||
| Persicaria lapathifolia (L.) S.F. Gray | hm | 1s | ||||
| Apiaceae | ||||||
| Cicuta virosa L. | hm | 2s | 5s | 3s | ||
| Menyanthaceae | ||||||
| Menyanthes trifoliata L. | hm | 28s | 185s | 155s | 95s | 16s |
| Brassicaceae | ||||||
| Bunias cochlearioides Murray | xe | 1s | ||||
| Brassicaceae gen. indet | oth | 1s | ||||
| Lamiaceae | ||||||
| Dracocephalum ruyschiana L. | xe | 1s | ||||
| Lamiaceae gen. indet. | oth | 1s | ||||
| Amaranthaceae | ||||||
| Corispermum crassifolium Turcz. | p | 1s | ||||
| Bryales gen. indet. | hm | 4st | 3st | 5st | 8st | 1st |
| sum | 158 | 746 | 285 | 192 | 113 |
| Sample Code | Charcoalified Plant Remains, Number of Fragments | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Max Size (mm) | Circularity | Wood | Bark | Roots | Needles | Seeds | Fungal Sclerotia | Total | Vitrified Charcoal | |
| 544-1 | 49 | 49 | ||||||||
| KS-21-1-3 | 9 | 0.64 | 50 | 15 | 1 | 4 | 90 | 160 | 18 | |
| 544-2 | 10 | 27 | 3 | 4 | 13 | 53 | 5 * | |||
| KS-21-1-2 | 7 | 0.43 | 6 | 43 | 49 | |||||
| 544-3 | 7 | 0.64 | 3 | 6 | 1 | 22 | 32 | 9 | ||
| KS-21-1-1 | 0.52 | 9 | 8 | 1 | 6 | 24 | 8 | |||
| Sample | Betula | Ericaceae | Pinus | Picea/Larix | Sclerotia of | |
|---|---|---|---|---|---|---|
| Mycorrhizal Fungi (Cenococcum Type) | Saprobic Fungi (Collybia Type) | |||||
| 544-1 | 32 | 17 | ||||
| KS-21-1-3 | 4 | 2 | 4 | 40 | 18 | 72 |
| 544-2 | 1 | 6 | 20 * | 13 | ||
| KS-21-1-2 | 4 | 2 | 36 | 7 | ||
| 544-3 | 2 | 2 | 12 | 10 ** | ||
| KS-21-1-1 | 3 | 6 | 5 | 1 ** | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuzmina, S.A.; Micharevich, M.V.; Basilyan, A.E.; Lytkin, V.M.; Shaposhnikov, G.I.; Vasilyeva, A.N.; Pavlova, M.P.; Ponomarenko, E.; Galanin, A.A. Reconstruction of the Late Holocene Environment of Central Yakutia Based on Fossil Invertebrates and Plants from a Buried Lake at the Vilyuy River Valley. Water 2023, 15, 2790. https://doi.org/10.3390/w15152790
Kuzmina SA, Micharevich MV, Basilyan AE, Lytkin VM, Shaposhnikov GI, Vasilyeva AN, Pavlova MP, Ponomarenko E, Galanin AA. Reconstruction of the Late Holocene Environment of Central Yakutia Based on Fossil Invertebrates and Plants from a Buried Lake at the Vilyuy River Valley. Water. 2023; 15(15):2790. https://doi.org/10.3390/w15152790
Chicago/Turabian StyleKuzmina, S. A., M. V. Micharevich, A. E. Basilyan, V. M. Lytkin, G. I. Shaposhnikov, A. N. Vasilyeva, M. P. Pavlova, E. Ponomarenko, and A. A. Galanin. 2023. "Reconstruction of the Late Holocene Environment of Central Yakutia Based on Fossil Invertebrates and Plants from a Buried Lake at the Vilyuy River Valley" Water 15, no. 15: 2790. https://doi.org/10.3390/w15152790