
Temporal Changes in Water Quality with Increasing Ambient Temperatures Affect the Distribution and Relative Abundance of 10 Species of Balitorid Fishes in Small Streams of Eastern Thailand
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Physicochemical Measurement
2.2. Fish Samples, Species Richness, and Relative Abundance
2.3. Statistical Analysis
3. Results
3.1. Environmental Data, Species Richness and Abundance of Balitorids
3.2. CCA Analysis of Balitorid Distribution, Abundance and Physicochemical Variables in Water
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gheewala, S.H.; Silalertruksa, T.; Nilsalab, P.; Lecksiwilai, N.; Sawaengsak, W.; Mungkung, R.; Ganasut, J. Water stress index and its implication for agricultural land-use policy in Thailand. Int. J. Environ. Sci. Technol. 2018, 15, 833–846. [Google Scholar] [CrossRef]
- Leelahakriengkrai, P.; Peerapornpisal, Y. Water quality and trophic status in main rivers of Thailand. Chiang Mai J. Sci. 2011, 38, 280–294. [Google Scholar]
- Amnuaylojaroen, T.; Limsakul, A.; Kirtsaeng, S.; Parasin, N.; Surapipith, V. Effect of the near-future climate change under RCP8. 5 on the heat stress and associated work performance in Thailand. Atmosphere 2022, 13, 325. [Google Scholar] [CrossRef]
- Li, H.Y.; Xu, J.; Xu, R.Q. The effect of temperature on the water quality of lake. Adv. Mater. Res. 2013, 821, 1001–1004. [Google Scholar] [CrossRef]
- Cheng, X.; Huang, Y.; Li, R.; Pu, X.; Huang, W.; Yuan, X. Impacts of water temperature on phosphorus release of sediments under flowing overlying water. J. Contam. Hydrol. 2020, 235, 103717. [Google Scholar] [CrossRef]
- Wagner, T.; Lottig, N.R.; Bartley, M.L.; Hanks, E.M.; Schliep, E.M.; Wikle, N.B.; King, K.B.; McCullough, I.; Stachelek, J.; Cheruvelil, K.S.; et al. Increasing accuracy of lake nutrient predictions in thousands of lakes by leveraging water clarity data. Limnol. Oceanogr. Lett. 2020, 5, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, J.; Levy, J.; Kempe, S. Increasing dissolved silica trends in the Rhine River: An effect of recovery from high P loads? Limnology 2011, 12, 63–73. [Google Scholar] [CrossRef]
- Muylaert, K.; Sanchez-Pérez, J.M.; Teissier, S.; Sauvage, S.; Dauta, A.; Vervier, P. Eutrophication and its effect on dissolved Si concentrations in the Garonne River (France). J. Limnol. 2009, 68, 368–374. [Google Scholar] [CrossRef]
- Emerson, K.; Russo, R.C.; Lund, R.E.; Thurston, R.V. Aqueous ammonia equilibrium calculations: Effect of pH and temperature. J. Fish. Res. Board Can. 1975, 32, 2379–2383. [Google Scholar] [CrossRef]
- Czeglédi, I.; Specziár, A.; Erős, T. Temporal dynamics of freshwater fish assemblages, their background and methods of quantifications—A synthesis. Fish Fish. 2022, 23, 93–108. [Google Scholar] [CrossRef]
- Horka, P.; Musilova, Z.; Holubova, K.; Jandova, K.; Kukla, J.; Rutkayova, J.; Jones, J.I. Anthropogenic nutrient loading affects both individual species and the trophic structure of river fish communities. Front. Ecol. Evol. 2023, 10, 1297. [Google Scholar] [CrossRef]
- Martin-Smith, K.M. Relationships between fishes and habitat in rainforest streams in Sabah, Malaysia. J. Fish Biol. 1998, 52, 458–482. [Google Scholar] [CrossRef]
- Plongsesthee, R.; Kottelat, M.; Beamish, F.W.H. Schistura crocotula, a new loach from Peninsular Thailand (Teleostei: Nemacheilidae). Ichthyol. Explor. Freshw. 2013, 24, 171–178. [Google Scholar]
- Yang, G.Y.; Dudgeon, D. Dietary variation and food selection by an algivorous loach (Pseudogastromyzon myersi: Balitoridae) in Hong Kong streams. Mar. Freshw. Res. 2010, 61, 49–56. [Google Scholar] [CrossRef]
- Smith, H.M. The fresh-water fishes of Siam, or Thailand. Bull. U. S. Natn. Mus. 1945, 188, 1–622. [Google Scholar] [CrossRef]
- Vidthayanon, C.; Kottelat, M. Three new species of fishes from Tham Phra Wang Daeng and Tham Phra Sai Ngam caves in northern Thailand (Teleostei: Cyprinidae and Balitoridae). Ichthyol. Explor. Freshw. 2003, 14, 159–174. [Google Scholar]
- Havird, J.C.; Page, L.M.; Tangjitjaroen, W.; Vidthayanon, C.; Grudpan, C.; Udduang, S. A new species of Lepidocephalichthys (Teleostei: Cobitidae) with distinctive sexual dimorphism and comments on relationships in southern lineages of Cobitidae. Zootaxa 2010, 2557, 1–18. [Google Scholar] [CrossRef]
- Singer, R.A.; Page, L.M. Revision of the zipper loaches, Acanthocobitis and Paracanthocobitis (Teleostei: Nemacheilidae), with descriptions of five new species. Copeia 2015, 2015, 378–401. [Google Scholar] [CrossRef]
- Havird, J.C.; Page, L.M. A revision of Lepidocephalichthys (Teleostei: Cobitidae), with descriptions of two new species from Thailand and Myanmar. Copeia 2010, 2010, 137–159. [Google Scholar] [CrossRef]
- Plongsesthee, R.; Page, L.M.; Beamish, W. Schistura aurantiaca, a new species from the Mae Khlong basin, Thailand (Teleostei: Nemacheilidae). Ichthyol. Explor. Freshw. 2011, 22, 169. [Google Scholar]
- Kangrang, P.; Page, L.M.; Beamish, F.W.H. Schistura tenebrosa, a new species from the Kwai Noi River System, Mae Khlong basin, Thailand (Teleostei: Nemacheilidae). Zootaxa 2012, 3586, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Page, L.M.; Plongsesthee, R.; Beamish, F.W.H.; Kangrang, P.; Randall, Z.S.; Singer, R.; Martin, Z.P. Schistura (Teleostei: Nemacheilidae) in the Mae Khlong basin in southwestern Thailand with description of a new species. Zootaxza 2012, 3586, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Singer, R.A.; Pfeiffer, J.M.; Page, L.M. A revision of the Paracanthocobitis zonalternans (Cypriniformes: Nemacheilidae) species complex with descriptions of three new species. Zootaxa 2017, 4324, 85–107. [Google Scholar] [CrossRef] [Green Version]
- Beamish, F.W.H.; Sa-ardrit, P.; Cheevaporn, V. Habitat and abundance of Balitoridae in small rivers in central Thailand. J. Fish Biol. 2008, 72, 2467–2484. [Google Scholar] [CrossRef]
- Beamish, F.W.H.; Plongsesthee, R. Species diversity, abundance and environmental associations of loaches (Nemacheilidae and Balitoridae) in the central region of Thailand. Nat. Hist. Bull. Siam Soc. 2015, 60, 89–111. [Google Scholar]
- Nithirojpakdee, P.; Beamish, F.W.H.; Boonphakdee, T. Diet diversity among five co-existing fish species in a tropical river: Integration of dietary and stable isotope data. Limnology 2014, 15, 99–107. [Google Scholar] [CrossRef]
- Hayden, B.; Tongnunui, S.; Beamish, F.W.H.; Nithirojpakdee, P.; Cunjak, R.A. Variation in stable-isotope ratios between fin and muscle tissues can alter assessment of resource use in tropical river fishes. J. Fish Biol. 2017, 91, 574–586. [Google Scholar] [CrossRef]
- Hayden, B.; Tongnunui, S.; Beamish, F.W.H.; Nithirojpakdee, P.; Soto, D.X.; Cunjak, R.A. Functional and trophic diversity of tropical headwater stream communities inferred from carbon, nitrogen and hydrogen stable isotope ratios. Food Webs 2021, 26, e00181. [Google Scholar] [CrossRef]
- Dundgeon, D. Niche specificities of four fish species (Homalopteridae, Cobitidae, Gobiidae) from a Hong Kong forest stream. Arch. Hydrobiol. 1987, 108, 349–364. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater, 23rd ed.; American Public Health Association: Washington, DC, USA, 2017. [Google Scholar]
- Giller, P.S.; Malmqvist, B. The Biology of Streams and Rivers; Oxford University Press: Oxford, UK, 1998; pp. 1–296. [Google Scholar]
- Tongnunui, S.; Beamish, F.W.H. Habitat and relative abundance of fishes in small rivers in eastern Thailand. Environ. Biol. Fishes 2009, 85, 209–220. [Google Scholar] [CrossRef]
- Tongnunui, S.; Beamish, F.W.H.; Kongchaiya, C. Fish species, relative abundances and environmental associations in small rivers of the Mae Klong river basin in Thailand. Agric. Nat. Resour. 2016, 50, 408–415. [Google Scholar] [CrossRef]
- Carle, F.L.; Strub, M.S. A new method for estimating population size from removal data. Biometrics 1987, 34, 621–630. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Eschmeyer’ s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 1 January 2023).
- Boyd, D.A.; Nithirojpakdee, P.; Deein, G.; Vidthayanon, C.; Grudpan, C.; Tangjitjaroen, W.; Pfeiffer, J.H.; Randall, Z.S.; Srisombat, T.; Page, L.M. Revision of the horseface loaches (Cobitidae, Acantopsis), with descriptions of three new species from Southeast Asia. Zootaxa 2017, 4341, 151–192. [Google Scholar] [CrossRef] [Green Version]
- Kottelat, M. Indochinese Nemacheilines: A Revision of Nemacheiline Loaches (Pisces: Cypriniformes) of Thailand, Burma, Laos, Cambodia and Southern Viet Nam; Friedrich Pfeil: München, Germany, 1990; pp. 1–262. [Google Scholar]
- Singer, R.A.; Page, L.M. Redescription of Schistura myrmekia (Fowler 1935) (Teleostei: Nemacheilidae). Zootaxa 2012, 3586, 347–352. [Google Scholar] [CrossRef]
- Randall, Z.S.; Page, L.M. Resurrection of the genus Homalopteroides (Teleostei: Balitoridae) with a redescription of H. modestus (Vinciguerra 1890). Zootaxa 2012, 3586, 329–346. [Google Scholar] [CrossRef] [Green Version]
- Randall, Z.S.; Page, L.M. On the paraphyly of Homaloptera (Teleostei: Balitoridae) and description of a new genus of hillstream loaches from the Western Ghats of India. Zootaxa 2015, 3926, 57–86. [Google Scholar] [CrossRef] [Green Version]
- Randall, Z.S.; Somarriba, G.A.; Tongnunui, S.; Page, L.M. Review of the spotted lizard loaches, Pseudohomaloptera (Cypriniformes: Balitoridae) with a re-description of Pseudohomaloptera sexmaculata and description of a new species from Sumatra. J. Fish Biol. 2023, 102, 225–240. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics; McGraw-Hill Book Company: New York, NY, USA, 1980. [Google Scholar]
- Mccune, B.; Mefford, M.J. PC-ORD, Multivariate Analysis of Ecological Data, Version 4; MjM Software Design: Gleneden Beach, OR, USA, 1999.
- Clarke, K.R.; Gorley, R.N. Primer V5: User Manual/Tutorial; Primer-E Ltd.: Plymouth, MA, USA, 2006; pp. 1–93. [Google Scholar]
- Clarke, K.R.; Somerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Olson, J.C.; Appeldoorn, R.S.; Schärer-Umpierre, M.T.; Cruz-Motta, J.J. Recovery when you are on your own: Slow population responses in an isolated marine reserve. PLoS ONE 2019, 14, e0223102. [Google Scholar]
- Vieira, D.C.; Fonseca, G. The importance of vertical and horizontal dimensions of the sediment matrix in structuring nematodes across spatial scales. PLoS ONE 2013, 8, e77704. [Google Scholar] [CrossRef] [Green Version]
- Hayes, D.B.; Ferreri, C.P.; Taylor, W.W. Linking fish habitat to their population dynamics. Can. J. Fish. Aquat. Sci. 2006, 53, 383–390. [Google Scholar] [CrossRef]
- Kottelat, M. Zoogeography of the fishes from Indochinese inland waters with an annotated check-list. Bull. Zool. Mus. 1989, 12, 1–55. [Google Scholar]
- Freyhof, J.; Serov, D.V. Nemacheiline loaches from Central Vietnam with descriptions of a new genus and 14 new species (Cypriniformes: Balitoridae). Ichthyol. Explor. Freshw. 2011, 12, 133–191. [Google Scholar]
- Kottelat, M. Diagnoses of a new genus and 64 new species of fishes from Laos (Teleostei: Cyprinidae, Balitoridae, Bagridae, Syngnathidae, Chaudhuriidae and Tetraodontidae). J. S. Asian Nat. Hist. 2000, 5, 37–82. [Google Scholar]
- Page, L.M.; Pfeiffer, J.M.; Suksri, S.; Randall, Z.S.; Boyd, D.A. Variation in the arrow loach, Nemacheilus masyae (Cypriniformes: Nemacheilidae), in mainland Southeast Asia with description of a new species. Copeia 2020, 108, 392–402. [Google Scholar] [CrossRef]
- Nithirojpakdee, P.; Plongsesthee, R.; Beamish, F.W.H.; Kangrang, P. Science and conservation of aquatic animals in Thailand. Int. J. Environ. Sci. Rural Dev. 2012, 3, 50–55. [Google Scholar]
- Ward-Campbell, M.S.; Beamish, F.W.H.; Kongchaiya, C. Morphological characteristics in relation to diet in five coexisting Thai fish species. J. Fish Biol. 2005, 67, 1266–1279. [Google Scholar] [CrossRef]
- Wetzel, R.G. Limnology: Lake and River Ecosystems; Gulf Professional Publishing: Boston, MA, USA, 2001; pp. 1–1006. [Google Scholar]
- Israeli, Y.; Emmanuel, S. Impact of grain size and rock composition on simulated rock weathering. Earth Surf. Dyn. 2018, 6, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Ballah, M.; Bhoyroo, V.; Neetoo, H. Assessment of the physico-chemical quality and extent of algal proliferation in water from an impounding reservoir prone to eutrophication. J. Ecol. Environ. 2019, 43, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nithirojpakdee, P.; Beamish, F.W.H.; Noakes, D.L. Maintaining fish diversity in Thailand: Variations in foraging behavior. Environ. Biol. Fishes 2012, 95, 227–236. [Google Scholar] [CrossRef]
- Toetz, D.W. Agricultural Impacts on Stream Water Quality; Oklahoma Water Resources Center, Oklahoma State University: Stillwater, OK, USA, 1990. [Google Scholar]
- Matthews, W.J.; Gelwick, F.P. Fishes of Crutcho Creek and the north Canadian river in central Oklahoma: Effects of urbanization. Southwest. Nat. 1990, 34, 403–410. [Google Scholar] [CrossRef]
- Gautama, R.K.; Sharma, S.K.; Mahiyab, S.; Chattopadhyaya, M.C. Contamination of heavy metals in aquatic media: Transport, toxicity and technologies for remediation. In Heavy Metals in Water: Presence, Removal and Safety; Sanjay, S., Ed.; The Royal Society of Chemistry: London, UK, 2014; pp. 1–24. [Google Scholar]
- Mehta, K. Impact of temperature on contaminants toxicity in fish fauna: A review. Indian J. Sci. Technol. 2017, 10, 1–6. [Google Scholar] [CrossRef]
- Olson, P.E. Metallothioneins in fish: Induction and use in environmental monitoring. In Toxicology of Aquatic Pollution: Physiological, Cellular and Molecular Approaches; Talor, E.W., Ed.; Cambridge University Press: Cambridge, UK, 1996; pp. 187–204. [Google Scholar]
- Robinson, C.L.; Tonn, W.M. Influence of environmental factors and piscivory in structuring fish assemblages of small Alberta lakes. Can. J. Fish. Aquat. Sci. 1989, 46, 81–89. [Google Scholar] [CrossRef]
- Edds, D.R. Fish assemblage structure and environmental correlates in Nepal’s Gandaki river. Copeia 1993, 1993, 48–60. [Google Scholar] [CrossRef]
- Jackson, D.A.; Harvey, H.H. Fish and benthic invertebrates: Community concordance and community–environment relationships. Can. J. Fish. Aquat. Sci. 1993, 50, 2641–2651. [Google Scholar] [CrossRef]
- Beamish, F.W.H.; Kangrang, P.; Nithirojpakdee, P.; Plongsesthee, R. Why Thai river fish occur where they are found. EnvironmentAsia 2012, 5, 1–16. [Google Scholar]
- Kang, B.; Deng, J.; Wu, Y.; Chen, L.; Zhang, J.; Qiu, H.; Lu, Y.; He, D. Mapping China’s freshwater fishes: Diversity and biogeography. Fish Fish. 2014, 15, 209–230. [Google Scholar] [CrossRef]
- Radinger, J.; Britton, J.R.; Carlson, S.M.; Magurran, A.E.; Alcaraz-Hernández, J.D.; Almodóvar, A.; Benejam, L.; Fernández-Delgado, C.; Nicola, G.G.; Oliva-Paterna, F.J.; et al. Effective monitoring of freshwater fish. Fish Fish. 2019, 20, 729–747. [Google Scholar] [CrossRef] [Green Version]
- Fry, F.E.J. The effects of the environmental factors on the physiology of fish. In Fish Physiology; Hoar, W.S., Randall, J.R., Brett, J.R., Eds.; Academic Press: London, UK, 1971; pp. 1–98. [Google Scholar]
- Hughes, R.M.; Gammon, J.R. Longitudinal changes in fish assemblages and water quality in the Willamette River, Oregon. Trans. Am. Fish. Soc. 1987, 116, 196–209. [Google Scholar] [CrossRef]
- Hughes, R.M.; Noss, R.F. Biological diversity and biological integrity: Current concerns for lakes and streams. Fisheries 1992, 17, 11–19. [Google Scholar] [CrossRef]
- Tsui, T.K.N.; Randall, D.J.; Chew, S.F.; Jin, Y.; Wilson, J.M.; Ip, Y.K. Accumulation of ammonia in the body and NH3 volatilization from alkaline regions of the body surface during ammonia loading and exposure to air in the weather loach Misgurnus anguillicaudatus. J. Exp. Biol. 2002, 205, 651–659. [Google Scholar] [CrossRef]
- Wang, Y.; Walsh, P.J. High ammonia tolerance in fishes of the family Batrachoididae (Toadfish and Midshipmen). Aquat. Toxicol. 2000, 50, 205–219. [Google Scholar] [CrossRef]
- Liu, M.J.; Guo, H.Y.; Zhu, K.C.; Liu, B.S.; Liu, B.; Guo, L.; Zhang, N.; Yang, J.W.; Jiang, S.G.; Zhang, D.C. Effects of acute ammonia exposure and recovery on the antioxidant response and expression of genes in the Nrf2-Keap1 signaling pathway in the juvenile golden pompano (Trachinotus ovatus). Aquat. Toxicol. 2021, 240, 105969. [Google Scholar] [CrossRef]
- Sharma, N.K.; Akhtar, M.S.; Pandey, N.; Singh, R.; Singh, A.K. Seasonal variation in thermal tolerance, oxygen consumption, antioxidative enzymes and non-specific immune indices of Indian hill trout, Barilius bendelisis (Hamilton, 1807) from central Himalaya, India. J. Therm. Biol. 2015, 52, 166–176. [Google Scholar] [CrossRef]
- Tongnunui, S.; Sooksawat, T.; Chotwiwatthanakun, C.; Supiwong, W.; Wattanakornsiri, A.; Beamish, F.W.H. Seasonal changes in upper thermal tolerances of freshwater Thai fishes. Water 2023, 15, 350. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
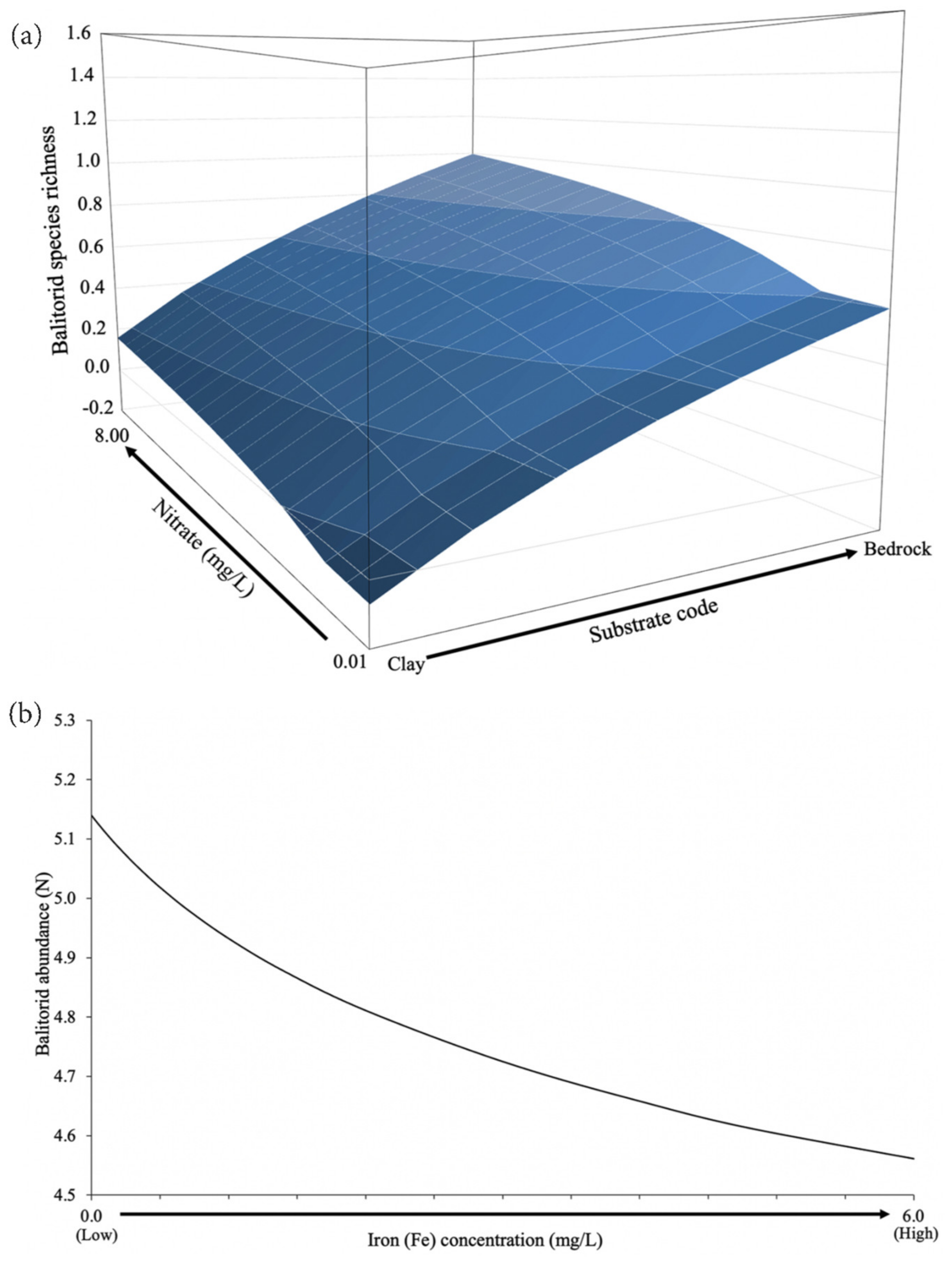{kind=link}
{kind=link}
{kind=link}
| Environmental Factors | GM ± SD | Environmental Factors | GM ± SD |
|---|---|---|---|
| Alkalinity, mg/L as CaCO3 | 127.1 ± 2.8 | Nitrate, mg NO3/L | 3.4 ± 2.3 |
| Ammonia, mg NH3·L−1 | 235.2 ± 7.6 | Oxygen, mg/L | 5.7 ± 1.4 |
| BOD, mg/L | 65.7 ± 7.8 | pH | 7.1 ± 0.7 |
| Canopy, % cover | 36.2 ± 4.1 | Silica, mg SiO2/L | 13.1 ± 3.5 |
| Conduct, μS/cm | 516.1 ± 2.88 | Substrate, (Code) | 3.5 ± 1.5 |
| Depth, cm | 22.0 ± 1.7 | Temperature, °C | 26.5 ± 1.1 |
| Discharge, L/s | 234.2 ± 5.5 | Turbid, NTU | 28.1 ± 2.4 |
| Elevation, m | 71.2 ± 2.0 | Velocity, cm/s | 23.5 ± 2.9 |
| Width, m | 3.7 ± 1.6 | Total Iron, mg Fe/L | 1.0 ± 1.4 |
| Length, m | 32.6 ± 1.8 | Phosphate, mg PO43−/L | 6.4 ± 2.4 |
| Color, CPU | 107.6 ± 9.6 |
| Species | ID | Sum of Average Fish | Relative Abundance | Occurrence Frequency (%) | |
|---|---|---|---|---|---|
| GM ± SD | Ranges | ||||
| Paracanthocobitis zonalternans (Blyth, 1860) | 1 | 86.1 | 0.86 ± 1.68 | 0.26–5.4 | 5.2 |
| Homalopteroides smithi (Hora, 1932) | 2 | 7.5 | 0.1 ± 1.23 | 0.7–3.1 | 4.2 |
| Homaloptera orthogoniata (Vaillant, 1902) | 3 | 1.3 | <0.1 | 0.0–0.2 | 3.1 |
| Nemacheilus masyae (Smith, 1933) | 4 | 6.1 | 0.1 ± 1.20 | 0.2–4 | 5.2 |
| Nemacheilus platiceps (Kottelat, 1990) | 5 | 8 | 0.1 ± 1.25 | 0.2–6.2 | 3.1 |
| Schistura kohchangensis (Smith, 1933) | 6 | 108.5 | 1.15 ± 1.88 | 0.8–66.7 | 5.2 |
| Acanthopsis rungthipae (Boyd et al., 2017) | 7 | 13.1 | 0.14 ± 1.31 | 0–13.1 | 1.1 |
| Acanthopsis cf. rungthipae (Boyd et al., 2017) | 8 | 31.2 | 0.33 ± 1.50 | 0.6–19.3 | 4.2 |
| Lepidocephalichthys berdmorei (Blyth, 1986) | 9 | 2.3 | <0.1 | 0.0–2.3 | 1.1 |
| Lepidocephalichthys hasselti (Valenciennes, 1846) | 10 | 179.6 | 2.0 ± 2.36 | 1–52.4 | 22.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tongnunui, S.; Beamish, F.W.H.; Sooksawat, T.; Wattanakornsiri, A.; Chotwiwatthanakun, C.; Supiwong, W.; Intacharoen, P.; Sudtongkong, C. Temporal Changes in Water Quality with Increasing Ambient Temperatures Affect the Distribution and Relative Abundance of 10 Species of Balitorid Fishes in Small Streams of Eastern Thailand. Water 2023, 15, 2791. https://doi.org/10.3390/w15152791
Tongnunui S, Beamish FWH, Sooksawat T, Wattanakornsiri A, Chotwiwatthanakun C, Supiwong W, Intacharoen P, Sudtongkong C. Temporal Changes in Water Quality with Increasing Ambient Temperatures Affect the Distribution and Relative Abundance of 10 Species of Balitorid Fishes in Small Streams of Eastern Thailand. Water. 2023; 15(15):2791. https://doi.org/10.3390/w15152791
Chicago/Turabian StyleTongnunui, Sampan, F. W. H. Beamish, Treerat Sooksawat, Amnuay Wattanakornsiri, Charoonroj Chotwiwatthanakun, Weerayuth Supiwong, Prasarn Intacharoen, and Chanyut Sudtongkong. 2023. "Temporal Changes in Water Quality with Increasing Ambient Temperatures Affect the Distribution and Relative Abundance of 10 Species of Balitorid Fishes in Small Streams of Eastern Thailand" Water 15, no. 15: 2791. https://doi.org/10.3390/w15152791