
Hydrogeomorphic Scaling and Ecohydraulics for Designing Rescaled Channel and Floodplain Geometry in Regulated Gravel–Cobble Bed Rivers for Pacific Salmon Habitat


Abstract
:1. Introduction
2. Hydrogeomorphic Scaling
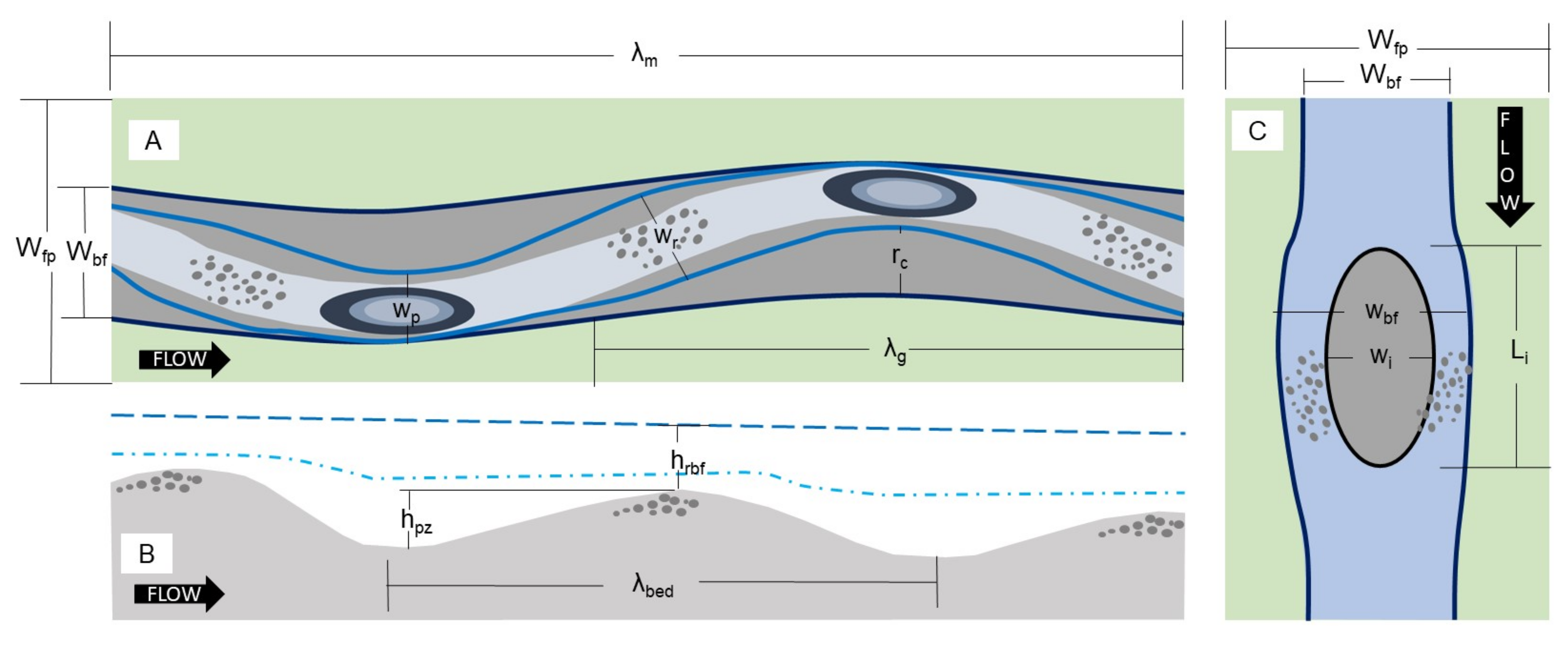
2.1. Scaling Variables
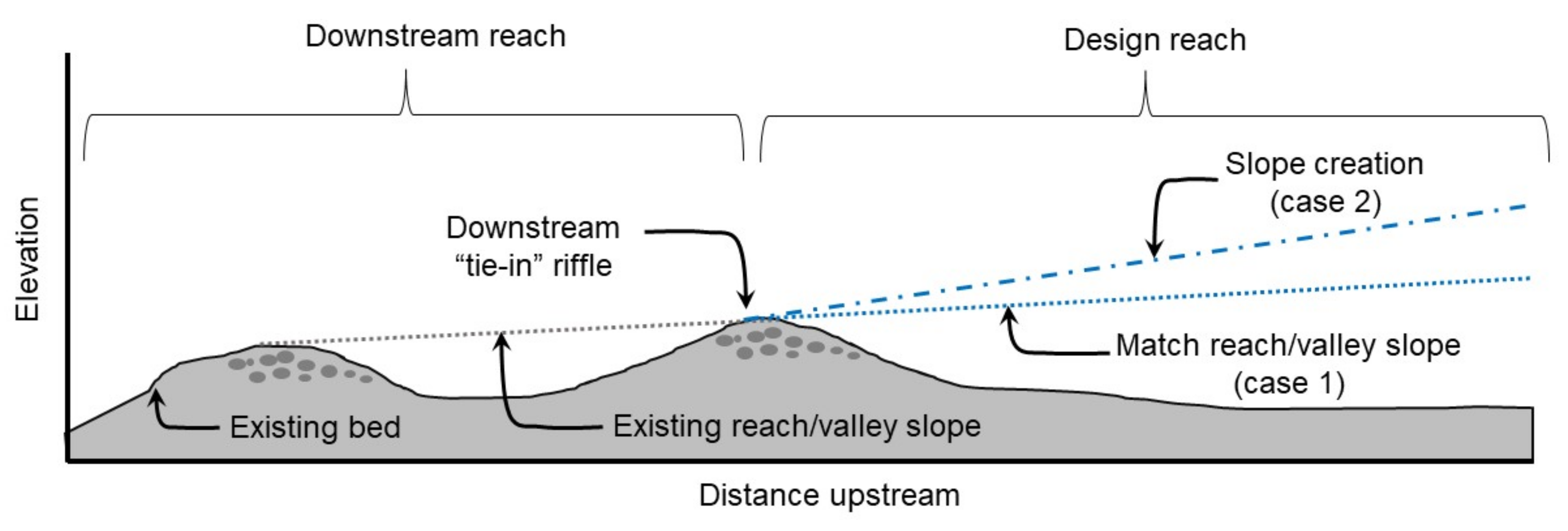
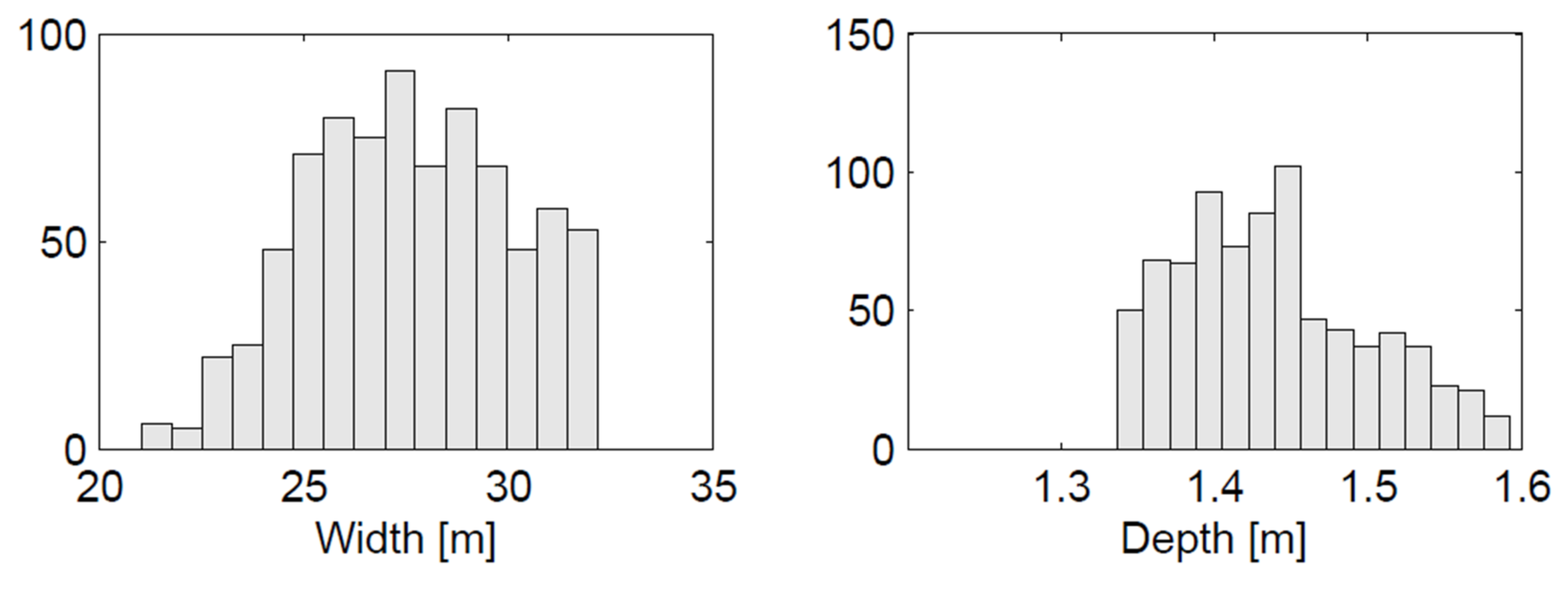
2.2. Channel Geometry
2.3. Planform Typology and Scaling
2.3.1. Alternate Bar Geometry
2.3.2. Anabranch Geometry
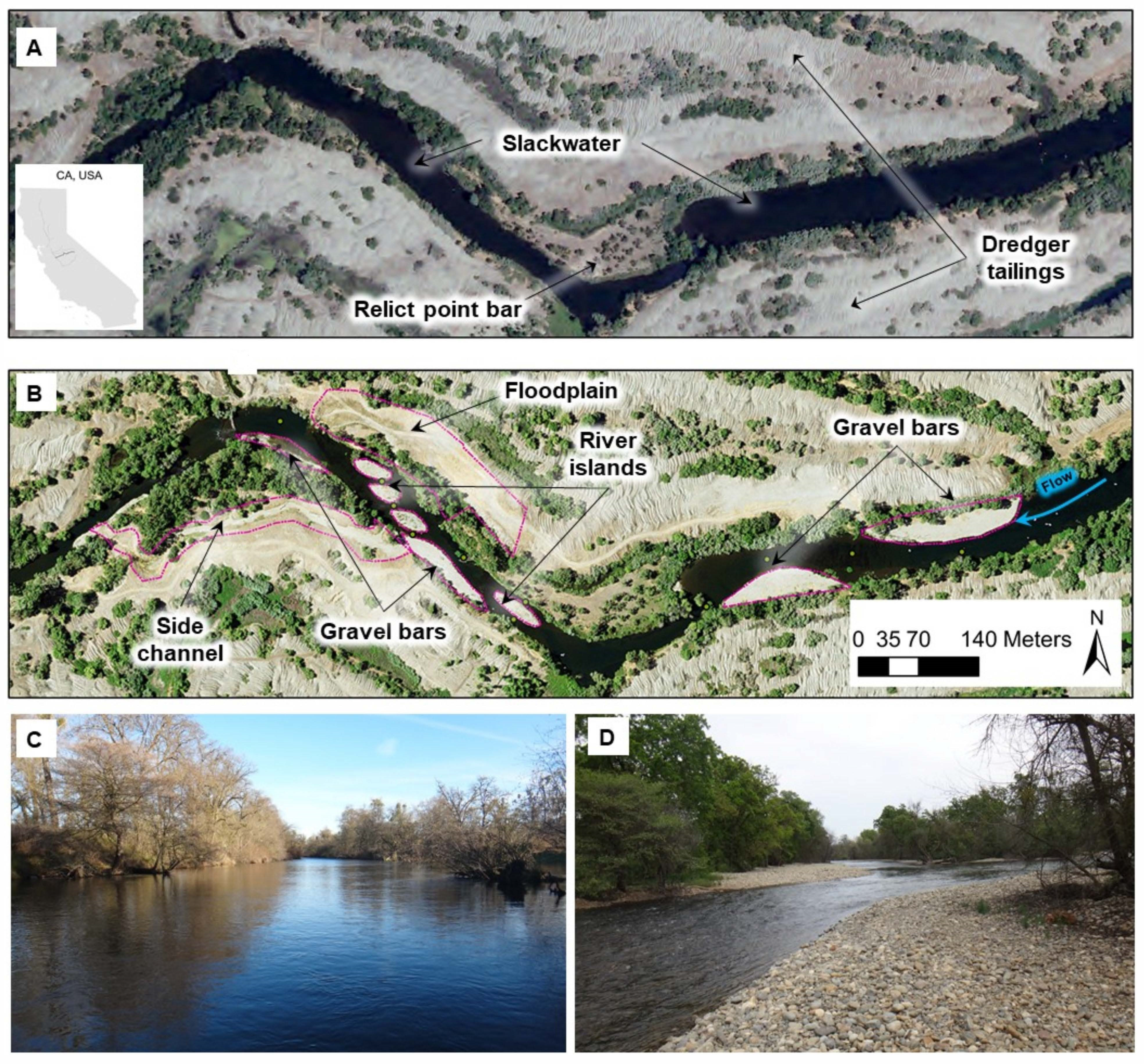
2.3.3. Floodplain and Side Channels
2.4. Pool–Riffle Variation
2.5. Which Hydrogeomorphic Scaling Relationships, or Does It Even Matter?
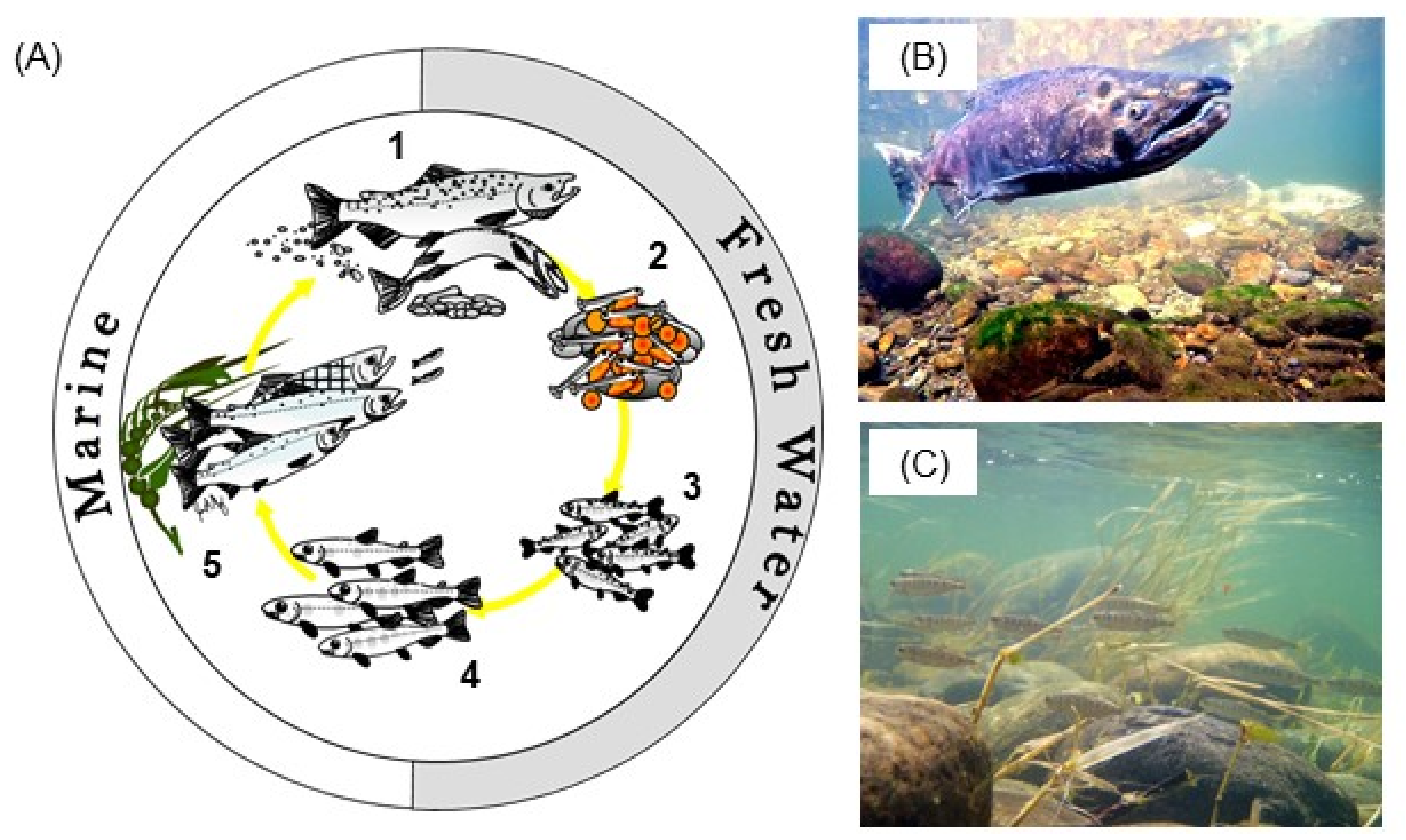
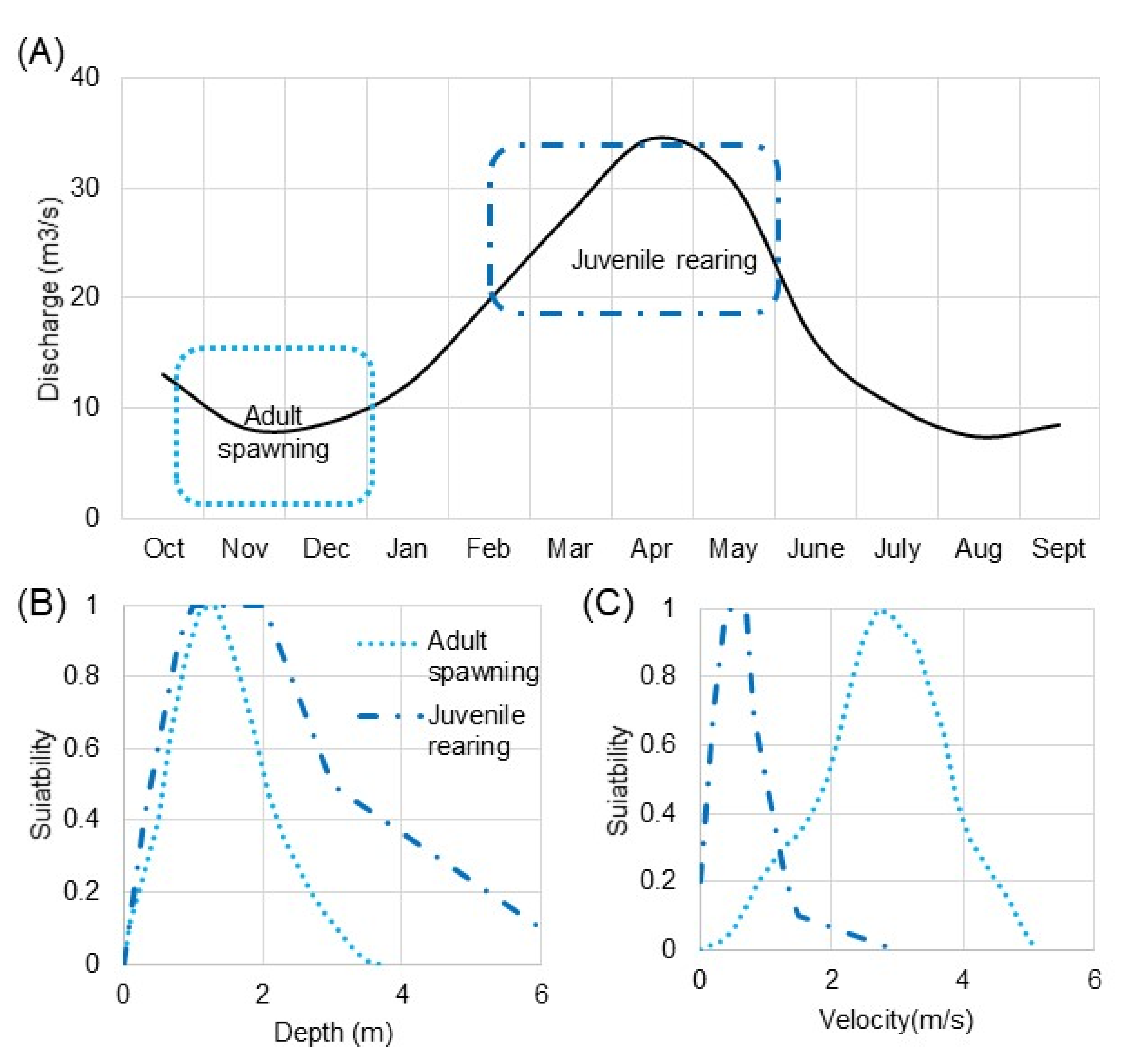
3. Ecohydraulic Scaling for Salmon Habitat

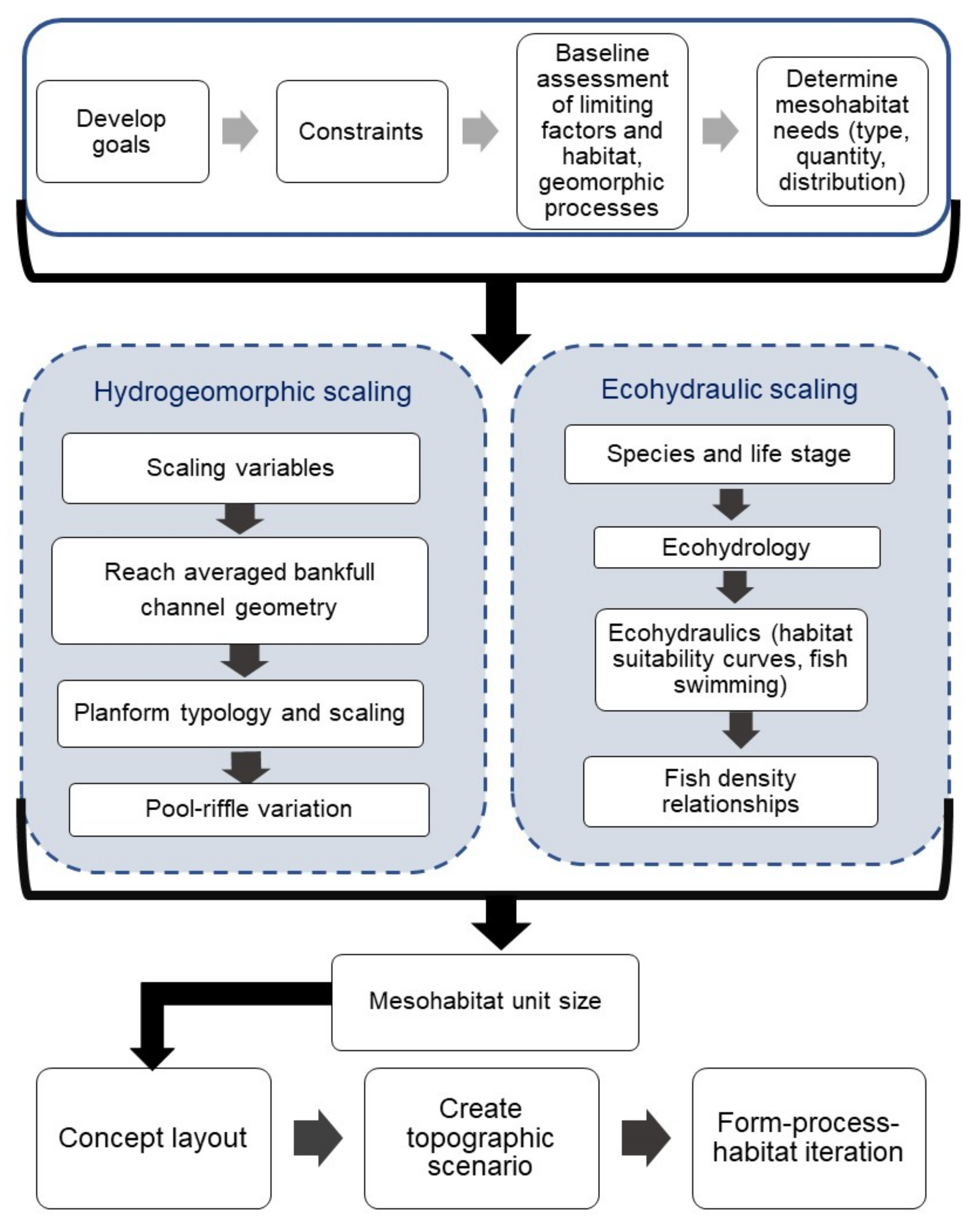
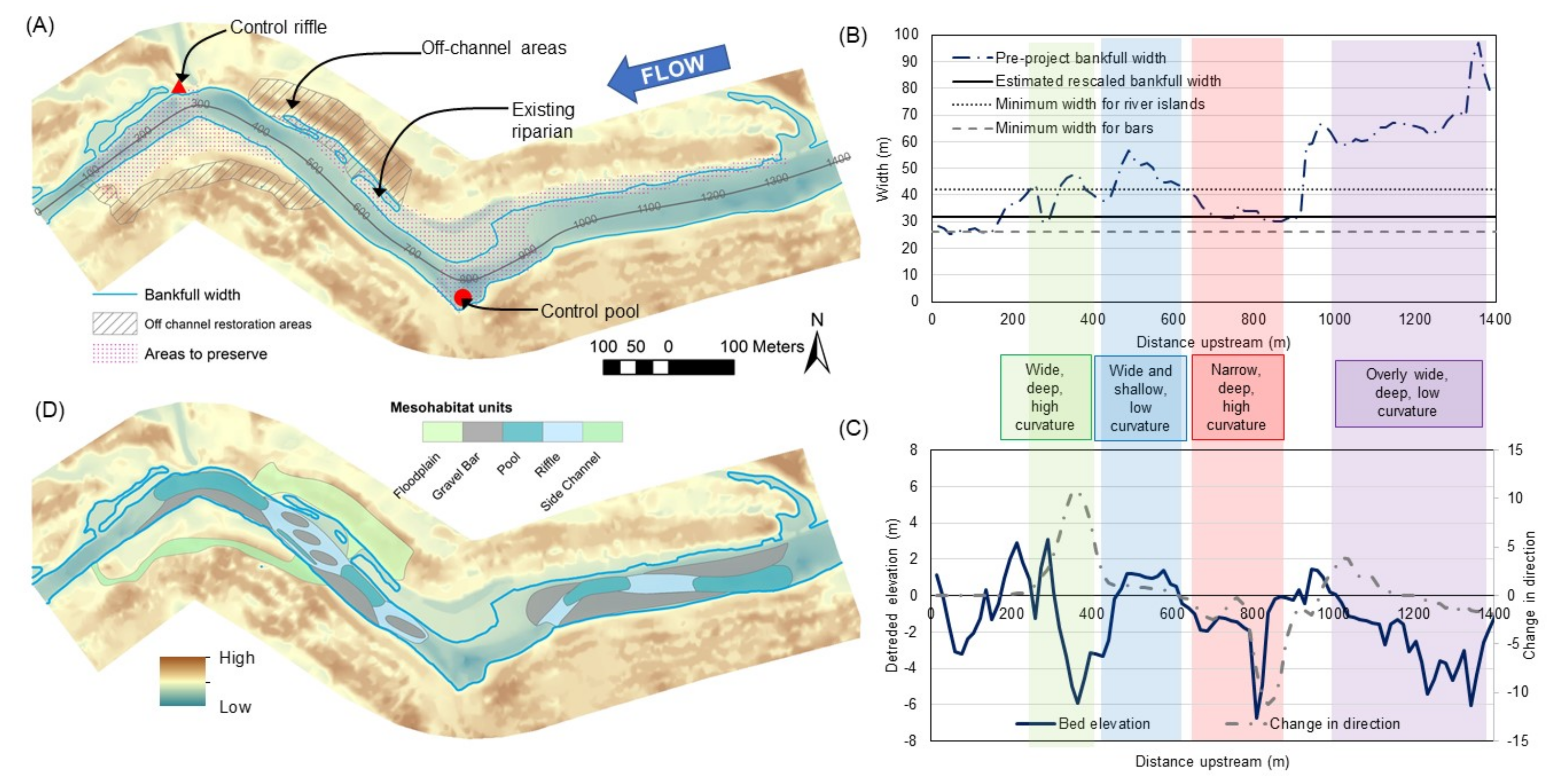
4. Creating Conceptual Mesohabitat Unit Designs
5. Discussion
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trush, W.J.; McBain, S.M.; Leopold, L.B. Attributes of an alluvial river and their relation to water policy and management. Proc. Natl. Acad. Sci. USA 2000, 97, 11858–11863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondolf, G.M.; Podolak, K.; Grantham, T.E. Restoring mediterranean-climate rivers. Hydrobiologia 2013, 719, 527–545. [Google Scholar] [CrossRef]
- Wheaton, J.M.; Pasternack, G.B.; Merz, J.E. Spawning habitat rehabilitation-I. Conceptual approach and methods. Int. J. River Basin Manag. 2004, 2, 3–20. [Google Scholar] [CrossRef]
- Wohl, E.; Lane, S.N.; Wilcox, A.C. The science and practice of river restoration. Water Resour. Res. 2015, 51, 5974–5997. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.F.; Thorne, C.R.; Castro, J.M.; Kondolf, G.M.; Mazzacano, C.S.; Rood, S.B.; Westbrook, C. Biomic river restoration: A new focus for river management. River Res. Appl. 2020, 36, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Pasternack, G.B. River Restoration: Disappointing, Nascent, Yet Desperately Needed. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Bernhardt, E.S.; Palmer, M.A.; Allan, J.D.; Alexander, G.; Barnas, K.; Brooks, S.; Carr, J.; Clayton, S.; Dahm, C.; Follstad-Shah, J.; et al. Synthesizing U.S. river restoration efforts. Science 2005, 308, 636–637. [Google Scholar] [CrossRef]
- Whipple, A.A.; Viers, J.H. Coupling landscapes and river flows to restore highly modified rivers. Water Resour. Res. 2019, 55, 4512–4532. [Google Scholar] [CrossRef]
- Merz, J.E.; Moyle, P.B. Salmon, wildlife, and wine: Marine-derived nutrients in human-dominated ecosystems of central California. Ecol. Appl. 2006, 16, 999–1009. [Google Scholar] [CrossRef] [Green Version]
- Stanford, J.A.; Ward, J.V.; Liss, W.J.; Frissell, C.A.; Williams, R.N.; Lichatowich, J.A.; Coutant, C.C. A General Protocol for Restoration Of Regulated Rivers. Regul. Rivers Res. Manag. 1996, 12, 391–413. [Google Scholar] [CrossRef]
- Kondolf, G.M. Hungry water: Effects of dams and gravel mining on river channels. Environ. Manag. 1997, 21, 533–551. [Google Scholar] [CrossRef]
- Graf, W.L. Downstream hydrologic and geomorphic effects of large dams on American rivers. Geomorphology 2006, 79, 336–360. [Google Scholar] [CrossRef]
- Grant, G.E. The Geomorphic Response of Gravel-Bed Rivers to Dams: Perspectives and Prospects. In Gravel-Bed Rivers: Processes, Tools, Environments; John Wiley & Sons: Chichester, UK; Hoboken, NJ, USA, 2012; pp. 165–181. ISBN 9780470688908. [Google Scholar]
- James, L.A.; Singer, M.B.; Ghoshal, S.; Megison, M. Historical channel changes in the lower Yuba and Feather Rivers, California: Long-term effects of contrasting river-management strategies. Spec. Pap. Geol. Soc. Am. 2009, 451, 7. [Google Scholar] [CrossRef] [Green Version]
- Tockner, K.; Stanford, J.A. Riverine flood plains: Present state and future trends. Environ. Conserv. 2002, 29, 308–330. [Google Scholar] [CrossRef] [Green Version]
- Yoshiyama, R.M.; Fisher, F.W.; Moyle, P.B. Historical Abundance and Decline of Chinook Salmon in the Central Valley Region of California. N. Am. J. Fish. Manag. 2004, 18, 487–521. [Google Scholar] [CrossRef]
- Hanrahan, T.P. Bedform morphology of salmon spawning areas in a large gravel-bed river. Geomorphology 2007, 86, 529–536. [Google Scholar] [CrossRef]
- Jacobson, R.B.; Galat, D.L. Flow and form in rehabilitation of large-river ecosystems: An example from the Lower Missouri River. Geomorphology 2006, 77, 249–269. [Google Scholar] [CrossRef]
- Brown, R.A.; Pasternack, G.B. Engineered channel controls limiting spawning habitat rehabilitation success on regulated gravel-bed rivers. Geomorphology 2008, 97, 631–654. [Google Scholar] [CrossRef] [Green Version]
- Tonkin, J.D.; Olden, J.D.; Merritt, D.M.; Reynolds, L.V.; Rogosch, J.S.; Lytle, D.A. Designing flow regimes to support entire river ecosystems. Front. Ecol. Environ. 2021, 19, 326–333. [Google Scholar] [CrossRef]
- Jager, H.I. Thinking outside the cHannel: Timing pulse flows to benefit salmon via indirect pathways. Ecol. Modell. 2014, 273, 117–127. [Google Scholar] [CrossRef]
- Hall, A.A.; Rood, S.B.; Higgins, P.S. Resizing a river: A downscaled, seasonal flow regime promotes riparian restoration. Restor. Ecol. 2011, 19, 351–359. [Google Scholar] [CrossRef]
- Thompson, D.M.; Stull, G.N. The Development and Historic Use of Habitat Structures in Channel Restoration in the United States: The Grand Experiment in Fisheries Management. Géographie Phys. Quat. 2004, 56, 45–60. [Google Scholar] [CrossRef] [Green Version]
- Ock, G.; Gaeuman, D.; McSloy, J.; Kondolf, G.M. Ecological functions of restored gravel bars, the Trinity River, California. Ecol. Eng. 2015, 83, 49–60. [Google Scholar] [CrossRef]
- Elkins, E.M.; Pasternack, G.B.; Merz, J.E. Use of slope creation for rehabilitating incised, regulated, gravel bed rivers. Water Resour. Res. 2007, 43, 1–16. [Google Scholar] [CrossRef]
- Pasternack, G.B.G.B.G.B.; Brown, R.A.R.A.R.A. Ecohydraulic Design of Riffle-Pool Relief and Morphological Unit Geometry in Support of Regulated Gravel-Bed River Rehabilitation. In Ecohydraulics: An Integrated Approach; Wiley: Hoboken, NJ, USA, 2013; pp. 337–355. ISBN 9781118526576. [Google Scholar]
- Schweizer, S.; Borsuk, M.E.; Reichert, P. Predicting the morphological and hydraulic consequences of river rehabilitation. River Res. Appl. 2007, 23, 303–322. [Google Scholar] [CrossRef]
- Morley, S.A.; Garcia, P.S.; Bennett, T.R.; Roni, P. Juvenile salmonid (Oncorhynchus spp.) use of constructed and natural side channels in Pacific Northwest rivers. Can. J. Fish. Aquat. Sci. 2005, 62, 2811–2821. [Google Scholar] [CrossRef]
- Gaeuman, D.; Stewart, R.; Schmandt, B.; Pryor, C. Geomorphic response to gravel augmentation and high-flow dam release in the Trinity River, California. Earth Surf. Process. Landforms 2017, 42, 2523–2540. [Google Scholar] [CrossRef]
- Merz, J.; Caldwell, L.; Beakes, M.; Hammersmark, C.; Sellheim, K. Balancing competing life-stage requirements in salmon habitat rehabilitation: Between a rock and a hard place. Restor. Ecol. 2019, 27, 661–671. [Google Scholar] [CrossRef]
- Mörtl, C.; De Cesare, G. Sediment Augmentation for River Rehabilitation and Management—A Review. Land 2021, 10, 1309. [Google Scholar] [CrossRef]
- James, L.A. Designing forward with an eye to the past: Morphogenesis of the lower Yuba River. Geomorphology 2015, 251, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, M.P.; Moyle, P.B. Effects of flow regime on fish assemblages in a regulated California stream. Ecol. Appl. 2001, 11, 530–539. [Google Scholar] [CrossRef]
- Kiernan, J.D.; Moyle, P.B.; Crain, P.K. Restoring native fish assemblages to a regulated California stream using the natural flow regime concept. Ecol. Appl. 2012, 22, 1475–1482. [Google Scholar] [CrossRef] [PubMed]
- Downs, P.W.; Singer, M.S.; Orr, B.K.; Diggory, Z.E.; Church, T.C. Restoring ecological integrity in highly regulated rivers: The role of baseline data and analytical references. Environ. Manag. 2011, 48, 847–864. [Google Scholar] [CrossRef] [PubMed]
- Harrison, L.R.; Legleiter, C.J.; Wydzga, M.A.; Dunne, T. Channel dynamics and habitat development in a meandering, gravel bed river. Water Resour. Res. 2011, 47, W04513. [Google Scholar] [CrossRef] [Green Version]
- Roni, P.; Hall, J.E.; Drenner, S.M.; Arterburn, D. Monitoring the effectiveness of floodplain habitat restoration: A review of methods and recommendations for future monitoring. WIREs Water 2019, 6, e1355. [Google Scholar] [CrossRef]
- Harrison, L.R.; Bray, E.; Overstreet, B.; Legleiter, C.J.; Brown, R.A.; Merz, J.E.; Bond, R.M.; Nicol, C.L.; Dunne, T. Physical Controls on Salmon Redd Site Selection in Restored Reaches of a Regulated, Gravel-Bed River. Water Resour. Res. 2019, 55, 8942–8966. [Google Scholar] [CrossRef]
- Cramer Fish Sciences. Merced River Ranch and Henderson Park Restoration Projects On The Merced River, California. Final Data Report; Cramer Fish Sciences: West Sacramento, CA, USA, 2019. [Google Scholar]
- Shields, F.D.; Copeland, R.R.; Klingeman, P.C.; Doyle, M.W.; Simon, A. Design for Stream Restoration. J. Hydraul. Eng. 2003, 129, 575–584. [Google Scholar] [CrossRef]
- Brown, R.A.; Pasternack, G.B.; Lin, T. The Topographic Design of River Channels for Form-Process Linkages. Environ. Manag. 2016, 57, 929–942. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.R.; Millar, R.G.; Newbury, R.W. Hydraulic design of riffles in gravel/cobble bed Rivers. Int. J. River Basin Manag. 2004, 2, 291–299. [Google Scholar] [CrossRef]
- Beechie, T.; Pess, G.; Roni, P.; Giannico, G. Setting River Restoration Priorities: A Review of Approaches and a General Protocol for Identifying and Prioritizing Actions. N. Am. J. Fish. Manag. 2008, 28, 891–905. [Google Scholar] [CrossRef]
- Wheaton, J.M.; Pasternack, G.B.; Merz, J.E. Spawning habitat rehabilitation-II. Using hypothesis development and testing in design, Mokelumne river, California, U.S.A. Int. J. River Basin Manag. 2004, 2, 21–37. [Google Scholar] [CrossRef]
- Merz, J.E.; Setka, J.D.; Pasternack, G.B.; Wheaton, J.M. Predicting benefits of spawning-habitat rehabilitation to salmonid (Oncorhynchus spp.) fry production in a regulated California river. Can. J. Fish. Aquat. Sci. 2004, 61, 1433–1446. [Google Scholar] [CrossRef] [Green Version]
- Sellheim, K.L.; Watry, C.B.; Rook, B.; Zeug, S.C.; Hannon, J.; Zimmerman, J.; Dove, K.; Merz, J.E. Juvenile Salmonid Utilization of Floodplain Rearing Habitat After Gravel Augmentation in a Regulated River. River Res. Appl. 2016, 32, 610–621. [Google Scholar] [CrossRef]
- Wheaton, J.M.; Brasington, J.; Darby, S.E.; Merz, J.; Pasternack, G.B.; Sear, D.; Vericat, D.; Joseph, M.W.; James, B.; Stephen, E.D.; et al. Linking geomorphic changes to salmonid habitat at a scale relevant to fish. River Res. Appl. 2010, 26, 469–486. [Google Scholar] [CrossRef]
- Brown, R.A.; Pasternack, G.B. How to build a digital river. Earth-Sci. Rev. 2019, 194, 283–305. [Google Scholar] [CrossRef]
- Wall, C.E.; Bouwes, N.; Wheaton, J.M.; Bennett, S.N.; Saunders, W.C.; McHugh, P.A.; Jordan, C.E. Design and monitoring of woody structures and their benefits to juvenile steelhead (Oncorhynchus mykiss) using a net rate of energy intake model. Can. J. Fish. Aquat. Sci. 2017, 74, 727–738. [Google Scholar] [CrossRef]
- Lane, B.A.; Pasternack, G.B.; Sandoval Solis, S. Integrated analysis of flow, form, and function for river management and design testing. Ecohydrology 2018, 11, e1969. [Google Scholar] [CrossRef] [Green Version]
- Wheaton, J.M.; Bouwes, N.; Mchugh, P.; Saunders, C.; Bangen, S.; Bailey, P.; Nahorniak, M.; Wall, E.; Jordan, C. Upscaling site-scale ecohydraulic models to inform salmonid population-level life cycle modeling and restoration actions—Lessons from the Columbia River Basin. Earth Surf. Process. Landforms 2017, 4137, 21–44. [Google Scholar] [CrossRef]
- Havis, R.N.; Alonso, C.V.; King, J.G. Modeling sediment in gravel-bedded streams using HEC-6. J. Hydraul. Eng. 1996, 122, 559–564. [Google Scholar] [CrossRef]
- Mosselman, E. Modelling Sediment Transport and Morphodynamics of Gravel-Bed Rivers. In Gravel-Bed Rivers: Processes, Tools, Environments; Wiley: Hoboken, NJ, USA, 2012; pp. 101–115. ISBN 9780470688908. [Google Scholar]
- Van De Wiel, M.J.; Coulthard, T.J.; Macklin, M.G.; Lewin, J. Embedding reach-scale fluvial dynamics within the CAESAR cellular automaton landscape evolution model. Geomorphology 2007, 90, 283–301. [Google Scholar] [CrossRef]
- Smith, S.M.; Prestegaard, K.L. Hydraulic performance of a morphology-based stream channel design. Water Resour. Res. 2005, 4, W11413. [Google Scholar] [CrossRef] [Green Version]
- Harper, D.; Ebrahimnezhad, M.; Climent I Cot, F. Artificial riffles in river rehabilitation: Setting the goals and measuring the successes. Aquat. Conserv. Mar. Freshw. Ecosyst. 1998, 8, 5–16. [Google Scholar] [CrossRef]
- Taylor Perron, J.; Fagherazzi, S. The legacy of initial conditions in landscape evolution. Earth Surf. Process. Landforms 2012, 37, 52–63. [Google Scholar] [CrossRef]
- Schwartz, J.S. Use of ecohydraulic-based mesohabitat classification and fish species traits for stream restoration design. Water 2016, 8, 520. [Google Scholar] [CrossRef] [Green Version]
- Fryirs, K.; Brierley, G. Assemblages of geomorphic units: A building block approach to analysis and interpretation of river character, behaviour, condition and recovery. Earth Surf. Process. Landforms 2021, 47, 92–108. [Google Scholar] [CrossRef]
- Beechie, T.J.; Sear, D.A.; Olden, J.D.; Pess, G.R.; Buffington, J.M.; Moir, H.; Roni, P.; Pollock, M.M. Process-based Principles for Restoring River Ecosystems. Bioscience 2010, 60, 209–222. [Google Scholar] [CrossRef] [Green Version]
- Leopold, L.B.; Wolman, M.G. River meanders. Bull. Geol. Soc. Am. 1960, 71, 769–793. [Google Scholar] [CrossRef]
- Jaeggi, M.N.R. Formation and Effects of Alternate Bars. J. Hydraul. Eng. 1984, 110, 142–156. [Google Scholar] [CrossRef]
- Crosato, A.; Mosselman, E. Simple physics-based predictor for the number of river bars and the transition between meandering and braiding. Water Resour. Res. 2009, 45, W03424. [Google Scholar] [CrossRef] [Green Version]
- Crosato, A.; Mosselman, E. An Integrated Review of River Bars for Engineering, Management and Transdisciplinary Research. Water 2020, 12, 596. [Google Scholar] [CrossRef] [Green Version]
- Eaton, B.; Millar, R. Predicting gravel bed river response to environmental change: The strengths and limitations of a regime-based approach. Earth Surf. Process. Landforms 2017, 42, 994–1008. [Google Scholar] [CrossRef]
- Schumm, S.A. Meander wavelength of alluvial rivers. Science 1967, 157, 1549–1550. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.P. River meanders and channel size. J. Hydrol. 1986, 88, 147–164. [Google Scholar] [CrossRef]
- Maddock, T. Indeterminate Hydraulics of Alluvial Channels. J. Hydraul. Div. 1970, 96, 2309–2323. [Google Scholar] [CrossRef]
- Doyle, M.W.; Shields, D.; Boyd, K.F.; Skidmore, P.B.; Dominick, D. Channel-Forming Discharge Selection in River Restoration Design. J. Hydraul. Eng. 2007, 133, 831–837. [Google Scholar] [CrossRef]
- Brown, L.R.; Bauer, M.L. Effects of hydrologic infrastructure on flow regimes of california’s central valley rivers: Implications for fish populationsy. River Res. Appl. 2010, 26, 751–765. [Google Scholar] [CrossRef]
- Zeug, S.C.; Sellheim, K.; Watry, C.; Wikert, J.D.; Merz, J. Response of juvenile Chinook salmon to managed flow: Lessons learned from a population at the southern extent of their range in North America. Fish. Manag. Ecol. 2014, 21, 155–168. [Google Scholar] [CrossRef]
- Brown, L.R.; Ford, T. Effects of flow on the fish communities of a regulated California River: Implications for managing native fishes. River Res. Appl. 2002, 18, 331–342. [Google Scholar] [CrossRef]
- Merz, J.E.; Delaney, D.G.; Setka, J.D.; Workman, M.L. Seasonal Rearing Habitat in a Large Mediterranean-Climate River: Management Implications at the Southern Extent of Pacific Salmon (Oncorhynchus spp.). River Res. Appl. 2016, 32, 1220–1231. [Google Scholar] [CrossRef]
- Sommer, T.; Harrell, B.; Nobriga, M.; Brown, R.; Moyle, P.; Kimmerer, W.; Schemel, L. California’s Yolo Bypass: Evidence that flood control Can Be compatible with fisheries, wetlands, wildlife, and agriculture. Fisheries 2001, 6, 6–16. [Google Scholar] [CrossRef]
- Fargione, J.E.; Bassett, S.; Boucher, T.; Bridgham, S.D.; Conant, R.T.; Cook-Patton, S.C.; Ellis, P.W.; Falcucci, A.; Fourqurean, J.W.; Gopalakrishna, T.; et al. Natural climate solutions for the United States. Sci. Adv. 2018, 4, eaat1869. [Google Scholar] [CrossRef] [Green Version]
- Church, M. Bed Material Transport and The Morphology Of Alluvial River Channels. Annu. Rev. Earth Planet. Sci. 2006, 24, 325–354. [Google Scholar] [CrossRef]
- Riebe, C.S.; Sklar, L.S.; Overstreet, B.T.; Wooster, J.K. Optimal reproduction in salmon spawning substrates linked to grain size and fish length. Water Resour. Res. 2014, 50, 898–918. [Google Scholar] [CrossRef] [Green Version]
- Kondolf, G.M.; Wolman, M.G. The sizes of salmonid spawning gravels. Water Resour. Res. 1993, 29, 2275–2285. [Google Scholar] [CrossRef]
- Fuller, W.B.; Thompson, S.E. The laws of proportioning concrete. Trans. Am. Soc. Civ. Eng 1906, 59, 67–143. [Google Scholar] [CrossRef]
- Leopold, L.B.; Wolman, M.G. River Channel Patterns: Braided, Meandering, and Straight; Govt. Print. Off.: Washington, DC, USA, 1957; Volume 282-B.U.S. [Google Scholar]
- Williams, G.P.; Wolman, M.G. Downstream Effects of Dams On Alluvial Rivers; Government Printing Office: Washington, DC, USA, 1984. [Google Scholar]
- Dade, W.B.; Renshaw, C.E.; Magilligan, F.J. Sediment transport constraints on river response to regulation. Geomorphology 2011, 126, 245–251. [Google Scholar] [CrossRef]
- Blench, T.; Erb, B. La Théorie Du Régime Appliquée À L’analyse Des Résultats Expérimentaux Concernant Les Transports De Fond. La Houille Blanche 1957, 43, 132–157. [Google Scholar] [CrossRef] [Green Version]
- Eaton, B.C.; Church, M.; Millar, R.G. Rational regime model of alluvial channel morphology and response. Earth Surf. Process. Landforms 2004, 29, 511–529. [Google Scholar] [CrossRef]
- Hey, R.D.; Thorne, C.R. Stable Channels with Mobile Gravel Beds. J. Hydraul. Eng. 1986, 112, 671–689. [Google Scholar] [CrossRef]
- Davidson, S.K.; Hey, R.D. Regime Equations for Natural Meandering. J. Hydraul. Eng. 2011, 137, 894–910. [Google Scholar] [CrossRef]
- Griffiths, G.A. Stable-channel design in gravel-bed rivers. J. Hydrol. 1981, 52, 291–305. [Google Scholar] [CrossRef]
- Hoover Mackin, J. Concept of the graded river. Bull. Geol. Soc. Am. 1948, 59, 463–512. [Google Scholar] [CrossRef] [Green Version]
- Davies, T.R.H.; Sutherland, A.J. Extremal hypotheses for river behavior. Water Resour. Res. 1983, 19, 141–148. [Google Scholar] [CrossRef]
- Huang, H.Q.; Nanson, G.C. Hydraulic geometry and maximum flow efficiency as products of the principle of least action. Earth Surf. Process. Landforms 2000, 25, 1–16. [Google Scholar] [CrossRef]
- Huang, H.Q.; Chang, H.H.; Nanson, G.C. Minimum energy as the general form of critical flow and maximum flow efficiency and for explaining variations in river channel pattern. Water Resour. Res. 2004, 40, W04502. [Google Scholar] [CrossRef]
- Yang, C.T. Formation of Riffles and Pools. Water Resour. Res. 1971, 7, 1567–1574. [Google Scholar] [CrossRef]
- Copeland, R.R. Application of channel stability methods—Case studies; US Army Corps of Engineers, Waterways Experiment Station: Alexandria, VA, USA, 1994. [Google Scholar]
- Stroth, T.R.; Bledsoe, B.P.; Nelson, P.A. Full spectrum analytical channel design with the capacity/supply ratio (CSR). Water 2017, 9, 271. [Google Scholar] [CrossRef] [Green Version]
- Joshi, I.; Dai, W.; Bilal, A.; Upreti, A.R.; He, Z. Evaluation and comparison of extremal hypothesis-based regime methods. Water 2018, 10, 271. [Google Scholar] [CrossRef] [Green Version]
- Kaless, G.; Mao, L.; Lenzi, M.A. Regime theories in gravel-bed rivers: Models, controlling variables, and applications in disturbed Italian rivers. Hydrol. Process. 2014, 28, 2348–2360. [Google Scholar] [CrossRef] [Green Version]
- Buffington, J.M.; Montgomery, D.R. A systematic analysis of eight decades of incipient motion studies, with special reference to gravel-bedded rivers. Water Resour. Res. 1997, 33, 1993–2029. [Google Scholar] [CrossRef] [Green Version]
- Parker, G. Self-formed straight rivers with equilibrium banks and mobile bed. Part 2. The gravel river. J. Fluid Mech. 1978, 89, 127–146. [Google Scholar] [CrossRef]
- Phillips, C.B.; Jerolmack, D.J. Bankfull Transport Capacity and the Threshold of Motion in Coarse-Grained Rivers. Water Resour. Res. 2019, 55, 11316–11330. [Google Scholar] [CrossRef]
- Lamb, M.P.; Dietrich, W.E.; Venditti, J.G. Is the critical shields stress for incipient sediment motion dependent on channel-bed slope? J. Geophys. Res. Earth Surf. 2008, 113, F02008. [Google Scholar] [CrossRef] [Green Version]
- Hickin, E.J. Vegetation and River Channel Dynamics. Can. Geogr./Le Géographe Can. 1984, 28, 111–126. [Google Scholar] [CrossRef]
- Phillips, R.T.J.; Robert, A. Hydrologic control of waveforms on small meandering rivers. Earth Surf. Process. Landforms 2007, 32, 1533–1546. [Google Scholar] [CrossRef]
- Parker, G.; Wilcock, P.R.; Paola, C.; Dietrich, W.E.; Pitlick, J. Physical basis for quasi-universal relations describing bankfull hydraulic geometry of single-thread gravel bed rivers. J. Geophys. Res. Earth Surf. 2007, 112, F04005. [Google Scholar] [CrossRef]
- Millar, R.G. Theoretical regime equations for mobile gravel-bed rivers with stable banks. Geomorphology 2005, 64, 207–220. [Google Scholar] [CrossRef]
- Copeland, R.R.; Mccomas, D.N.; Thorne, C.R.; Soar, P.J.; Jonas, M.M.; Fripp, J.B. Hydraulic Design of Stream Restoration Projects; Coastal and Hydraulics Laboratory: Washington, DC, USA, 2001. [Google Scholar]
- Wilcock, P.R.; Crowe, J.C. Surface-based transport model for mixed-size sediment. J. Hydraul. Eng. 2003, 129, 120–128. [Google Scholar] [CrossRef]
- Eaton, B.C.; Millar, R.G.; Davidson, S. Channel patterns: Braided, anabranching, and single-thread. Geomorphology 2010, 120, 353–364. [Google Scholar] [CrossRef]
- Kleinhans, M.G. Sorting out river channel patterns. Prog. Phys. Geogr. 2010, 34, 287–326. [Google Scholar] [CrossRef]
- Wyrick, J.R.; Klingeman, P.C. Proposed fluvial island classification scheme and its use for river restoration. River Res. Appl. 2011, 27, 814–825. [Google Scholar] [CrossRef]
- Hintz, W.D.; Porreca, A.P.; Garvey, J.E.; Phelps, Q.E.; Tripp, S.J.; Hrabik, R.A.; Herzog, D.P. Abiotic Attributes Surrounding Alluvial Islands Generate Critical Fish Habitat. River Res. Appl. 2015, 31, 1218–1226. [Google Scholar] [CrossRef]
- Entwistle, N.; Heritage, G.; Milan, D. Ecohydraulic modelling of anabranching rivers. River Res. Appl. 2019, 35, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Hundey, E.J.; Ashmore, P.E. Length scale of braided river morphology. Water Resour. Res. 2009, 45, W08409–W08447. [Google Scholar] [CrossRef]
- Kleinhans, M.G.; van den Berg, J.H. River channel and bar patterns explained and predicted by an empirical and a physics-based method. Earth Surf. Process. Landforms 2011, 36, 721–738. [Google Scholar] [CrossRef]
- Parker, G. On the cause and characteristic scales of meandering and braiding in rivers. J. Fluid Mech. 1976, 76, 457–480. [Google Scholar] [CrossRef]
- Bledsoe, B.P.; Watson, C.C. Logistic analysis of channel pattern thresholds: Meandering, braiding, and incising. Geomorphology 2001, 38, 281–300. [Google Scholar] [CrossRef]
- Struiksma, N.; Olesen, K.W.; Flokstra, C.; de Vriend, H.J. Bed deformation in curved alluvial channels. Delft Neth. Delft Hydraul. LAB. Mar. 1985, 23, 57–79. [Google Scholar] [CrossRef]
- Colombini, M.; Seminara, G.; Tubino, M. Finite-amplitude alternate bars. J. Fluid Mech. 1987, 181, 213–232. [Google Scholar] [CrossRef]
- Repetto, R.; Tubino, M. Topographic expressions of bars in channels with variable width. Phys. Chem. Earth Part B Hydrol. Ocean. Atmos. 2001, 26, 71–76. [Google Scholar] [CrossRef]
- Nelson, J.M. The initial instability and finite-amplitude stability of alternate bars in straight channels. Earth Sci. Rev. 1990, 29, 97–115. [Google Scholar] [CrossRef]
- Wilkinson, S.N.; Rutherfurd, I.D.; Keller, R.J. An experimental test of whether bar instability contributes to the formation, periodicity and maintenance of pool-riffle sequences. Earth Surf. Process. Landforms 2008, 33, 1742–1756. [Google Scholar] [CrossRef]
- Furbish, D.J.; Thorne, S.D.; Byrd, T.C.; Warburton, J.; Cudney, J.J.; Handel, R.W. Irregular bed forms in steep, rough channels: 2. Field observations. Water Resour. Res. 1998, 34, 3649–3659. [Google Scholar] [CrossRef]
- Ferguson, R.I. Meander irregularity and wavelength estimation. J. Hydrol. 1975, 26, 315–333. [Google Scholar] [CrossRef]
- Ackers, P.; Charlton, F.G. Meander geometry arising from varying flows. J. Hydrol. 1970, 11, 230–252. [Google Scholar] [CrossRef]
- Hey, R.D. Geometry of river meanders. Nature 1976, 262, 482–484. [Google Scholar] [CrossRef]
- Soar, P.J.; Thorne, C.R. Channel Restoration Design for Meandering Rivers; Coastal and Hydraulics Laboratory: Washington, DC, USA, 2001. [Google Scholar]
- Ikeda, S. Prediction of alternate bar wavelength and height. J. Hydraul. Eng. 1984, 110, 371–386. [Google Scholar] [CrossRef]
- Church, M.; Rice, S.P. Form and growth of bars in a wandering gravel-bed river. Earth Surf. Process. Landforms 2009, 34, 1422–1432. [Google Scholar] [CrossRef]
- Meshkova, L.V.; Carling, P.A. Discrimination of alluvial and mixed bedrock-alluvial multichannel river networks. Earth Surf. Process. Landforms 2013, 38, 1299–1316. [Google Scholar] [CrossRef]
- Liu, X.; Huang, H.Q.; Nanson, G.C. The morphometric variation of islands in the middle and lower Yangtze River: A variational analytical explanation. Geomorphology 2016, 261, 273–281. [Google Scholar] [CrossRef]
- Burge, L.M. Stability, morphology and surface grain size patterns of channel bifurcation in gravel-cobble bedded anabranching rivers. Earth Surf. Process. Landforms 2006, 31, 1211–1226. [Google Scholar] [CrossRef]
- Bolla Pittaluga, M.; Coco, G.; Kleinhans, M.G. A unified framework for stability of channel bifurcations in gravel and sand fluvial systems. Geophys. Res. Lett. 2015, 42, 7521–7536. [Google Scholar] [CrossRef] [Green Version]
- Van Denderen, R.P.; Schielen, R.M.J.; Blom, A.; Hulscher, S.J.M.H.; Kleinhans, M.G. Morphodynamic assessment of side channel systems using a simple one-dimensional bifurcation model and a comparison with aerial images. Earth Surf. Process. Landforms 2017, 43, 1169–1182. [Google Scholar] [CrossRef] [Green Version]
- Geist, D.R.; Dauble, D.D. Redd site selection and spawning habitat use by fall chinook salmon: The importance of geomorphic features in large rivers. Environ. Manag. 1998, 22, 655–669. [Google Scholar] [CrossRef]
- Brown, R.A.R.A.; Pasternack, G.B.G.B. Bed and width oscillations form coherent patterns in a partially confined, regulated gravel-cobble-bedded river adjusting to anthropogenic disturbances. Earth Surf. Dyn. 2017, 5, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Duffin, J.; Carmichael, R.A.; Yager, E.M.; Benjankar, R.; Tonina, D. Detecting multi-scale riverine topographic variability and its influence on Chinook salmon habitat selection. Earth Surf. Process. Landforms 2021, 46, 1026–1040. [Google Scholar] [CrossRef]
- Dauble, D.D.; Geist, D.R. Comparison of mainstem spawning habitats for two populations of fall chinook salmon in the Columbia River basin. Regul. Rivers Res. Manag. 2000, 16, 345–361. [Google Scholar] [CrossRef]
- Shallin Busch, D.; Sheer, M.; Burnett, K.; Mcelhany, P.; Cooney, T. Landscape-level model to predict spawning habitat for lower Columbia river fall Chinook salmon (oncorhynchus tshawytscha). River Res. Appl. 2013, 29, 297–312. [Google Scholar] [CrossRef]
- Moir, H.J.; Pasternack, G.B. Relationships between mesoscale morphological units, stream hydraulics and Chinook salmon (Oncorhynchus tshawytscha) spawning habitat on the Lower Yuba River, California. Geomorphology 2008, 100, 527–548. [Google Scholar] [CrossRef]
- Keller, E.A.; Melhorn, W.N. Rhythmic spacing and origin of pools and riffles. Bull. Geol. Soc. Am. 1978, 89, 723–730. [Google Scholar] [CrossRef]
- Harvey, A.M. Some aspects of the relations between channel characteristics and riffle spacing in meandering. Am. J. Sci. 1975, 275, 470–478. [Google Scholar] [CrossRef]
- Thompson, D.M. Pool-Riffle. In Treatise on Geomorphology; Academic Press: Waltham, MA, USA, 2013; Volume 9, pp. 364–378. ISBN 9780080885223. [Google Scholar]
- Carling, P.A.; Orr, H.G. Morphology of Riffle Pool Sequences in the River Severn, England. Earth Surf. Process. Landforms 2000, 4, 369–384. [Google Scholar] [CrossRef]
- Yalin, M.S. River Mechanics; Peragamon Press: New York, NY, USA, 1971; ISBN 0080401902. [Google Scholar]
- MacVicar, B.J.; Obach, L.; Best, J.L. Large-Scale Coherent Flow Structures in Alluvial Pools. In Coherent Flow Structures at Earth’s Surface; John Wiley & Sons, Ltd.: New York, NY, USA, 2013; pp. 243–259. ISBN 9781118527221. [Google Scholar]
- Gregory, K.J.; Gurnell, A.M.; Hill, C.T.; Tooth, S. Stability of the pool-riffle sequence in changing river channels. Regul. Rivers Res. Manag. 1994, 9, 35–43. [Google Scholar] [CrossRef]
- Montgomery, D.R.; Buffington, J.M.; Smith, R.D.; Schmidt, K.M.; Pess, G. Pool Spacing in Forest Channels. Water Resour. Res. 1995, 31, 1097–1105. [Google Scholar] [CrossRef]
- Thompson, D.M. Random controls on semi-rhythmic spacing of pools and riffles in constriction-dominated rivers. Earth Surf. Process. Landforms 2001, 26, 1195–1212. [Google Scholar] [CrossRef]
- Thompson, D.M.; McCarrick, C.R. A flume experiment on the effect of constriction shape on the formation of forced pools. Hydrol. Earth Syst. Sci. 2010, 14, 1321–1330. [Google Scholar] [CrossRef] [Green Version]
- Thompson, D.M.; Hoffman, K.S. Equilibrium pool dimensions and sediment-sorting patterns in coarse-grained, New England channels. Geomorphology 2001, 38, 301–316. [Google Scholar] [CrossRef]
- Thompson, D.M.; Nelson, J.M.; Wohl, E.E. Interactions between pool geometry and hydraulics. Water Resour. Res. 1998, 34, 3673–3681. [Google Scholar] [CrossRef] [Green Version]
- Wohl, E.; Legleiter, C.J. Controls on pool characteristics along a resistant-boundary channel. J. Geol. 2003, 111, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Wyrick, J.R.; Pasternack, G.B. Geospatial organization of fluvial landforms in a gravel–cobble river: Beyond the riffle–pool couplet. Geomorphology 2014, 213, 48–65. [Google Scholar] [CrossRef] [Green Version]
- Grant, G.E.; Swanson, F.J.; Wolman, M.G. Pattern and origin of stepped-bed morphology in high-gradient streams, Western Cascades, Oregon. Bull. Geol. Soc. Am. 1990, 102, 340–352. [Google Scholar] [CrossRef]
- Wohl, E.E.; Vincent, K.R.; Merritts, D.J. Pool and riffle characteristics in relation to channel gradient. Geomorphology 1993, 6, 99–110. [Google Scholar] [CrossRef]
- Caamaño, D.; Goodwin, P.; Buffington, J.M.; Liou, J.C.P.; Daley-Laursen, S. Unifying criterion for the velocity reversal hypothesis in gravel-bed rivers. J. Hydraul. Eng. 2009, 135, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Melville, B.W. Local scour at bridge abutments. J. Hydraul. Eng. 1992, 118, 615–631. [Google Scholar] [CrossRef]
- Sawyer, A.M.; Pasternack, G.B.; Merz, J.E.; Escobar, M.; Senter, A.E. Construction constraints for geomorphic-unit rehabilitation on regulated gravel-bed rivers. River Res. Appl. 2009, 25, 416–437. [Google Scholar] [CrossRef]
- Resop, J.P.; Hession, W.C.; Wynn-Thompson, T. Quantifying the parameter uncertainty in the cross-sectional dimensions for a stream restoration design of a gravel-bed stream. J. Soil Water Conserv. 2014, 69, 306–315. [Google Scholar] [CrossRef]
- Bjornn, T.; Bjornn, T.; Reiser, D.; Reiser, D. Habitat requirements of salmonids in streams. Am. Fish. Soc. Spec. Publ. 1991, 19, 83–138. [Google Scholar]
- Brown, R.A.; Pasternack, G.B. Comparison of methods for analysing salmon habitat rehabilitation designs for regulated rivers. River Res. Appl. 2009, 25, 745–772. [Google Scholar] [CrossRef]
- Montgomery, D.R.; Beamer, E.M.; Pess, G.R.; Quinn, T.P. Channel type and salmonid spawning distribution and abundance. Can. J. Fish. Aquat. Sci. 1999, 56, 377–387. [Google Scholar] [CrossRef]
- Beechie, T.J.; Liermann, M.; Beamer, E.M.; Henderson, R. A Classification of Habitat Types in a Large River and Their Use by Juvenile Salmonids. Trans. Am. Fish. Soc. 2005, 134, 717–729. [Google Scholar] [CrossRef]
- Williams, P.B.; Andrews, E.; Opperman, J.J.; Bozkurt, S.; Moyle, P.B. Quantifying Activated Floodplains on a Lowland Regulated River: Its Application to Floodplain Restoration in the Sacramento Valley. San Fr. Estuary Watershed Sci. 2009, 7. [Google Scholar] [CrossRef] [Green Version]
- Hickey, J.T.; Huff, R.; Dunn, C.N. Using habitat to quantify ecological effects of restoration and water management alternatives. Environ. Model. Softw. 2015, 70, 16–31. [Google Scholar] [CrossRef]
- Chanson, H. Fundamentals of open channel flows. In Environmental Hydraulics for Open Channel Flows; Elsevier: Amsterdam, The Netherlands, 2004; pp. 11–34. [Google Scholar]
- Newbury, R.; Bates, D.; Alex, K.L. Restoring habitat hydraulics with constructed riffles. Geophys. Monogr. Ser. 2011, 194, 353–366. [Google Scholar] [CrossRef]
- California Department of Fish and Game Lower Mokelumne River Fisheries Management Plan; California Department of Fish and Game: Sacramento, CA, USA, 1991.
- Raleigh, R.; Miller, W. Habitat Suitability Index Models and Instream Flow Suitability Curves: Chinook Salmon; University of California Libraries: Washington, DC, USA, 1986. [Google Scholar]
- Bell, M.C. Fisheries Handbook of Engineering Requirements and Biological Criteria; University of Michigan Library: Seattle, WA, USA, 1986. [Google Scholar]
- Weeber, M.A.; Giannico, G.R.; Jacobs, S.E. Effects of Redd Superimposition by Introduced Kokanee on the Spawning Success of Native Bull Trout. N. Am. J. Fish. Manag. 2010, 30, 47–54. [Google Scholar] [CrossRef]
- Chapman, D.W.; Weitkamp, D.E.; Welsh, T.L.; Dell, M.B.; Schadt, T.H. Effects of River Flow on the Distribution of Chinook Salmon Redds. Trans. Am. Fish. Soc. 1986, 115, 537–547. [Google Scholar] [CrossRef]
- Grant, J.W.A.; Kramer, D.L. Territory size as a predictor of the upper limit to population density of juvenile salmonids in streams. Can. J. Fish. Aquat. Sci. 1990, 47, 1724–1737. [Google Scholar] [CrossRef]
- Brown, R.A.; Pasternack, G.B.; Wallender, W.W. Synthetic river valleys: Creating prescribed topography for form-process inquiry and river rehabilitation design. Geomorphology 2014, 214, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Bartz, K.K.; Lagueux, K.M.; Scheuerell, M.D.; Beechie, T.; Haas, A.D.; Ruckelshaus, M.H. Translating restoration scenarios into habitat conditions: An initial step in evaluating recovery strategies for Chinook salmon (Oncorhynchus tshawytscha). Can. J. Fish. Aquat. Sci. 2006, 63, 1578–1595. [Google Scholar] [CrossRef]
- Beechie, T.J.; Pess, G.R.; Imaki, H.; Martin, A.; Alvarez, J.; Goodman, D.H. Comparison of potential increases in juvenile salmonid rearing habitat capacity among alternative restoration scenarios, Trinity River, California. Restor. Ecol. 2015, 23, 75–84. [Google Scholar] [CrossRef]
- Brown, R.A.; Pasternack, G.B. Hydrologic and topographic variability modulate channel change in mountain rivers. J. Hydrol. 2014, 510, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Richards, K.S. Channel width and the riffle-pool sequence. Bull. Geol. Soc. Am. 1976, 87, 883–890. [Google Scholar] [CrossRef]
- Brew, A.K.; Morgan, J.A.; Nelson, P.A. Bankfull width controls on riffle-pool morphology under conditions of increased sediment supply: Field observations during the Elwha River dam removal project. In Proceedings of the Proceedings of the 3rd Joint Federal Interagency Conference on Sedimentation and Hydrologic Modeling, Reno, NV, USA, 19–23 April 2015. [Google Scholar]
- White, J.Q.; Pasternack, G.B.; Moir, H.J. Valley width variation influences riffle-pool location and persistence on a rapidly incising gravel-bed river. Geomorphology 2010, 121, 206–221. [Google Scholar] [CrossRef] [Green Version]
- Pasternack, G.B.; Baig, D.; Weber, M.D.; Brown, R.A. Hierarchically nested river landform sequences. Part 2: Bankfull channel morphodynamics governed by valley nesting structure. Earth Surf. Process. Landforms 2018, 43, 2519–2532. [Google Scholar] [CrossRef] [Green Version]
- Lisle, T.E. Stabilization of a gravel channel by large streamside obstructions and bedrock bends. Jacoby Creek, northwestern California. Geol. Soc. Am. Bull. 1986, 97, 999–1011. [Google Scholar] [CrossRef]
- Rhoads, B.L.; Welford, M.R. Initiation of river meandering. Prog. Phys. Geogr. 1991, 15, 127–156. [Google Scholar] [CrossRef]
- Luchi, R.; Zolezzi, G.; Tubino, M. Modelling mid-channel bars in meandering channels. Earth Surf. Process. Landforms 2010, 35, 902–917. [Google Scholar] [CrossRef]
- Schwartz, J.S.; Neff, K.J.; Dworak, F.E.; Woockman, R.R. Restoring riffle-pool structure in an incised, straightened urban stream channel using an ecohydraulic modeling approach. Ecol. Eng. 2015, 78, 112–126. [Google Scholar] [CrossRef] [Green Version]
- Wheaton, J.M.; Pasternack, G.B.; Merz, J.E. Use of habitat heterogeneity in salmonid spawning habitat rehabilitation design. In Proceedings of the Fifth International Symposium on Ecohydraulics: Aquatic Habitats: Analysis and Restoration, IAHR-AIRH, Madrid, Spain, 12–17 September; 2004; pp. 791–796. [Google Scholar]
- Pasternack, G.B.; Wang, C.L.; Merz, J.E. Application of a 2D hydrodynamic model to design of reach-scale spawning gravel replenishment on the Mokelumne River, California. River Res. Appl. 2004, 20, 205–225. [Google Scholar] [CrossRef]
- Kui, L.; Stella, J.C.; Shafroth, P.B.; House, P.K.; Wilcox, A.C. The long-term legacy of geomorphic and riparian vegetation feedbacks on the dammed Bill Williams River, Arizona, USA. Ecohydrology 2017, 10, e1839. [Google Scholar] [CrossRef]
- Gordon, E.; Meentemeyer, R.K. Effects of dam operation and land use on stream channel morphology and riparian vegetation. Geomorphology 2006, 82, 412–429. [Google Scholar] [CrossRef]
- Stella, J.C.; Vick, J.C.; Orr, B.K. Riparian vegetation dynamics on the Merced River. In Proceedings of the California Riparian Systems: Processes and Floodplains Management, Ecology, and Restoration (2001 Riparian Habitat and Floodplains Conference Proceedings), Sacramento, CA, USA, 12–15 March 2003; pp. 302–314. [Google Scholar]
- Bair, J.H. Limiting riparian hardwood encroachment along the Trinity River, California. In Proceedings of the Engineering Approaches to Ecosystem Restoration, Denver, CO, USA, 22–27 March 1998. [Google Scholar]
- Wohl, E.; Bledsoe, B.P.; Jacobson, R.B.; Poff, N.L.; Rathburn, S.L.; Walters, D.M.; Wilcox, A.C. The natural sediment regime in rivers: Broadening the foundation for ecosystem management. Bioscience 2015, 65, 358–371. [Google Scholar] [CrossRef] [Green Version]
- DeVries, P. Riverine salmonid egg burial depths: Review of published data and implications for scour studies. Can. J. Fish. Aquat. Sci. 1997, 54, 1685–1698. [Google Scholar] [CrossRef]
- Pasternack, G.B.; Bounrisavong, M.K.; Parikh, K.K. Backwater control on riffle-pool hydraulics, fish habitat quality, and sediment transport regime in gravel-bed rivers. J. Hydrol. 2008, 357, 125–139. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, R.B.; Galat, D.L. Design of a naturalized flow regime-an example from the Lower Missouri River, USA. Ecohydrology 2008, 1, 81–104. [Google Scholar] [CrossRef] [Green Version]
- Merz, J.E.; Pasternack, G.B.; Wheaton, J.M. Sediment budget for salmonid spawning habitat rehabilitation in a regulated river. Geomorphology 2006, 76, 207–228. [Google Scholar] [CrossRef]
- Moore, H.E.; Rutherfurd, I.D. Lack of maintenance is a major challenge for stream restoration projects. River Res. Appl. 2017, 33, 1387–1399. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Width (m) | Depth (m) | W/D | Assumptions |
|---|---|---|---|---|
| UBCRM (Eaton and Millar 2017) | 28.7 | 1.40 | 20.4 | μ’ = 0.98/ Type I vegetation |
| Parker et al., 2007 | 29.5 | 1.26 | 23.5 | r = 1.5 |
| Equations (2) and (3) | 31.7 | 1.26 | 25.2 | r = 1.5 |
| Hey and Thorne, 1983 | 32.6 | 1.43 | 22.8 | Type I vegetation |
| Average | 30.6 | 1.34 | 23.0 | |
| Standard deviation | 1.59 | 0.08 | 1.73 | |
| Coefficient of variation | 0.05 | 0.06 | 0.08 |
| Mesohabitat Unit | Example(s) | GCS Relationship | Citations |
|---|---|---|---|
| Riffle | Riffles located at point bar crossover | Low curvature, wider than average, higher than average bed elevation | [128,171,172,173] |
| Pool | Pools located adjacent to river bends or adjacent to gravel bars and forcing elements | High curvature, narrower than average, lower than average bed elevation | [146,147,149,174] |
| Lateral gravel bar | Gravel bars located on inward side of bend or downstream of an obstruction | High curvature, wider than average at bankfull and greater flow | [128,144,175,176] |
| River island | Islands located in channel expansions and/or after channel bends | Wider than average, low curvature at bankfull or greater flow | [94,177] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brown, R.A. Hydrogeomorphic Scaling and Ecohydraulics for Designing Rescaled Channel and Floodplain Geometry in Regulated Gravel–Cobble Bed Rivers for Pacific Salmon Habitat. Water 2022, 14, 670. https://doi.org/10.3390/w14040670
Brown RA. Hydrogeomorphic Scaling and Ecohydraulics for Designing Rescaled Channel and Floodplain Geometry in Regulated Gravel–Cobble Bed Rivers for Pacific Salmon Habitat. Water. 2022; 14(4):670. https://doi.org/10.3390/w14040670
Chicago/Turabian StyleBrown, Rocko A. 2022. "Hydrogeomorphic Scaling and Ecohydraulics for Designing Rescaled Channel and Floodplain Geometry in Regulated Gravel–Cobble Bed Rivers for Pacific Salmon Habitat" Water 14, no. 4: 670. https://doi.org/10.3390/w14040670