
Litter Mass Loss of the Invasive Rhus typhina L. and Native Koelreuteria paniculata Laxm. Trees Alters Soil N-Fixing Bacterial Community Composition under Different N Forms
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Determination of the Decomposition Variables
2.3. Determination of Soil Physicochemical Properties and Soil Enzyme Activities
2.4. Determination of NFB
2.5. Statistical Analysis
3. Results
3.1. Differences in the Decomposition Variables
3.2. Differences in Soil Physicochemical Properties and Soil Enzyme Activities
3.3. Differences in NFB Alpha Diversity
3.4. The Contribution Intensity of Soil Physicochemical Properties, Soil Enzyme Activities, and NFB Alpha Diversity to the Litter Mass Loss of the Two Trees
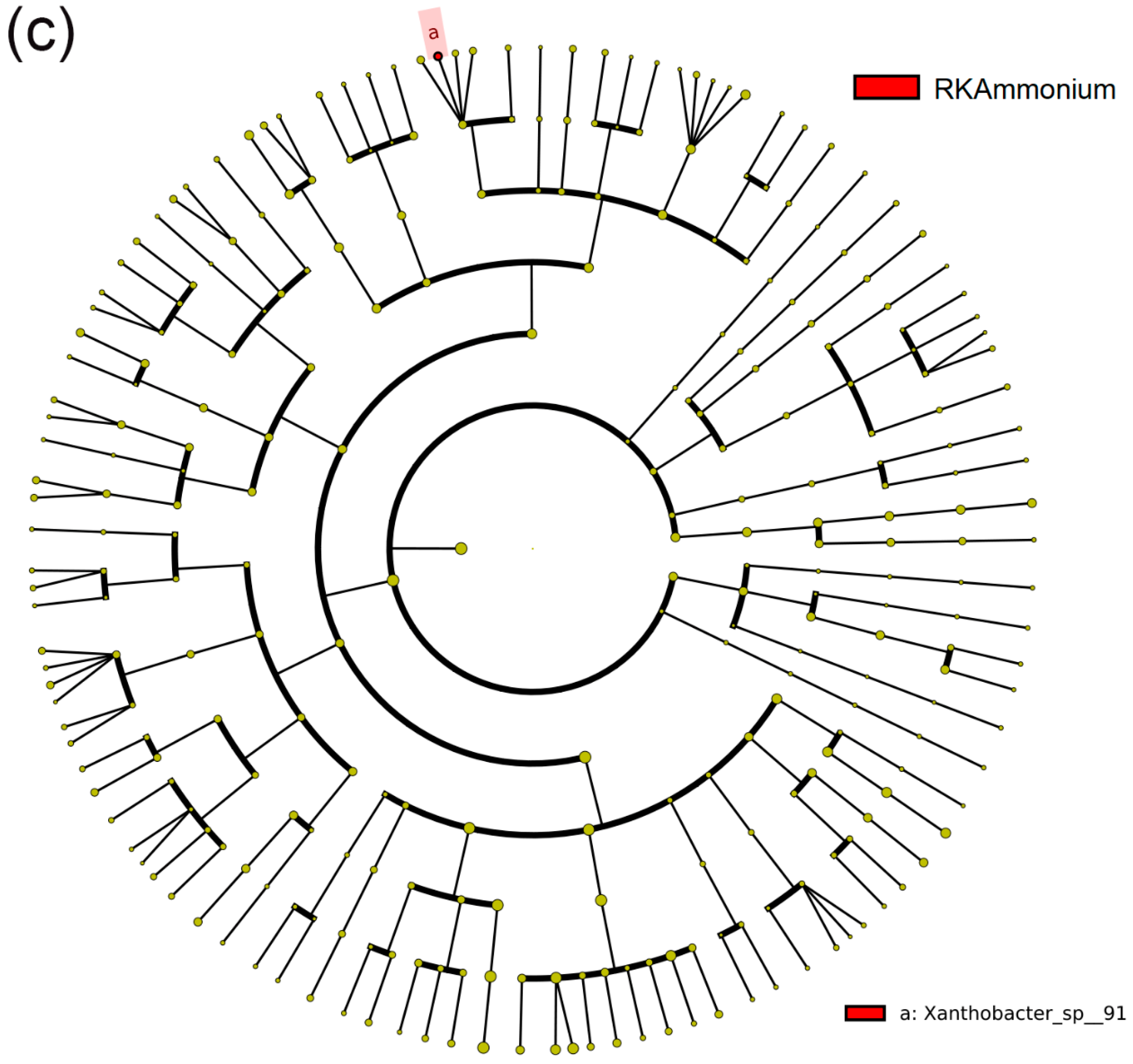
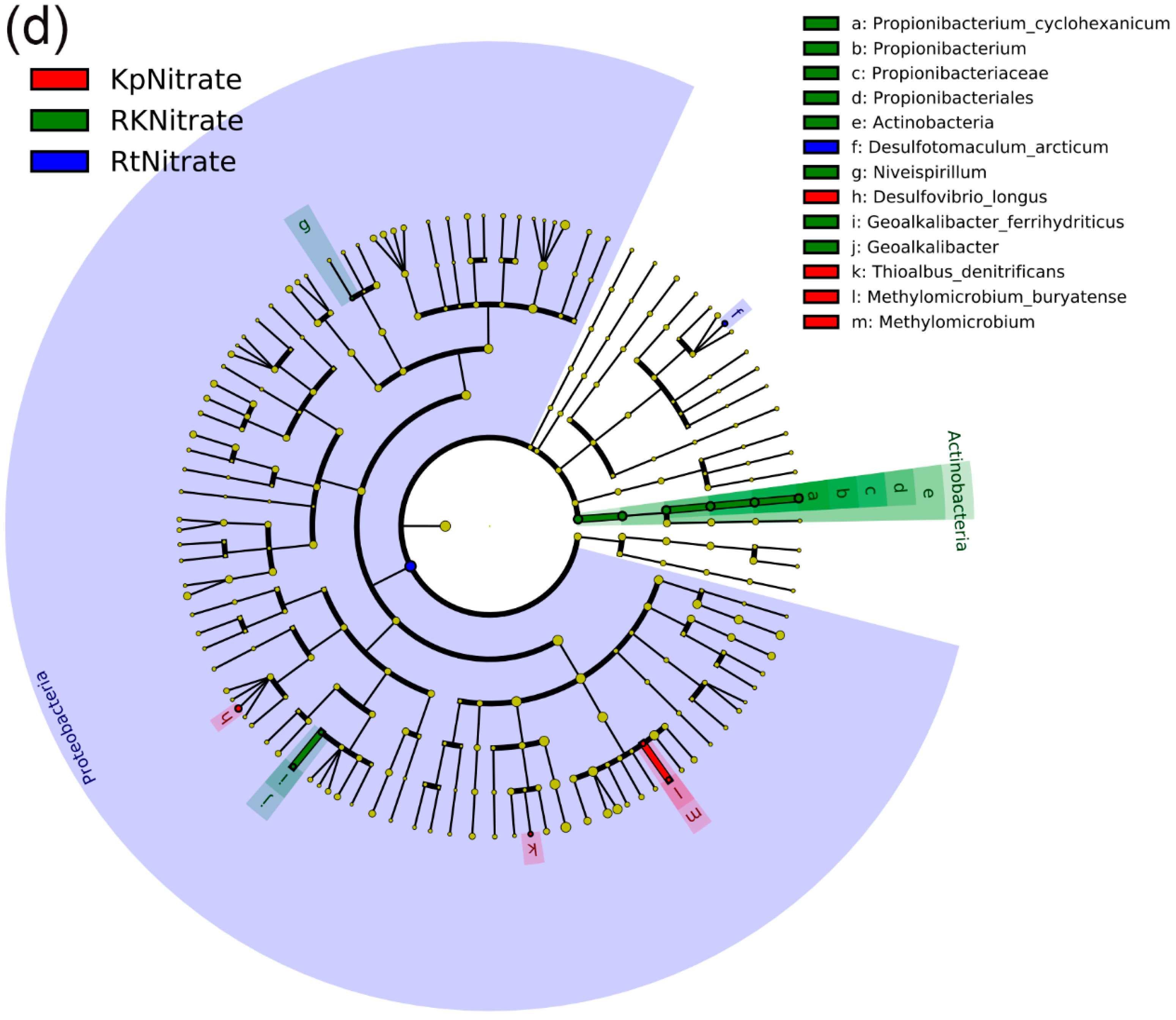
3.5. Differences in the NFB Community Structure among Different Treatments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Q.W.; Zhang, X.Y.; Liang, J.F.; Gao, J.Q.; Xu, X.L.; Yu, F.H. High nitrogen uptake and utilization contribute to the dominance of invasive Spartina alterniflora over native Phragmites australis. Biol. Fertil. Soils 2021, 57, 1007–1013. [Google Scholar] [CrossRef]
- Parepa, M.; Fischer, M.; Bossdorf, O. Environmental variability promotes plant invasion. Nat. Commun. 2013, 4, 1604. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.L.; Lei, Y.B.; Wang, R.F.; Callaway, R.M.; Alfonso, V.B.; Inderjit; Li, Y.P.; Zheng, Y.L. Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant. Proc. Natl. Acad. Sci. USA 2009, 106, 1853–1856. [Google Scholar] [CrossRef] [PubMed]
- Mantoani, M.C.; Gonzalez, A.B.; Sancho, L.G.; Osborne, B.A. Growth, phenology and N-utilization by invasive populations of Gunnera tinctoria. J. Plant Ecol. 2020, 13, 589–600. [Google Scholar] [CrossRef]
- Davidson, A.M.; Jennions, M.; Nicotra, A.B. Do invasive species show higher phenotypic plasticity than native species and, if so, is it adaptive? A meta-analysis. Ecol. Lett. 2011, 14, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Kamutando, C.N.; Vikram, S.; Kamgan-Nkuekam, G.; Makhalanyane, T.P.; Greve, M.; Le Roux, J.J.; Richardson, D.M.; Cowan, D.; Valverde, A. Soil nutritional status and biogeography influence rhizosphere microbial communities associated with the invasive tree Acacia dealbata. Sci. Rep. 2017, 7, 6472. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.W.; Yang, M.Z.; Chen, Y.J.; Chen, L.M.; Zhang, D.Z.; Mei, L.; Shi, Y.T.; Zhang, H.B. Changes in non-symbiotic nitrogen-fixing bacteria inhabiting rhizosphere soils of an invasive plant Ageratina adenophora. Appl. Soil Ecol. 2012, 54, 32–38. [Google Scholar] [CrossRef]
- Li, J.; He, J.Z.; Liu, M.; Yan, Z.Q.; Xu, X.L.; Kuzyakov, Y. Invasive plant competitivity is mediated by nitrogen use strategies and rhizosphere microbiome. Soil Biol. Biochem. 2024, 192, 109361. [Google Scholar] [CrossRef]
- Reed, S.C.; Townsend, A.R.; Cleveland, C.C.; Nemergut, D.R. Microbial community shifts influence patterns in tropical forest nitrogen fixation. Oecologia 2010, 164, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Chinnadurai, C.; Gopalaswamy, G.; Balachandar, D. Long term effects of nutrient management regimes on abundance of bacterial genes and soil biochemical processes for fertility sustainability in a semi-arid tropical Alfisol. Geoderma 2014, 232, 563–572. [Google Scholar] [CrossRef]
- Huhe; Borjigin, S.; Buhebaoyin; Wu, Y.P.; Li, M.Q.; Cheng, Y.X. Microbial nitrogen-cycle gene abundance in soil of cropland abandoned for different periods. PLoS ONE 2016, 11, e0154697. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Schloter-Hai, B.; Schloter, M.; van Elsas, J.D.; Salles, J.F. Temporal dynamics of abundance and composition of nitrogen-fixing communities across agricultural soils. PLoS ONE 2013, 8, e74500. [Google Scholar]
- Lindsay, E.A.; Colloff, M.J.; Gibb, N.L.; Wakelin, S.A. The abundance of microbial functional genes in grassy woodlands is influenced more by soil nutrient enrichment than by recent weed invasion or livestock exclusion. Appl. Environ. Microbiol. 2010, 76, 5547–5555. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Zhou, J.W.; Liu, J.; Jiang, K.; Du, D.L. Responses of soil N-fixing bacteria communities to Amaranthus retroflexus invasion under different forms of N deposition. Agric. Ecosyst. Environ. 2017, 247, 329–336. [Google Scholar] [CrossRef]
- Ren, B.H.; Meng, M.; Yu, J.X.; Ma, X.W.; Li, D.Y.; Li, J.H.; Yang, J.Y.; Bai, L.; Feng, Y.L. Invasion by Cenchrus spinifex changes the soil microbial community structure in a sandy grassland ecosystem. Heliyon 2023, 9, e20860. [Google Scholar] [CrossRef] [PubMed]
- Lau, J.A.; Suwa, T. The changing nature of plant-microbe interactions during a biological invasion. Biol. Invasions 2016, 18, 3527–3534. [Google Scholar] [CrossRef]
- Wang, Z.J.; Li, X.; Wang, J.H.; Qi, S.S.; Dai, Z.C.; Du, D.L. Effect of nitrogen-fixing bacteria on resource investment of the root system in an invasive clonal plant under low nutritional environment. Flora 2022, 297, 152166. [Google Scholar] [CrossRef]
- Cuassolo, F.; Diaz Villanueva, V.; Modenutti, B. Litter decomposition of the invasive Potentilla anserina in an invaded and non-invaded freshwater environment of North Patagonia. Biol. Invasions 2020, 22, 1055–1065. [Google Scholar] [CrossRef]
- Liao, C.Z.; Peng, R.H.; Luo, Y.Q.; Zhou, X.H.; Wu, X.W.; Fang, C.M.; Chen, J.K.; Li, B. Altered ecosystem carbon and nitrogen cycles by plant invasion: A meta-analysis. New Phytol. 2008, 177, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Arif, M.; Ding, D.D.; Li, J.J.; He, X.R.; Li, C.X. Invasive plants and species richness impact litter decomposition in Riparian Zones. Front. Plant Sci. 2022, 13, 955656. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.L.; Zhong, S.S.; Yu, Y.L.; Li, Y.; Li, C.; Xu, Z.Y.; Liu, J.; Wang, C.Y.; Du, D.L. Heavy metal contamination alters the co-decomposition of leaves of the invasive tree Rhus typhina L. and the native tree Koelreuteria paniculata Laxm. Plants 2023, 12, 2523. [Google Scholar] [CrossRef] [PubMed]
- Marchante, E.; Marchante, H.; Freitas, H.; Kjøller, A.; Struwe, S. Decomposition of an N-fixing invasive plant compared with a native species: Consequences for ecosystem. Appl. Soil Ecol. 2019, 138, 19–31. [Google Scholar] [CrossRef]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, L.; Liu, X.J.; Li, W.Q.; Lu, S.H.; Zheng, L.X.; Bai, Z.C.; Cai, G.Y.; Zhang, F.S. Atmospheric organic nitrogen deposition in China. Atmos. Environ. 2012, 49, 422. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, L.; Guo, X.; Xu, Y.; Luo, L. Atmospheric wet and dry deposition of dissolved inorganic nitrogen to the South China Sea. Sci. China-Earth Sci. 2020, 63, 1339–1352. [Google Scholar] [CrossRef]
- Cornell, S.E. Atmospheric nitrogen deposition: Revisiting the question of the importance of the organic component. Environ. Pollut. 2011, 159, 2214–2222. [Google Scholar] [CrossRef] [PubMed]
- Sinsabaugh, R.L.; Carreiro, M.M.; Repert, D.A. Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss. Biogeochemistry 2002, 60, 1–24. [Google Scholar] [CrossRef]
- Li, C.; Li, Y.; Zhong, S.S.; Xu, Z.L.; Xu, Z.Y.; Zhu, M.W.; Wei, Y.Q.; Wang, C.Y.; Du, D.L. Do the leaves of multiple invasive plants decompose more easily than a native plant’s under nitrogen deposition with different forms? Nitrogen 2024, 5, 202–218. [Google Scholar] [CrossRef]
- Saiz, E.; Sgouridis, F.; Drijfhout, F.P.; Peichl, M.; Nilsson, M.B.; Ullah, S. Chronic atmospheric reactive nitrogen deposition suppresses biological nitrogen fixation in Peatlands. Environ. Sci. Technol. 2021, 55, 1310–1318. [Google Scholar] [CrossRef] [PubMed]
- Berthrong, S.T.; Yeager, C.M.; Gallegos-Graves, L.; Steven, B.; Eichorst, S.A.; Jackson, R.B.; Kuske, C.R. Nitrogen fertilization has a stronger effect on soil nitrogen-fixing bacterial communities than elevated atmospheric CO2. Appl. Environ. Microbiol. 2014, 80, 3103–3112. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.L.; Chen, J.Z.; Wang, D.; Deng, M.M.; Wu, M.Y.; Tong, B.L.; Liu, J.M. Simulated atmospheric nitrogen deposition inhibited the leaf litter decomposition of Cinnamomum migao H. W. Li in Southwest China. Sci. Rep. 2021, 11, 1748. [Google Scholar] [CrossRef]
- Whalen, E.D.; Smith, R.G.; Grandy, A.S.; Frey, S.D. Manganese limitation as a mechanism for reduced decomposition in soils under atmospheric nitrogen deposition. Soil Biol. Biochem. 2018, 127, 252–263. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Suseela, V. Nitrogen availability modulates the impacts of plant invasion on the chemical composition of soil organic matter. Soil Biol. Biochem. 2021, 156, 108195. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Jiang, C.D.; Zhang, J.Z.; Zhang, H.J.; Shi, L. Ecophysiological evaluation of the potential invasiveness of Rhus typhina in its non-native habitats. Tree Physiol. 2009, 29, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Kossah, R.; Zhang, H.; Chen, W. Antimicrobial and antioxidant activities of Chinese sumac (Rhus typhina L.) fruit extract. Food Control 2011, 22, 128–132. [Google Scholar] [CrossRef]
- Tan, X.F.; Guo, X.; Guo, W.H.; Liu, S.N.; Du, N. Invasive Rhus typhina invests more in height growth and traits associated with light acquisition than do native and non-invasive alien shrub species. Trees 2018, 32, 1103–1112. [Google Scholar] [CrossRef]
- Fu, Y.D.; Xu, W.; Wen, Z.; Han, M.J.; Sun, J.H.; Tang, A.H.; Liu, X. Enhanced atmospheric nitrogen deposition at a rural site in northwest China from 2011 to 2018. Atmos. Res. 2020, 245, 105071. [Google Scholar] [CrossRef]
- Luo, X.S.; Liu, X.J.; Pan, Y.P.; Wen, Z.; Xu, W.; Zhang, L.; Kou, C.L.; Lv, J.L.; Goulding, K. Atmospheric reactive nitrogen concentration and deposition trends from 2011 to 2018 at an urban site in north China. Atmos. Environ. 2020, 224, 117298. [Google Scholar] [CrossRef]
- Zhu, J.X.; Chen, Z.; Wang, Q.F.; Xu, L.; He, N.P.; Jia, Y.L.; Zhang, Q.Y.; Yu, G.R. Potential transition in the effects of atmospheric nitrogen deposition in China. Environ. Pollut. 2020, 258, 113739. [Google Scholar] [CrossRef] [PubMed]
- National Bureau of Statistics. Zhenjiang Statistical Yearbook 2022; China Statistics Press: Beijing, China, 2022.
- Cornell, S.E.; Jickells, T.D.; Cape, J.N.; Rowland, A.P.; Duce, R.A. Organic nitrogen deposition on land and coastal environments: A review of methods and data. Atmos. Environ. 2003, 37, 2173–2191. [Google Scholar] [CrossRef]
- Xu, Z.L.; Zhong, S.S.; Yu, Y.L.; Wang, Y.Y.; Cheng, H.Y.; Du, D.L.; Wang, C.Y. Rhus typhina L. triggered greater allelopathic effects than Koelreuteria paniculata Laxm under ammonium fertilization. Sci. Hortic. 2023, 309, 111703. [Google Scholar] [CrossRef]
- Voříšková, J.; Baldrian, P. Fungal community on decomposing leaf litter undergoes rapid successional changes. ISME J. 2013, 7, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wei, Z.; Huangfu, C.; Chen, X.; Yang, D. Litter mixture dominated by leaf litter of the invasive species, Flaveria bidentis, accelerates decomposition and favors nitrogen release. J. Plant Res. 2017, 130, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Hoorens, B.; Aerts, R.; Stroetenga, M. Does initial litter chemistry explain litter mixture effects on decomposition. Oecologia 2003, 137, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.L.; Scullion, J.; Worgan, H.; Gwynn-Jones, D. Litter of the invasive shrub Rhododendron ponticum (Ericaceae) modifies the decomposition rate of native UK woodland litter. Ecol. Indic. 2019, 107, 105597. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Zhong, S.S.; Xu, Z.L.; Yu, Y.L.; Liu, J.; Wang, Y.Y.; Guo, E.; Wang, C.Y. Rhus typhina decreased soil nitrogen contents and peroxidase activity following the addition of nitrogen. Int. J. Environ. Sci. Technol. 2023, 111, 17–22. [Google Scholar] [CrossRef]
- Yu, Y.L.; Cheng, H.Y.; Wang, C.Y.; Du, D.L. Heavy drought reduces the decomposition rate of the mixed litters of two composite invasive alien plants. J. Plant Ecol. 2023, 16, rtac047. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ceccanti, B.; Cervelli, S.; Matarese, E. Extraction of phosphatase, urease, proteases, organic-carbon, and nitrogen from soil. Soil Sci. Soc. Am. J. 1980, 44, 1011–1016. [Google Scholar] [CrossRef]
- Guan, S.Y. Soil Enzyme and Its Research Methods; Agricultural Press: Beijing, China, 1986. [Google Scholar]
- Zhang, J.E. Experimental Methods and Techniques Commonly Used in Ecology; Chemical Industry Press: Beijing, China, 2006. [Google Scholar]
- Perucci, P.; Casucci, C.; Dumontet, S. An improved method to evaluate the o-diphenol oxidase activity of soil. Soil Biol. Biochem. 2000, 32, 1927–1933. [Google Scholar] [CrossRef]
- Gaby, J.C.; Buckley, D.H. A comprehensive aligned nifH gene database: A multipurpose tool for studies of nitrogen-fixing bacteria. Database 2014, 2014, bau001. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Peng, M.J.; Li, Y.G. Phylogenetic diversity of nitrogen-fixing bacteria and the nifH gene from mangrove rhizosphere soil. Can. J. Microbiol. 2012, 58, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Hu, B.; Zhu, J.B.; Shen, S.J.; Yu, G.Q. nifH promoter activity is regulated by DNA supercoiling in Sinorhizobium meliloti. Acta Biochim. Biophys. Sin. 2005, 37, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Poly, F.; Ranjard, L.; Nazaret, S.; Gourbière, F.; Monrozier, L.J. Comparison of nifH gene pools in soils and soil microenvironments with contrasting properties. Appl. Environ. Microbiol. 2001, 67, 2255–2262. [Google Scholar] [CrossRef] [PubMed]
- Norris, M.D.; Blair, J.M.; Johnson, L.C. Land cover change in eastern Kansas: Litter dynamics of closed-canopy eastern redcedar forests in tallgrass prairie. Can. J. Bot. Rev. Can. Bot. 2001, 79, 214–222. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Ecosystem consequences of biological invasions. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 59–80. [Google Scholar] [CrossRef]
- Vila, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarosik, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pysek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.D.; Lockaby, B.G.; Brantley, E.F. Influence of Chinese Privet (Ligustrum sinense) on decomposition and nutrient availability in riparian forests. Invasive Plant Sci. Manag. 2011, 4, 437–447. [Google Scholar] [CrossRef]
- Schuster, M.J.; Dukes, J.S. Non-additive effects of invasive tree litter shift seasonal N release: A potential invasion feedback. Oikos 2014, 123, 1101–1111. [Google Scholar] [CrossRef]
- Gartner, T.B.; Cardon, Z.G. Decomposition dynamics in mixed species leaf litter. Oikos 2004, 104, 230–246. [Google Scholar] [CrossRef]
- Torres, N.; Herrera, I.; Fajardo, L.; Bustamante, R.O. Meta-analysis of the impact of plant invasions on soil microbial communities. BMC Ecol. Evol. 2021, 21, 172. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Staver, A.C. Enhanced activity of soil nutrient-releasing enzymes after plant invasion: A meta-analysis. Ecology 2019, 100, e02830. [Google Scholar] [CrossRef] [PubMed]
- Keet, J.H.; Ellis, A.G.; Hui, C.; Novoa, A.; Le Roux, J.J. Impacts of invasive Australian acacias on soil bacterial community composition, microbial enzymatic activities, and nutrient availability in fynbos soils. Microb. Ecol. 2021, 82, 704–721. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Yan, W.B.; Quan, G.M.; Zhang, J.E.; Liang, K.M. Soil microbial carbon utilization, enzyme activities and nutrient availability responses to Bidens pilosa and a non-invasive congener under different irradiances. Sci. Rep. 2017, 7, 11309. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.D.; Nielsen, C.; Hughes, R.F. Elevated enzyme activities in soils under the invasive nitrogen-fixing tree Falcataria moluccana. Soil Biol. Biochem. 2006, 38, 1537–1544. [Google Scholar] [CrossRef]
- Xu, H.W.; Liu, Q.; Wang, S.Y.; Yang, G.S.; Xue, S. A global meta-analysis of the impacts of exotic plant species invasion on plant diversity and soil properties. Sci. Total Environ. 2022, 810, 152286. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yuan, J.H.; Wang, Y.; Butterly, C.R.; Tong, D.L.; Zhou, B.; Li, X.Z.; Zhang, H.B. Effects of exotic Spartina alterniflora invasion on soil phosphorus and carbon pools and associated soil microbial community composition in coastal wetlands. ACS Omega 2021, 6, 5730–5738. [Google Scholar] [CrossRef] [PubMed]
- Stefanowicz, A.M.; Stanek, M.; Nobis, M.; Zubek, S. Species–specific effects of plant invasions on activity, biomass, and composition of soil microbial communities. Biol. Fertil. Soils 2016, 52, 841–852. [Google Scholar] [CrossRef]
- Zubek, S.; Majewska, M.L.; Błaszkowski, J.; Stefanowicz, A.M.; Nobis, M.; Kapusta, P. Invasive plants affect arbuscularmycorrhizal fungi abundance and species richness as well as the performance of native plants grown in invaded soils. Biol. Fertil. Soils 2016, 52, 879–893. [Google Scholar] [CrossRef]
- Sun, X.; Gao, C.; Guo, L. Changes in soil microbial community and enzyme activity along an exotic plant Eupatorium adenophorum invasion in a Chinese secondary forest. Chin. Sci. Bull. 2013, 58, 4101–4108. [Google Scholar] [CrossRef]
- Scharfy, D.; Güsewell, S.; Gessner, M.O.; Venterink, H.O. Invasion of Solidago gigantea in contrasting experimental plant communities: Effects on soil microbes, nutrients and plant–soil feedbacks. J. Ecol. 2010, 98, 1379–1388. [Google Scholar] [CrossRef]
- Stanek, M.; Zubek, S.; Stefanowicz, A.M. Differences in phenolics produced by invasive Quercus rubra and native plant communities induced changes in soil microbial properties and enzymatic activity. For. Ecol. Manag. 2021, 482, 118901. [Google Scholar] [CrossRef]
- Yu, Y.L.; Xu, Z.L.; Zhong, S.S.; Cheng, H.Y.; Guo, E.R.; Wang, C.Y. The co-invasion of the three Asteraceae invasive plants can synergistically increase soil phenol oxidase activity. Biol. Bull. 2023, 50, 467–473. [Google Scholar]
- Li, Y.; Wang, C.M.; Gao, S.J.; Wang, P.; Qiu, J.C.; Shang, S.H. Impacts of simulated nitrogen deposition on soil enzyme activity in a northern temperate forest ecosystem depend on the form and level of added nitrogen. Eur. J. Soil Biol. 2021, 103, 103287. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Wang, S.; Wei, M.; Yu, Y.L.; Wang, C.Y. Alien invasive plant Amaranthus spinosus mainly altered the community structure instead of the α diversity of soil N-fixing bacteria under drought. Acta Oecol. 2021, 113, 103788. [Google Scholar] [CrossRef]
- Wang, C.Y.; Jiang, K.; Zhou, J.W.; Wu, B.D. Solidago canadensis invasion affects soil N-fixing bacterial communities in heterogeneous landscapes in urban ecosystems in East China. Sci. Total Environ. 2018, 631–632, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.T.; Yang, C.J.; Huang, K.H.; Huang, J.C.; Lin, T.C. Changes of precipitation acidity related to sulfur and nitrogen deposition in forests across three continents in north hemisphere over last two decades. Sci. Total Environ. 2022, 806, 150552. [Google Scholar] [CrossRef] [PubMed]
- Matocha, C.J.; Grove, J.H.; Karathanasis, T.D.; Vandiviere, M. Changes in soil mineralogy due to nitrogen fertilization in an agroecosystem. Geoderma 2016, 263, 176–184. [Google Scholar] [CrossRef]
- Craine, J.M.; Morrow, C.; Fierer, N. Microbial nitrogen limitation increases decomposition. Ecology 2007, 88, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Thirukkumaran, C.M.; Parkinson, D. Microbial respiration, biomass, metabolic quotient and litter decomposition in a lodgepole pine forest floor amended with nitrogen and phosphorous fertilizers. Soil Biol. Biochem. 2000, 32, 59–66. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil pH | Soil Total N Content (g/kg) | Soil Urease Activity (μg/g soil/d) | Soil Protease Activity (μg/g soil/d) | Soil Polyphenol Oxidase Activity (μmol/g soil/h) | |
|---|---|---|---|---|---|
| Control | 6.54 ± 0.03 a–c | 8.31 ± 0.41 de | 76.67 ± 5.48 b | 486.78 ± 53.12 b–e | 79.91 ± 6.96 ab |
| Ammonium | 6.53 ± 0.07 a–c | 8.94 ± 0.41 de | 73.28 ± 2.64 b | 838.24 ± 66.48 a | 86.67 ± 7.22 a |
| Nitrate | 6.47 ± 0.03 a–d | 9.33 ± 0.44 de | 98.08 ± 15.08 b | 251.47 ± 13.20 e–h | 50.21 ± 4.05 b–d |
| Urea | 6.13 ± 0.09 de | 15.92 ± 0.37 c | 95.50 ± 25.05 b | 501.94 ± 41.44 b–e | 30.30 ± 2.79 d |
| MixN | 6.10 ± 0.06 e | 15.42 ± 0.50 c | 78.59 ± 21.97 b | 731.19 ± 76.38 ab | 31.79 ± 6.59 d |
| Rt | 6.67 ± 0.07 ab | 9.20 ± 0.33 de | 86.16 ± 8.89 b | 653.43 ± 48.04 a–c | 36.37 ± 1.85 cd |
| RtAmmonium | 6.33 ± 0.07 b–e | 9.22 ± 0.20 de | 85.36 ± 14.53 b | 133.31 ± 36.15 gh | 27.74 ± 2.24 d |
| RtNitrate | 6.50 ± 0.10 a–c | 9.40 ± 0.41 de | 79.08 ± 21.52 b | 553.44 ± 71.11 a–d | 32.86 ± 6.22 d |
| RtUrea | 6.40 ± 0.00 a–e | 9.38 ± 0.27 de | 82.78 ± 12.34 b | 89.882 ± 11.91 h | 45.06 ± 17.21 d |
| RtMixN | 6.533 ± 0.07 a–c | 9.39 ± 0.11 de | 79.56 ± 10.38 b | 430.23 ± 54.23 c–f | 44.74 ± 5.0 cd |
| Kp | 6.67 ± 0.07 ab | 10.06 ± 0.40 d | 87.45 ± 8.18 b | 112.10 ± 24.30 gh | 65.15 ± 2.61 a–c |
| KpAmmonium | 6.60 ± 0.00 a–c | 8.33 ± 0.52 de | 85.84 ± 8.14 b | 359.54 ± 23.23 d–h | 50.30 ± 1.86 b–d |
| KpNitrate | 6.27 ± 0.07 c–e | 9.31 ± 0.34 de | 207.76 ± 19.63 a | 456.49 ± 67.90 b–f | 50.14 ± 6.75 b–d |
| KpUrea | 6.43 ± 0.03 a–e | 9.25 ± 0.48 de | 86.32 ± 14.70 b | 248.44 ± 42.42 e–h | 45.62 ± 1.39 cd |
| KpMixN | 6.53 ± 0.03 a–c | 9.23 ± 0.04 de | 47.67 ± 7.91 b | 199.972 ± 71.46 f–h | 47.74 ± 1.53 cd |
| RK | 6.40 ± 0.06 a–e | 7.82 ± 0.62 e | 76.02 ± 10.19 b | 258.54 ± 47.60 e–h | 51.47 ± 6.31 b–d |
| RKAmmonium | 6.43 ± 0.09 a–e | 8.35 ± 0.44 de | 94.86 ± 9.13 b | 274.70 ± 46.49 d–h | 46.10 ± 7.40 cd |
| RKNitrate | 6.630.12 ab | 21.73 ± 0.23 a | 64.90 ± 14.08 b | 276.72 ± 86.13 d–h | 29.26 ± 2.016 d |
| RKUrea | 6.70 ± 0.06 a | 19.30 ± 0.53 b | 85.04 ± 5.32 b | 483.76 ± 55.60 b–f | 26.82 ± 2.58 d |
| RKMixN | 6.60 ± 0.10 a–c | 9.78 ± 0.52 de | 87.45 ± 7.86 b | 391.85 ± 51.37 c–g | 41.23 ± 2.93 cd |
| OTU’s Species Index | Simpson’s Dominance Index | Chao1′s Richness Index | ACE’s Richness Index | |
|---|---|---|---|---|
| Control | 941.67 ± 25.56 a | 0.81 ± 0.01 a–c | 1597.13 ± 52.36 a | 1591.61 ± 33.56 a–d |
| Ammonium | 897.33 ± 19.62 ab | 0.79 ± 0.00 a–c | 1489.10 ± 3.04 ab | 1534.02 ± 44.11 a–e |
| Nitrate | 941.00 ± 35.57 a | 0.83 ± 0.02 a–c | 1588.74 ± 62.94 a | 1619.08 ± 68.79 ab |
| Urea | 904.33 ± 22.81 ab | 0.85 ± 0.02 ab | 1583.79 ± 57.64 a | 1656.02 ± 38.10 a |
| MixN | 823.67 ± 21.17 a–c | 0.86 ± 0.02 a | 1435.62 ± 107.20 ab | 1487.79 ± 59.44 a–f |
| Rt | 819.67 ± 0.67 a–c | 0.81 ± 0.01 a–c | 1385.54 ± 37.03 ab | 1417.76 ± 32.54 b–f |
| RtAmmonium | 921.00 ± 20.31 ab | 0.79 ± 0.01 a–c | 1560.81 ± 22.94 ab | 1603.92 ± 17.34 a–c |
| RtNitrate | 895.00 ± 9.45 ab | 0.79 ± 0.01 a–c | 1486.11 ± 25.69 ab | 1530.61 ± 20.43 a–e |
| RtUrea | 755.33 ± 31.57 c | 0.81 ± 0.01 a–c | 1325.73 ± 23.99 ab | 1396.96 ± 35.28 c–f |
| RtMixN | 861.67 ± 37.24 a–c | 0.77 ± 0.00 a–c | 1370.22 ± 24.60 ab | 1444.76 ± 35.28 a–f |
| Kp | 856.33 ± 19.70 a–c | 0.75 ± 0.02 a–c | 1383.04 ± 73.72 ab | 1362.96 ± 54.94 ef |
| KpAmmonium | 849.00 ± 15.72 a–c | 0.74 ± 0.02 bc | 1385.36 ± 21.76 ab | 1375.59 ± 24.04 ef |
| KpNitrate | 834.67 ± 56.16 a–c | 0.73 ± 0.03 c | 1368.83 ± 130.51 ab | 1379.77 ± 72.14 d–f |
| KpUrea | 876.67 ± 13.25 a–c | 0.74 ± 0.02 a–c | 1430.61 ± 25.78 ab | 1428.08 ± 12.92 b–f |
| KpMixN | 844.67 ± 21.94 a–c | 0.79 ± 0.04 a–c | 1288.93 ± 55.25 b | 1341.60 ± 39.35 ef |
| RK | 821.67 ± 12.17 a–c | 0.76 ± 0.02 a–c | 1312.43 ± 32.68 ab | 1323.73 ± 26.17 ef |
| RKAmmonium | 875.00 ± 20.11 a–c | 0.78 ± 0.04 a–c | 1443.83 ± 34.18 ab | 1448.28 ± 29.61 a–f |
| RKNitrate | 806.67 ± 8.41 bc | 0.78 ± 0.03 a–c | 1317.81 ± 14.34 ab | 1306.66 ± 7.45 f |
| RKUrea | 795.00 ± 28.43 bc | 0.74 ± 0.03 bc | 1412.85 ± 40.51 ab | 1350.84 ± 38.09 ef |
| RKMixN | 832.00 ± 15.63 a–c | 0.73 ± 0.03 c | 1383.86 ± 44.21 ab | 1356.12 ± 25.61 ef |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Li, C.; Cheng, H.; Xu, Z.; Zhong, S.; Zhu, M.; Wei, Y.; Xu, Z.; Du, D.; Wang, C.; et al. Litter Mass Loss of the Invasive Rhus typhina L. and Native Koelreuteria paniculata Laxm. Trees Alters Soil N-Fixing Bacterial Community Composition under Different N Forms. Atmosphere 2024, 15, 424. https://doi.org/10.3390/atmos15040424
Li Y, Li C, Cheng H, Xu Z, Zhong S, Zhu M, Wei Y, Xu Z, Du D, Wang C, et al. Litter Mass Loss of the Invasive Rhus typhina L. and Native Koelreuteria paniculata Laxm. Trees Alters Soil N-Fixing Bacterial Community Composition under Different N Forms. Atmosphere. 2024; 15(4):424. https://doi.org/10.3390/atmos15040424
Chicago/Turabian StyleLi, Yue, Chuang Li, Huiyuan Cheng, Zhelun Xu, Shanshan Zhong, Mawei Zhu, Yuqing Wei, Zhongyi Xu, Daolin Du, Congyan Wang, and et al. 2024. "Litter Mass Loss of the Invasive Rhus typhina L. and Native Koelreuteria paniculata Laxm. Trees Alters Soil N-Fixing Bacterial Community Composition under Different N Forms" Atmosphere 15, no. 4: 424. https://doi.org/10.3390/atmos15040424