
Changes in the Fluorescence of Biological Particles Exposed to Environmental Conditions in the National Capitol Region
 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. CAGE System
2.2. Experimental Design
2.3. Preparation, Enumeration, and Generation of Bioaerosols
2.3.1. Btk Spores
2.3.2. MS2
2.4. Aerosol Generation
2.5. Aerosol Size, Concentration, and Fluorescence Measurement
2.6. Ambient Environmental Measurements
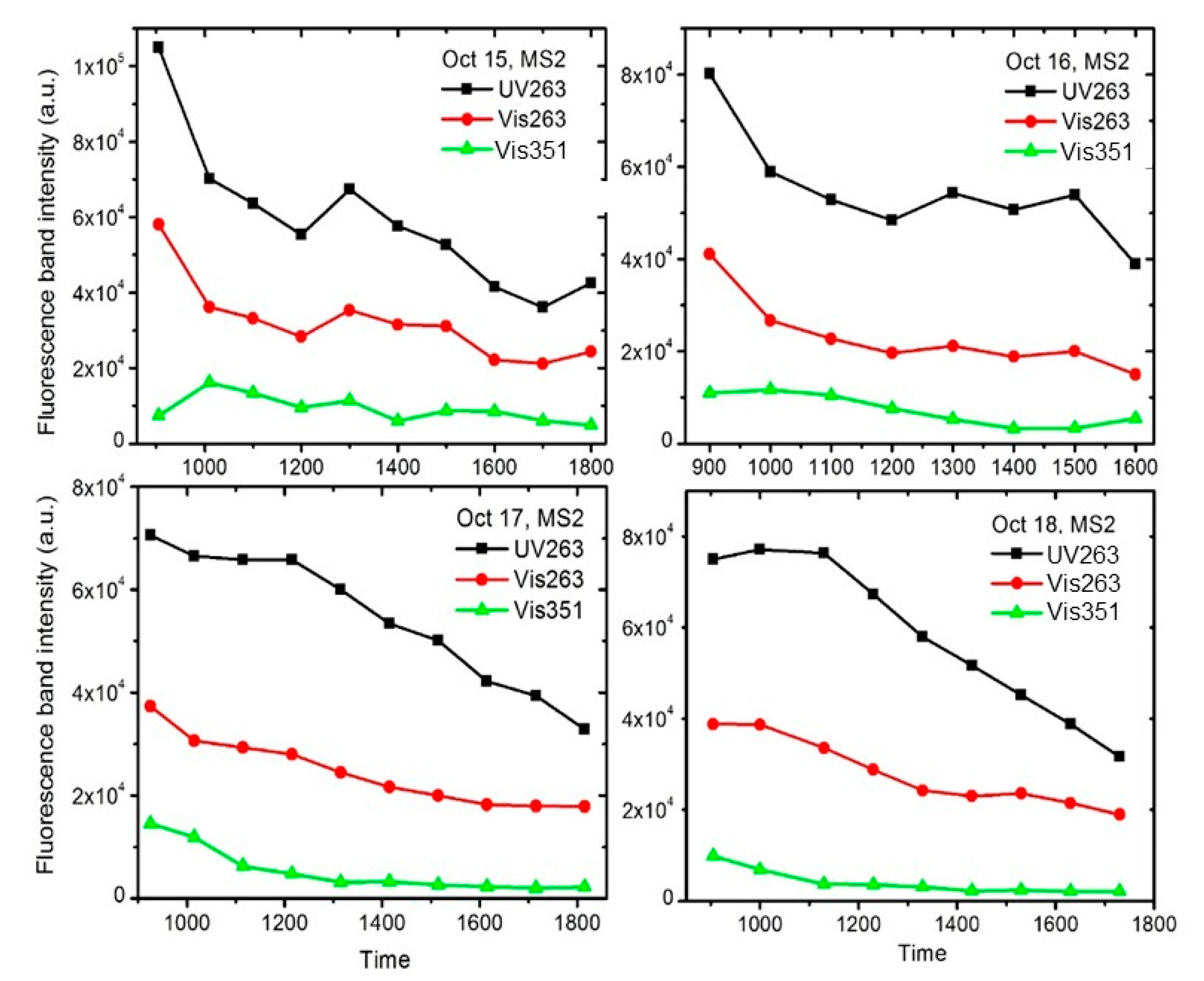
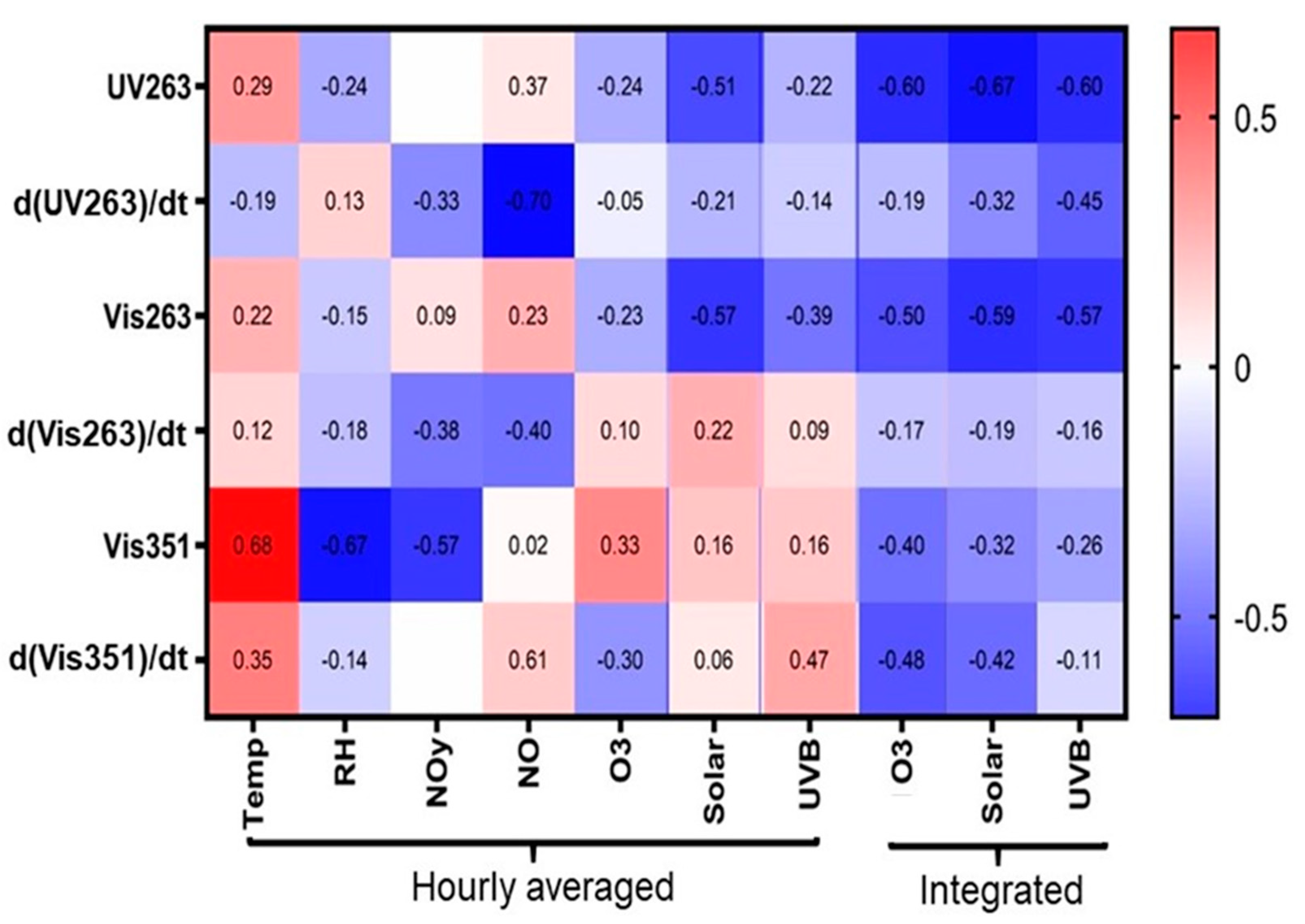
3. Results and Discussions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Möhler, O.; DeMott, P.J.; Vali, G.; Levin, Z. Microbiology and Atmospheric Processes: The Role of Biological Particles in Cloud Physics. Biogeosciences 2007, 4, 1059–1071. [Google Scholar] [CrossRef]
- Beedham, R.J.; Davies, C.H. The UK Biological-Warfare Program: Dual-Use Contributions to the Field of Aerobiology. Nonprolif. Rev. 2021, 27, 309–322. [Google Scholar] [CrossRef]
- Liu, D.; Mariman, R.; Gerlofs-Nijland, M.E.; Boere, J.F.; Folkerts, G.; Cassee, F.R.; Pinelli, E. Microbiome Composition of Airborne Particulate Matter from Livestock Farms and Their Effect on Innate Immune Receptors and Cells. Sci. Total Environ. 2019, 688, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Mazar, Y.; Cytryn, E.; Erel, Y.; Rudich, Y. Effect of Dust Storms on the Atmospheric Microbiome in the Eastern Mediterranean. Environ. Sci. Technol. 2016, 50, 4194–4202. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Lu, R.; Xie, Z.S.; Wang, J.L.; Fan, C.L.; Liu, P.X.; Li, Y.P. Concentration and Size Distribution Characteristics of Culturable Bioaerosols at Various Air Quality Levels During Fall and Winter in Xi’an, China. Huanjing Kexue/Environ. Sci. 2017, 38, 4494–4500. [Google Scholar] [CrossRef]
- Han, C.; Xie, M.; Qi, J.; Zhang, W.; Li, X.; Zhang, D. Size Distribution Characteristics of Culturable Bioaerosols in Relation to Air Quality Levels in Qingdao. Res. Environ. Sci. 2016, 29, 1264–1271. [Google Scholar] [CrossRef]
- Santarpia, J.L. Bioaerosols in the Environment: Populations, Measurement and Processes; The Royal Society of Chemistry: London, UK, 2016. [Google Scholar]
- Sharma, R.; Ranjan, R.; Kapardar, R.K.; Grover, A. “Unculturable” Bacterial Diversity: An Untapped Resource. Curr. Sci. 2005, 89, 72–77. [Google Scholar]
- Cox, C.S.; Hood, A.M.; Baxter, J. Method for Comparing Concentrations of the Open-Air Factor. Appl. Microbiol. 1973, 26, 640–642. [Google Scholar] [CrossRef]
- Hood, A.M. Open-Air Factors in Enclosed Systems. J. Hyg. 1974, 72, 53–60. [Google Scholar] [CrossRef]
- Hood, A.M. The Effect of Open-Air Factors on the Virulence and Viability of Airborne. Epidemiol. Infect. 2009, 137, 753–761. [Google Scholar] [CrossRef]
- Donaldson, A.I.; Ferris, N.P. The Survival of Foot-and-Mouth Disease Virus in Open Air Conditions. J. Hyg. 1975, 74, 409–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthony Cox, R.; Ammann, M.; Crowley, J.N.; Griffiths, P.T.; Herrmann, H.; Hoffmann, E.H.; Jenkin, M.E.; Faye McNeill, V.; Mellouki, A.; Penkett, C.J.; et al. Opinion: The Germicidal Effect of Ambient Air (Open-Air Factor) Revisited. Atmos. Chem. Phys. 2021, 21, 13011–13018. [Google Scholar]
- Pan, Y.-L.; Kalume, A.; Wang, C.; Santarpia, J. Atmospheric Aging Processes of Bioaerosols under Laboratory-Controlled Conditions: A Review. J. Aerosol Sci. 2021, 155, 105767. [Google Scholar] [CrossRef]
- Ratnesar-Shumate, S.; Pan, Y.-L.; Hill, S.C.; Kinahan, S.; Corson, E.; Eshbaugh, J.; Santarpia, J.L. Fluorescence Spectra and Biological Activity of Aerosolized Bacillus Spores and MS2 Bacteriophage Exposed to Ozone at Different Relative Humidities in a Rotating Drum. J. Quant. Spectrosc. Radiat. Transf. 2015, 153, 13–28. [Google Scholar] [CrossRef]
- Santarpia, J.L.; Pan, Y.-L.; Hill, S.C.; Baker, N.; Cottrell, B.; McKee, L.; Ratnesar-Shumate, S.; Pinnick, R.G. Changes in Fluorescence Spectra of Bioaerosols Exposed to Ozone in a Laboratory Reaction Chamber to Simulate Atmospheric Aging. Opt. Express 2012, 20, 29867–29881. [Google Scholar] [CrossRef]
- Pan, Y.-L.; Hill, S.C.; Pinnick, R.G.; Santarpia, J.L.; Baker, N.; Alvarez, B.; Ratnesar-Shumate, S.; Cottrell, B.; McKee, L. Fluorescence Spectra of Bioaerosols Exposed to Ozone in a Laboratory Reaction Chamber to Simulate Atmospheric Processing. In Proceedings of the SPIE—The International Society for Optical Engineering, Orlando, FL, USA, 25–29 April 2011; Volume 8018. [Google Scholar]
- Pan, Y.-L.; Santarpia, J.L.; Ratnesar-Shumate, S.; Corson, E.; Eshbaugh, J.; Hill, S.C.; Williamson, C.C.; Coleman, M.; Bare, C.; Kinahan, S. Effects of Ozone and Relative Humidity on Fluorescence Spectra of Octapeptide Bioaerosol Particles. J. Quant. Spectrosc. Radiat. Transf. 2014, 133, 538–550. [Google Scholar] [CrossRef]
- Kinahan, S.M.; Tezak, M.S.; Siegrist, C.M.; Lucero, G.; Servantes, B.L.; Santarpia, J.L.; Kalume, A.; Zhang, J.; Felton, M.; Williamson, C.C.; et al. Changes of Fluorescence Spectra and Viability from Aging Aerosolized E. coli Cells under Various Laboratory-Controlled Conditions in an Advanced Rotating Drum. Aerosol Sci. Technol. 2019, 53, 1261–1276. [Google Scholar] [CrossRef]
- Adhikari, A.; Reponen, T.; Grinshpun, S.A.; Martuzevicius, D.; Lemasters, G. Correlation of Ambient Inhalable Bioaerosols with Particulate Matter and Ozone: A Two-Year Study. Environ. Pollut. 2006, 140, 16–28. [Google Scholar] [CrossRef]
- Chi, M.C.; Li, C.S. Fluorochrome in Monitoring Atmospheric Bioaerosols and Correlations with Meteorological Factors and Air Pollutants. Aerosol Sci. Technol. 2007, 41, 672–678. [Google Scholar] [CrossRef]
- Kowalski, M.; Pastuszka, J.S. Effect of Ambient Air Temperature and Solar Radiation on Changes in Bacterial and Fungal Aerosols Concentrationin the Urban Environment. Ann. Agric. Environ. Med. 2018, 25, 259–261. [Google Scholar] [CrossRef]
- Santarpia, J.L.; Ratnesar-Shumate, S.; Gilberry, J.U.; Quizon, J.J. Relationship between Biologically Fluorescent Aerosol and Local Meteorological Conditions. Aerosol Sci. Technol. 2013, 47, 655–661. [Google Scholar] [CrossRef]
- Pecson, B.M.; Martin, L.V.; Kohn, T. Quantitative PCR for Determining the Infectivity of Bacteriophage MS2 upon Inactivation by Heat, UV-B Radiation, and Singlet Oxygen: Advantages and Limitations of an Enzymatic Treatment to Reduce False-Positive Results. Appl. Environ. Microbiol. 2009, 75, 5544–5554. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.H.; Lee, J.E. Variation in the fluorescence intensity of thermally-exposed bacterial bioaerosols. J. Aerosol Sci. 2013, 65, 101–110. [Google Scholar] [CrossRef]
- Schuit, M.; Ratnesar-Shumate, S.; Yolitz, J.; Williams, G.; Weaver, W.; Green, B.; Miller, D.; Krause, M.; Beck, K.; Wood, S.; et al. Airborne SARS-CoV-2 is Rapidly Inactivated by Simulated Sunlight. J. Infect. Dis. 2020, 222, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Verreault, D.; Marcoux-Voiselle, M.; Turgeon, N.; Moineau, S.; Duchaine, C. Resistance of Aerosolized Bacterial Viruses to Relative Humidity and Temperature. Appl. Environ. Microbiol. 2015, 81, 7305–7311. [Google Scholar] [CrossRef] [PubMed]
- Peccia, J.; Werth, H.M.; Miller, S.; Hernandez, M. Effects of Relative Humidity on the Ultraviolet Induced Inactivation of Airborne Bacteria. Aerosol Sci. Technol. 2001, 35, 728–740. [Google Scholar] [CrossRef]
- Kowalski, W.J.; Bahnfleth, W.P.; Whittam, T.S. Bactericidal Effects of High Airborne Ozone Concentrations on Escherichia coli and Staphylococcus aureus. Ozone Sci. Eng. 1998, 20, 205–221. [Google Scholar] [CrossRef]
- Zhang, F.; Xi, J.; Huang, J.-J.; Hu, H.-Y. Effect of inlet ozone concentration on the performance of a micro-bubble ozonation system for inactivation of Bacillus subtilis spores. Sep. Purif. Technol. 2013, 114, 126–133. [Google Scholar] [CrossRef]
- Clauss, M.; Linke, S.; Tautz, C.; Tautz, C.; Bromann, S. Development of a Novel Bioaerosol Chamber to Determine Survival Rates of Airborne Staphylococci. Atmosphere 2022, 13, 869. [Google Scholar] [CrossRef]
- Chen, L.; Bao, K.; Li, K.; Lv, B.; Bao, Z.; Lin, C.; Wu, X.; Zheng, C.; Gao, X.; Cen, K. Ozone and Secondary Organic Aerosol Formation of Toluene/NOx Irradiations under Complex Pollution Scenarios. Aerosol Air Qual. Res. 2017, 17, 1760–1771. [Google Scholar] [CrossRef]
- Park, J.-H.; Babar, Z.B.; Baek, S.J.; Kim, H.S.; Lim, H.-J. Effects of NOx on the molecular composition of secondary organic aerosol formed by the ozonolysis and photooxidation of α-pinene. Atmos. Environ. 2017, 166, 263–275. [Google Scholar] [CrossRef]
- Šantl-Temkiv, T.; Sikoparija, B.; Maki, T.; Carotenuto, F.; Amato, P.; Yao, M.; Morris, C.E.; Schnell, R.; Jaenicke, R.; Pöhlker, C.; et al. Bioaerosol field measurements: Challenges and perspectives in outdoor studies. Aerosol Sci. Technol. 2020, 54, 520–546. [Google Scholar] [CrossRef]
- Peng, J.; Hu, M.; Guo, S.; Du, Z.; Zheng, J.; Shang, D.; Levy Zamora, M.; Zeng, L.; Shao, M.; Wu, Y.-S.; et al. Markedly Enhanced Absorption and Direct Radiative Forcing of Black Carbon under Polluted Urban Environments. Proc. Natl. Acad. Sci. USA 2016, 113, 4266–4271. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, D.; Dommen, J.; Kalberer, M.; Prévôt, A.S.H.; Richter, R.; Sax, M.; Steinbacher, M.; Weingartner, E.; Baltensperger, U. Secondary Organic Aerosol Formation by Irradiation of 1,3,5-Trimethylbenzene–NOx–H2O in a New Reaction Chamber for Atmospheric Chemistry and Physics. Environ. Sci. Technol. 2005, 39, 2668–2678. [Google Scholar] [CrossRef]
- Goldberg, L.J.; Watkins, H.M.S.; Boerke, E.E.; Chatigny, M.A. The Use of a Rotating Drum for the Study of Aerosols over Extended Periods of Time. Am. J. Epidemiol. 1958, 68, 85–93. [Google Scholar] [CrossRef]
- Antonietti, C.G. Development of the Captive Aerosol Growth and Evolution Chamber System. Master’s Thesis, Texas A&M University, College Station, TX, USA, 2014. [Google Scholar]
- Haddrell, A.E.; Thomas, R.J. Aerobiology: Experimental Considerations, Observations, and Future Tools. Appl. Environ. Microbiol. 2017, 83, e00809-17. [Google Scholar] [CrossRef]
- Verreault, D.; Duchaine, C.; Marcoux-Voiselle, M.; Turgeon, N.; Roy, C.J. Design of an Environmentally Controlled Rotating Chamber for Bioaerosol Aging Studies. Inhal. Toxicol. 2014, 26, 554–558. [Google Scholar] [CrossRef]
- Krumins, V.; Son, E.K.; Mainelis, G.; Fennell, D.E. Retention of Inactivated Bioaerosols and Ethene in a Rotating Bioreactor Constructed for Bioaerosol Activity Studies. Clean Soil Air Water 2008, 36, 593–600. [Google Scholar] [CrossRef]
- Ratnesar-Shumate, S.; Wagner, M.L.; Kerechanin, C.; House, G.; Brinkley, K.M.; Bare, C.; Baker, N.A.; Quizon, R.; Quizon, J.; Proescher, A.; et al. Improved Method for the Evaluation of Real-Time Biological Aerosol Detection Technologies. Aerosol Sci. Technol. 2011, 45, 635–644. [Google Scholar] [CrossRef]
- Pan, Y.-L.; Hill, S.C.; Santarpia, J.L.; Brinkley, K.; Sickler, T.; Coleman, M.; Williamson, C.; Gurton, K.; Felton, M.; Pinnick, R.G.; et al. Spectrally-Resolved Fluorescence Cross Sections of Aerosolized Biological Live Agents and Simulants Using Five Excitation Wavelengths in a BSL-3 Laboratory. Opt. Express 2014, 22, 8165–8189. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santarpia, J.L.; Collins, D.R.; Ratnesar-Shumate, S.A.; Glen, C.C.; Sanchez, A.L.; Antonietti, C.G.; Taylor, J.; Taylor, N.F.; Bare, C.A.; Kinahan, S.M.; et al. Changes in the Fluorescence of Biological Particles Exposed to Environmental Conditions in the National Capitol Region. Atmosphere 2022, 13, 1358. https://doi.org/10.3390/atmos13091358
Santarpia JL, Collins DR, Ratnesar-Shumate SA, Glen CC, Sanchez AL, Antonietti CG, Taylor J, Taylor NF, Bare CA, Kinahan SM, et al. Changes in the Fluorescence of Biological Particles Exposed to Environmental Conditions in the National Capitol Region. Atmosphere. 2022; 13(9):1358. https://doi.org/10.3390/atmos13091358
Chicago/Turabian StyleSantarpia, Joshua L., Don R. Collins, Shanna A. Ratnesar-Shumate, Crystal C. Glen, Andres L. Sanchez, Carlos G. Antonietti, Jilliane Taylor, Nathan F. Taylor, Christopher A. Bare, Sean M. Kinahan, and et al. 2022. "Changes in the Fluorescence of Biological Particles Exposed to Environmental Conditions in the National Capitol Region" Atmosphere 13, no. 9: 1358. https://doi.org/10.3390/atmos13091358