
Effects of Climate Change on Forest Regeneration in Central Spain


1
Department of Biodiversity, Ecology and Evolution, Faculty of Biological Sciences, Universidad Complutense de Madrid, 28040 Madrid, Spain
2
Draba Ingeniería y Consultoría Medioambiental, 28200 San Lorenzo de El Escorial, Spain
Atmosphere 2022, 13(7), 1143; https://doi.org/10.3390/atmos13071143
Submission received: 23 June 2022
/
Revised: 14 July 2022
/
Accepted: 15 July 2022
/
Published: 18 July 2022
(This article belongs to the Special Issue Feature Papers in Atmosphere Science)
Abstract
:The Mediterranean climate has dry and hot summers, which is harsh for plants, especially seedlings. During the 1950s and 1960s, most reforestations carried out in Central Spain, a Mediterranean climate area, were successful, but in recent decades an increasing difficulty in forest regeneration has been observed, often attributed to increased summer drought. This study analyses changes in climatic parameters related to forest regeneration through statistical treatment of meteorological data series from the mid-twentieth century to the present. Simple and multiple regressions and ANOVAs were performed for five parameters, considering annual, summer and extended summer values. Rainfall reduction and prolongation of the summer drought period were not statistically significant. The change that better explains regeneration problems is the increase in temperature, especially in July and August, which was mostly significant between 2002 and 2021. Raising temperatures increase the vapor pressure deficit, exacerbating drought effects and plant mortality. Climate change scenarios point to an increase in temperatures until 2100; thus, the tipping point for natural regeneration of some species could be passed. The most affected species are those at their ecological limit. It is necessary to facilitate the adaptation of these forests to climate change, since their future will depend on the actions carried out today.
1. Introduction
Climate change is altering climatic parameters in an indisputable way, raising temperatures in the Mediterranean region. It has a more uncertain influence on precipitation, which is undoubtedly suffering greater irregularity, and probably, a reduction. This is one of the most significant threats for Mediterranean forests, which are suffering a decline due to the combined effect of warming and drought [1,2].
In recent decades, episodes of forest mortality related to droughts and heat waves have occurred [2,3,4]. In addition, since the last quarter of the 20th century, an increasing difficulty in the regeneration of Mediterranean forests has been observed [5], frequently associated with increased summer drought that hampers regeneration, causing seedling and even adult tree mortality [6,7,8,9]. Regeneration problems are especially important in the Mediterranean, but have also been identified in boreal areas [10,11].
The Mediterranean climate has two growing seasons, one in spring and the other in autumn, separated by a dry and hot summer period. Overcoming this summer period is harsh for plants, especially for young seedlings. Mediterranean plants have adapted their life cycles to the vegetative periods, which justify the abundance of therophytes and which are sometimes ephemeral. Perennial species must be able to survive the summer period, for which they have developed adaptations [12]; however, despite these adaptations, germination and early development of seedlings is a critical stage. One of the main climate-mediated bottlenecks that limits natural regeneration is initial seedling survival [13]. Seedlings must overcome the first summer period within a few months of life, depending on whether they were germinated in spring or autumn. This period results in drastic selection, with more than 90% of seedlings often dying [14]; many tree species are extremely dependent on summer moisture conditions [7]. There is evidence of tree regeneration failure of some species in the Iberian Peninsula forests, even after reforestation, and of transitions to shrublands [15,16,17]. Effects are more intense in species at their ecological limit, such as Pinus sylvestris L. [9], which are sometimes replaced by drought-tolerant species [18]. However, more dry-tolerant species such as Pinus pinea L. are suffering seed production reductions as a response to climate change [19], affecting regeneration.
Not only pines are affected: Quercus species are also suffering regeneration problems [20,21], causing increased concern about long-term persistence in Central Spain [22]. Quercus species, which form many Mediterranean forests, are resprouters, making regeneration problems less evident; even so, most young specimens are resprouts and not individuals born from acorns.
Recurrent droughts reduce forest resilience and the ability to regenerate [23]. Adult trees positively affected seedling survival; therefore, canopy decline affects regeneration, favoring permanent changes in forest composition and shrub transitions [24,25]. In addition, tree seeds of many Mediterranean species are big to guarantee reserves to the seedlings, limiting the regeneration of the crown influence area [26].
Drier conditions may cause adult mortality to not be compensated by seedling recruitment [27], producing differences between canopy and seedling layer composition [28]; both factors may result in future shifts in species composition.
During the 1950s and 1960s, numerous reforestations were carried out in Central Spain using different pine species, which proved to be successful; the climatic conditions allowed the small pines planted to take root and grow. However, the success rate of current reforestations is much lower, which suggests that changes in climatic parameters affecting forest regeneration have occurred over the last decades.
A major problem in analyzing variations over time in the Mediterranean climate is the enormous inter- and intra-annual variability, requiring a statistical treatment of long series of data to determine whether trends are significant. An example of this irregularity was observed in 2021, when there was heavy snowfall and frost in January in Central Spain at low altitudes, and record temperatures in August of 47.4 °C in Andalusia and 42.7 °C in Madrid [29]. In addition, annual climatic conditions do not necessarily fit with summer ones; a rainy year can be extremely dry in summer, and vice versa.
The aim of this study is to analyze significant changes in climatic and bioclimatic parameters that could be related to seedling survival and forest regeneration, particularly in summer conditions, through the statistical treatment of meteorological data series from the mid-twentieth century to the present.
2. Materials and Methods
To analyze climate variability, we considered five meteorological stations located in Central Spain (Figure 1, Table 1), a Mediterranean climate zone. We searched for complete weather stations with long data series that were located on the southern and northern slopes of the Spanish Central Range, from the foothills to the summits. The area is dominated by pine forests (Pinus pinea L., P. pinaster Aiton, P. nigra J.F. Arnold and P. sylvestris L.) and stands of Quercus rotundifolia Lam., Q. faginea Lam. and Q. pyrenaica Willd.
As pointed out in the introduction, regeneration problems are common throughout Mediterranean Spain, especially in continental areas. Numerous works highlight the regeneration problems of Pinus and Quercus stands in Central Spain [5,17,22,30,31,32,33,34], which sometimes go far back in time. Forest inventories prepared for management plans in this region frequently show an absence of regeneration, even of young trees, which confirms that it is a problem that has been present for decades. Local conditions may mitigate or exacerbate the climate impact on seedlings [15]; sunny exposures are much more unfavorable than shady ones, and even locally, medium shade areas increase seedling survival during dry summers [6]. Stands without regeneration problems are frequently located in shady and higher areas, but despite these exceptions associated with favorable local conditions, regeneration problems are a widespread problem in the region, and therefore can be correlated with the regional climate.
The World Meteorological Organization [35] states that at least 10 years of observations are needed to develop statistical benchmarks, and 30 years for precipitation, although climate trends indicate that such short periods may not be representative. For this study we sought to include at least two precipitation periods (60 years). Meteorological stations with such long time series are scarce, limiting selection possibilities. Firstly, three stations with complete data for 70 years (1952–2021) were selected, two in Madrid city and the other in the Central Range. In order to consider the northern slope, the Segovia station was included, with records for 62 years (1960–2021). Finally, the Colmenar Viejo station was considered with a shorter period of records of 44 years (1978–2021), but was seen as interesting because it is located at an average altitude between Madrid and the Central Range, and at the same elevation as Segovia on the opposite slope. The climatic data for these stations came from the Spanish State Meteorological Agency [36].
Monthly mean temperature and precipitation data were collected for each year, using them to calculate the water balance sheet [37,38] and the ombrothermic diagram [39]. Five study parameters were selected (Table 2): (i) mean temperature (T); (ii) rainfall (R); (iii) potential evapotranspiration (PET), calculated [40] and included in the water balance sheet; (iv) physiological drought (PD), obtained from the water balance sheet; and (v) duration of summer drought (SD), obtained from the ombrothermic diagram.
Three study periods were considered: (i) annual, which is useful as a general reference, but not significant for understanding forest regeneration problems; (ii) summer (S), including the months of July and August, the hottest and driest in this region; (iii) extended summer (SE), which includes June to September, spanning the whole summer but also the end of spring and the beginning of autumn. The summer drought duration had only one period, summer, which, depending on the year, may also extend into spring, autumn or both.
Firstly, simple regressions and ANOVAs were performed for each parameter studied, period and meteorological station, determining whether the results were statistically significant at a confidence level of 95% (p < 0.05); the Pearson correlation coefficient and the R2 value were also obtained. Two multiple regressions were performed for each meteorological station, one using summer parameters and the other with extended summer parameters; the duration of the summer drought was incorporated in both regressions. In the regressions a stepwise approach was used, eliminating at each step the variable that was not statistically significant and had a higher p-value. The process was repeated until a result was obtained in which all the variables considered were statistically significant.
In order to determine variations over the studied period, analysis was carried out by dividing the series into three sub-periods of two decades each: 1962–1981 (excluding station 3191E with no data in that period), 1982–2011 and 2012–2021. In each sub-period, simple regressions for all parameters were conducted to determine whether their variation is statistically significant.
Calculations were performed using Statgraphics Centurion 19 software (®Statgraphics Technologies, Inc., The Plains, VA, USA).
3. Results
To evaluate the results, it is necessary to be aware of the time differences in the data series: 70 years in three stations, 62 in one and 44 in another; comparison of results should be taken with caution when considering the global period of time, although this does not apply to the analysis by sub-periods, which was equal in all the meteorological stations.
There seems to be a slight increase in the duration of summer drought, but the ANOVAs performed for the simple regressions (Table 3) indicate that it is not statistically significant in any case. For precipitation, in most cases the annual, summer and expanded summer precipitation have negative correlation coefficients, which would point to a reduction, but in 87% of the cases the result is not statistically significant. Conversely, the increase in temperatures is statistically representative in almost all cases (93%), with R2 values between 12 and 65%; only the annual temperature variation at station 3191E, with a shorter data series, is not significant. PET is statistically significant for all stations and periods, with R2 values between 9 and 65%. PD is statistically significant in the three stations with the longest study period, with R2 values between 11 and 30%; at station 2465 it is only significant in summer and at station 3191E there is no period of statistical significance for this parameter.
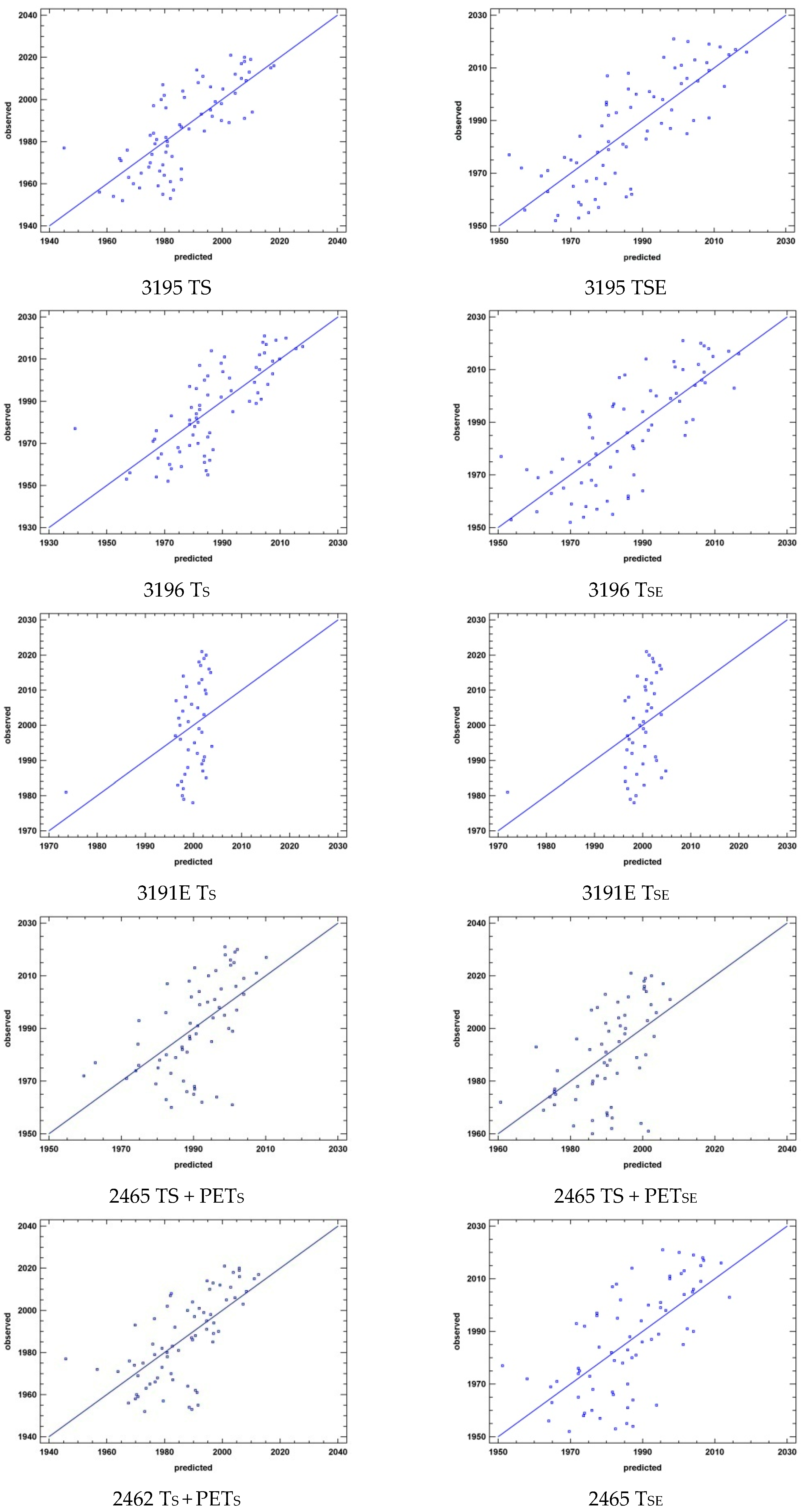
Multiple regressions (Figure 2, Table 4) show a low relationship between variables, which are progressively excluded at each new stage of the regression because they are not statistically significant. The last variable that remains is, in all cases, the temperature. In the summer period, the two northernmost stations (2462 and 2465) also have PETS as a statistically significant variable. In the expanded summer period, only station 2462 also has PETSE as a statistically significant variable.
To assess the variability of the temporal series, simple regressions were performed for all parameters and for all stations in three sub-periods of two decades each (Table 5): 1962–1981 (excluding station 3191E with no data in that period), 1982–2011 and 2012–2021. In the first period (1952–1981), only station 2465 shows a statistically significant variation in TA, PETA and PETS. In the second period (1982–2011), three stations have no significant changes, and two show some significant values in TA, PETA and PDS (3196) and in RA and PDA (2465). In the third period (2012–2021), the only station without significant changes is 2465; in the rest, the increase in annual and summer temperatures is significant, and in three cases, PETS is significant. Only in one case is the variation in summer precipitation significant, but it is an increase, not a reduction.
In conclusion, the reduction in rainfall and the prolongation of the summer drought period were discarded as determining factors in the regeneration problems of the forest stands, as they were not statistically significant. The parameter that better explained these problems was the increase in temperature in all the periods studied, but especially in July and August. This variation was generally non-significant between 1952 and 2001, but became mostly significant between 2002 and 2021. Although variations in the extended summer temperature were significant over the whole period studied, they were not significant in the two-decadal sub-period analysis.
4. Discussion
Progressive regeneration problems in Mediterranean forest stands, especially in pine forests, are often attributed to increased summer drought [6,7,8,9,20]. However, our results show that there was not a significant increase in summer drought in the studied period, at least in the strict sense of the term “drought”. Neither reduction in precipitation nor increases in the length of the summer drought period were statistically significant, as a rule. The increase in summer physiological drought was statistically significant for the whole period, but much less so when analyzing two-decadal sub-periods.
The increase in summer temperatures was statistically significant in all cases, and induced an increase in PET during this period; temperature increments may have had a positive effect on Northern Europe forest growth, but a negative effect on Mediterranean forests due to the increase in evapotranspiration and drought stress [41]. Raising temperatures increased the vapor pressure deficit (VPD), as a 3 °C increase raised the VPD by 45%, which affects water use by plants, exacerbating drought effects [42]. The VPD reflects the potential for the atmosphere to extract water from terrestrial ecosystems [43], and has been established as a major contributor in drought-induced plant mortality [44]. Elevated temperatures may also exacerbate carbon starvation and hydraulic failure [45]. A recent study showed that the impacts of VPD-induced atmospheric dryness on ecosystem production are at least equally, if not more important, than soil moisture [46]. There is evidence on the negative relationship between seedling survival and August average maximum temperature [34].
As a consequence, the problem does not seem to lie in reduced rainfall or length of the dry period as compared to the 1950s or 1960s, but in increased physiological stress induced by rising temperatures, a process that is widespread worldwide [1].
This result is worrying for Mediterranean forest stands, since the regionalized climate change scenarios for Spain [47] point to an increase in maximum temperatures and PET until 2100 (Table 6), with values for the latter parameter in the study area ranging between 8–10% for the RCP 4.5 scenario and 20–34% for the RCP 8.5 scenario. With this variation, the tipping point for the natural regeneration of some species could be passed, something that may already have happened in some locations. Projections in the US show that if the VPD continues to increase as projected by climate models, forest drought stress in the 2050s will exceed that of the most severe droughts in the last 1000 years [48].
The species most affected by this increase in summer temperatures are those located at their ecological limit [9,30]. Among them, two significant species are Pinus sylvestris and Quercus pyrenaica. The former has wide altitudinal range, between 1100 to 2200 m, allowing for greater adaptive capacity; in the future the lower stands will probably decline due to a lack of regeneration, and the species will take refuge at higher altitudes, where the increase in summer temperatures is less critical. However, populations of this species without altitudinal shift possibilities may suffer increased drought-induced mortality [49] or even disappear progressively or abruptly in the most unfavorable situations [50]. The second species has a narrow ecological range; the expected dramatic reductions in the extension of sub-Mediterranean environments [51] where this species grows and its limited ability to move to higher or lower elevations makes it likely to disappear or become rare in the medium or long term. Higher altitude stands have improved resistance and resilience [52], a result consistent with our findings, as altitude reduced temperature, and with it, the VPD. Other pine species, such as P. nigra and P. pinaster, which occupy mid-mountain areas between 800 and 1200 m, are also suffering regeneration problems, and will also have to move up the mountains to compensate for the increase in temperatures; in mixed stands of P. pinea and P. pinaster in Central Spain, a lack of regeneration of the second species has been observed [17,34]. Altitudinal shifts are already occurring; for example, with Fagus sylvatica L. in Northwest Spain [53]. Changes in climate will lead to a progressive dominance of Mediterranean species and a rarefaction of boreo-alpine species located at their southern limit of distribution [54], such as P. sylvestris in the studied area.
The increase in temperatures observed since the 1950s, which has been particularly significant in the last 20 years, will continue to increase with an intensity that will depend on human efforts to mitigate climate change. Consequently, the problems of regeneration of Mediterranean forests will increase in the future, reaching tipping points in many places (probably already reached in some sites), where regeneration of the currently existing main species will be impossible. If no action is taken, the stands will age without regeneration until they will eventually disappear as the trees die of old age.
It is important to identify these tipping points in order to facilitate the adaptation of forests to climate change. Some options include: assisted regeneration, modifying the main species if necessary; use of plants from drought-resistant provenance regions; inclusion of resprouter species; adaptation of nursery plants to water stress; use of containers according to the species root growth; use of water-retainer hydrogels in plantations; microcatchments for runoff harvesting; treeshelters; deeper planting holes; organic amendments; and biotic interactions to facilitate establishment [17,55,56,57,58].
Change processes in forests are often slow, but may be inexorable. Therefore, the future of these forests will depend on the actions undertaken at present.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vilalta, J.; Lloret, F.; Breshears, D.D. Drought-induced forest decline: Causes, scope and implications. Biol. Lett. 2012, 8, 689–691. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Lloret, F.; Montoya, R. Severe drought effects on Mediterranean woody flora in Spain. For. Sci. 2001, 47, 214–218. [Google Scholar] [CrossRef]
- Landmann, G.; Dreyer, E. Impacts of drought and heat on forests. Synthesis of available knowledge, with emphasis on the 2003 event in Europe. Ann. For. Sci. 2006, 63, 567–568. [Google Scholar] [CrossRef] [Green Version]
- Pardos, M.; Madrigal, G.; de Dios-García, J.; Gordo, J.; Calama, R. Sapling recruitment in mixed stands in the Northern Plateau of Spain: A patch model approach. Trees-Struct. Funct. 2021, 35, 2043–2058. [Google Scholar] [CrossRef]
- Marañón, T.; Zamora, R.; Villar, R.; Zavala, M.A.; Quero, J.L.; Pérez-Ramos, I.; Mendoza, I.; Castro, J. Regeneration of tree species and restoration under constrasted Mediterranean habitats: Field and glasshouse experiments. Int. J. Ecol. Environ. Sci. 2004, 30, 187–196. [Google Scholar]
- Matías, L.; Zamora, R.; Castro, J. Sporadic rainy events are more critical than increasing of drought intensity for woody species recruitment in a Mediterranean community. Oecologia 2012, 169, 833–844. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.J.; Dobbertin, M.; Fernández-Cancio, Á.; Vilà-Cabrera, A.; Manzanedo, R.D.; Zavala, M.A.; Navarro-Cerrillo, R.M. Contrasting vulnerability and resilience to drought-induced decline of densely planted vs. natural rear-edge Pinus nigra forests. For. Ecol. Manag. 2013, 310, 956–967. [Google Scholar] [CrossRef]
- Marqués, L.; Madrigal-González, J.; Zavala, M.A.; Camarero, J.J.; Hartig, F. Last-century forest productivity in a managed dry-edge Scots pine population: The two sides of climate warming. Ecol. Appl. 2018, 28, 95–105. [Google Scholar] [CrossRef]
- Hogg, E.H.; Schwarz, A.G. Regeneration of planted conifers across climatic moisture gradients on the Canadian prairies: Implications for distribution and climate change. J. Biogeogr. 1997, 24, 527–534. [Google Scholar] [CrossRef]
- Dulamsuren, C.; Wommelsdorf, T.; Zhao, F.; Xue, Y.; Zhumadilov, B.Z.; Leuschner, C.; Hauck, M. Increased summer temperatures reduce the growth and regeneration of Larix sibirica in southern boreal forests of Eastern Kazakhstan. Ecosystems 2013, 16, 1536–1549. [Google Scholar] [CrossRef]
- Nardini, A.; Lo Gullo, M.A.; Trifilò, P.; Salleo, S. The challenge of the Mediterranean climate to plant hydraulics: Responses and adaptations. Environ. Exp. Bot. 2014, 103, 68–79. [Google Scholar] [CrossRef]
- Calama, R.; Manso, R.; Lucas-Borja, M.; Espelta, J.; Piqué, M.; Bravo, F.; del Peso, C.; Pardos, M. Natural regeneration in Iberian pines: A review of dynamic processes and proposals for management. For. Syst. 2017, 26, eR02S. [Google Scholar] [CrossRef]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Seedling establishment of a boreal tree species (Pinus sylvestris) at its southernmost distribution limit: Consequences of being in a marginal Mediterranean habitat. J. Ecol. 2004, 92, 266–277. [Google Scholar] [CrossRef]
- Benavides, R.; Escudero, A.; Coll, L.; Ferrandis, P.; Ogaya, R.; Gouriveau, F.; Peñuelas, J.; Valladares, F. Recruitment patterns of four tree species along elevation gradients in Mediterranean mountains: Not only climate matters. For. Ecol. Manag. 2016, 360, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Karavani, A.; Boer, M.M.; Baudena, M.; Colinas, C.; Díaz-Sierra, R.; Pemán, J.; Luis, M.; Enríquez-de-Salamanca, Á.; Resco, V. Fire induced deforestation in drought-prone Mediterranean forests: Drivers and unknowns from leaves to communities. Ecol. Monogr. 2018, 88, 141–169. [Google Scholar] [CrossRef] [Green Version]
- Enríquez-de-Salamanca, Á. Dynamics of mediterranean pine forests reforested after fires. J. For. Res. 2022. [CrossRef]
- Gazol, A.; Camarero, J.J.; Sangüesa-Barreda, G.; Vicente-Serrano, S.M. Post-drought resilience after forest die-off: Shifts in regeneration, composition, growth and productivity. Front. Plant Sci. 2018, 871, 1546. [Google Scholar] [CrossRef]
- Mutke, S.; Gordo, J.; Gil, L. Variability of Mediterranean stone pine cone production: Yield loss as response to climate change. Agric. For. Meteorol. 2005, 132, 263–272. [Google Scholar] [CrossRef]
- Gentilesca, T.; Colangelo, M.; Nolè, A.; Ripullone, F.; Camarero, J.J. Drought-induced oak decline in the western Mediterranean region: An overview on current evidences, mechanisms and management options to improve forest resilience. iForest 2017, 10, 796–806. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Fayos, P.; Monleon, V.J.; Espigares, T.; Nicolau, J.M.; Bochet, E. Increasing aridity threatens the sexual regeneration of Quercus ilex (holm oak) in Mediterranean ecosystems. PLoS ONE 2020, 15, e0239755. [Google Scholar] [CrossRef] [PubMed]
- Plieninger, T.; Rolo, V.; Moreno, G. Large-scale patterns of Quercus ilex, Quercus suber, and Quercus pyrenaica regeneration in Central-Western Spain. Ecosystems 2010, 13, 644–660. [Google Scholar] [CrossRef]
- Lloret, F.; Siscart, D.; Dalmases, C. Canopy recovery after drought dieback in holm-oak Mediterranean forests of Catalonia (NE Spain). Glob. Chang. Biol. 2004, 10, 2092–2099. [Google Scholar] [CrossRef]
- Ibáñez, B.; Gómez-Aparicio, L.; Stoll, P.; Ávila, J.M.; Pérez-Ramos, I.M.; Marañón, T. A Neighborhood analysis of the consequences of Quercus suber decline for regeneration dynamics in Mediterranean forests. PLoS ONE 2015, 10, e0117827. [Google Scholar] [CrossRef] [Green Version]
- Saura-Mas, S.; Bonas, A.; Lloret, F. Plant community response to drought-induced canopy defoliation in a Mediterranean Quercus ilex forest. Eur. J. For. Res. 2015, 134, 261–272. [Google Scholar] [CrossRef]
- Calama, R.; Montero, G. Cone and seed production from stone pine (Pinus pinea L.) stands in Central Range (Spain). Eur. J. Forest. Res. 2017, 126, 23–35. [Google Scholar] [CrossRef]
- Lloret, F.; Peñuelas, J.; Prieto, P.; Llorens, L.; Estiarte, M. Plant community changes induced by experimental climate change: Seedling and adult species composition. Perspect. Plant Ecol. Evol. Syst. 2009, 11, 53–63. [Google Scholar] [CrossRef]
- Pérez-Ramos, I.M.; Marañón, T. Community-level seedling dynamics in Mediterranean forests: Uncoupling between the canopy and the seedling layers. J. Veg. Sci. 2012, 23, 526–540. [Google Scholar] [CrossRef] [Green Version]
- WMO. State of the Global Climate 2021; World Meteorological Organization: Gèneve, Switzerland, 2022; Available online: https://library.wmo.int/doc_num.php?explnum_id=11178 (accessed on 5 June 2022).
- Gea-Izquierdo, G.; Montes, F.; Gavilán, R.G.; Cañellas, I.; Rubio, A. Is this the end? Dynamics of a relict stand from pervasively deforested ancient Iberian pine forests. Eur. J. For. Res. 2015, 134, 525–536. [Google Scholar] [CrossRef]
- Pardos, M.; Montes, F.; Aranda, I.; Cañellas, I. Influence of environmental conditions on germinant survival and diversity of Scots pine (Pinus sylvestris L.) in central Spain. Eur. J. For. Res. 2007, 126, 37–47. [Google Scholar] [CrossRef]
- Manso, R.; Calama, R.; Madrigal, G.; Pardos, M. A silvicultureoriented spatio-temporal model for germination in Pinus pinea L. in the Spanish Northern Plateau based on a direct seeding experiment. Eur. J. For. Res. 2013, 132, 969–982. [Google Scholar] [CrossRef] [Green Version]
- Manso, R.; Pukkala, T.; Pardos, M.; Miina, J.; Calama, R. Modelling Pinus pinea forest management to attain natural regeneration under present and future climatic scenarios. Can. J. For. Res. 2014, 44, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Fernández, D.; Montes, F.; Sánchez-González, M.; Gordo, F.J.; Cañellas, I. Regeneration dynamics of mixed stands of Pinus pinaster Ait. and Pinus pinea L. in Central Spain. Eur. J. For. Res. 2018, 137, 17–27. [Google Scholar] [CrossRef]
- WMO. Guide to Climatological Practices; World Meteorological Organization, Gèneve, Switzerland. 2018. Available online: https://library.wmo.int/doc_num.php?explnum_id=5541 (accessed on 5 June 2022).
- AEMET. AEMET OpenData. Agencia Estatal de Meteorología. 2022. Available online: https://opendata.aemet.es/centrodedescargas/inicio (accessed on 7 June 2022).
- Thornthwaite, C.W.; Mather, J.R. The water balance. Publ. Climatol. 1955, 8, 5–86. [Google Scholar]
- Thornthwaite, C.W.; Mather, J.R. Instructions for evaluating the water balance. Publ. Climatol. 1957, 10, 185–204. [Google Scholar]
- Gaussen, H.; Bagnouls, F. Dry season and xerothermic index. Bull. Soc. Hist. Nat. Toulouse 1953, 88, 193–240. [Google Scholar]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Viñegla, B.; Lechuga, V.; Linares, J.C. Natural regeneration and drought effects in the Mediterranean basin. In Forest Management of Mediterranean Forests under the New Context of Climate Change: Building Alternatives for the Coming Future; Lucas-Borja, M.E., Ed.; Nova Science Publishers: New York, NY, USA, 2013; pp. 53–69. [Google Scholar]
- Will, R.E.; Wilson, S.M.; Zou, C.B.; Hennessey, T.C. Increased vapor pressure deficit due to higher temperature leads to greater transpiration and faster mortality during drought for tree seedlings common to the forest–grassland ecotone. New Phytol. 2013, 200, 366–374. [Google Scholar] [CrossRef]
- Broz, A.; Retallack, G.J.; Maxwell, T.M.; Silva, L.C.R. A record of vapour pressure deficit preserved in wood and soil across biomes. Sci. Rep. 2021, 11, 662. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Qin, Z.; Lin, S.; Chen, X.; Chen, B.; He, B.; Wei, J.; Yuan, W. Large influence of atmospheric vapor pressure deficit on ecosystem production efficiency. Nat. Commun. 2022, 13, 1653. [Google Scholar] [CrossRef] [PubMed]
- AdapteCCa. Visor de escenarios de cambio climático. Plataforma Sobre Adaptación al Cambio Climático en España. Ministerio para la Transición Ecológica y el Reto Demográfico. 2022. Available online: https://escenarios.adaptecca.es (accessed on 8 June 2022).
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Martinez-Vilalta, J.; Piñol, J. Drought-induced mortality and hydraulic architecture in pine populations of the NE Iberian Peninsula. For. Ecol. Manag. 2002, 161, 247–256. [Google Scholar] [CrossRef]
- Fernández, A.; Navarro, R.M.; Sánchez, R.; Fernández, R.; Manrique, E. Viabilidad fitoclimática de las repoblaciones de pino silvestre (Pinus sylvestris L.) en la Sierra de los Filabres (Almería). Ecosistemas 2011, 20, 124–144. [Google Scholar]
- Sánchez de Dios, R.; Benito-Garzón, M.; Sainz-Ollero, H. Present and future extension of the Iberian submediterranean territories as determined from the distribution of marcescent oaks. Plant. Ecol. 2009, 204, 189–205. [Google Scholar] [CrossRef]
- Rubio-Cuadrado, Á.; Camarero, J.J.; Aspizua, R.; Sánchez-González, M.; Gil, L.; Montes, F. Abiotic factors modulate post-drought growth resilience of Scots pine plantations and rear-edge Scots pine and oak forests. Dendrochronologia 2018, 51, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Ogaya, R.; Boada, M.; Jump, A.S. Migration, invasion and decline: Changes in recruitment and forest structure in a warming-linked shift of European beech forest in Catalonia (NE Spain). Ecography 2007, 30, 829–837. [Google Scholar] [CrossRef]
- Mendoza, I.; Zamora, R.; Castro, J. A seeding experiment for testing tree-community recruitment under variable environments: Implications for forest regeneration and conservation in Mediterranean habitats. Biol. Conserv. 2009, 142, 1491–1499. [Google Scholar] [CrossRef]
- Chirino, E.; Vilagrosa, A.; Cortina, J.; Valdecantos, A.; Fuentes, D.; Trubat, R.; Luis, V.C.; Puértolas, J.; Bautista, S.; Baeza, M.J.; et al. Ecological restoration in degraded drylands: The need to improve the seedling quality and site conditions in the field. In Forest Management; Grossberg, S.P., Ed.; Nova Science Publishers: New York, NY, USA, 2009; pp. 85–158. [Google Scholar]
- Hlásny, T.; Mátyás, C.; Seidl, R.; Kulla, L.; Merganicová, K.; Trombik, J.; Dobor, L.; Barcza, Z.; Konôpka, B. Climate change increases the drought risk in central European forests: What are the options for adaptation? Lesn. Cas. For. J. 2014, 60, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Konnert, M.; Fady, B.; Gömöry, D.; A’Hara, S.; Wolter, F.; Ducci, F.; Koskela, J.; Bozzano, M.; Maaten, T.; Kowalczyk, J. Use and Transfer of Forest Reproductive Material in Europe in the Context of Climate Change; Euforgen, Bioversity International: Rome, Italy, 2015. [Google Scholar]
- Seidel, H.; Schunk, C.; Matiu, M.; Menzel, A. Diverging drought resistance of scots pine provenances revealed by infrared thermography. Front. Plant Sci. 2016, 7, 1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Location of the studied meteorological stations.

Figure 2.
Multiple regression results. Meteorological station codes in Table 1; studied parameter abbreviations in Table 2.


{kind=link}
{kind=link}
Table 1.
Meteorological stations studied.
| Code | Name | X | Y | Elevation | Period |
|---|---|---|---|---|---|
| 3195 | Madrid—Retiro | 442470 | 4473702 | 667 m | 1952–2021 |
| 3196 | Madrid—Cuatro Vientos | 433267 | 4469738 | 690 m | 1952–2021 |
| 3191E | Colmenar Viejo-FAMET | 435367 | 4505305 | 1004 m | 1978–2021 |
| 2465 | Segovia | 405190 | 4533294 | 1005 m | 1960–2021 |
| 2462 | Puerto de Navacerrada | 414745 | 4516276 | 1894 m | 1952–2021 |
Coordinates EPSG: 25,830 (ETRS89/UTM zone 30N).
Table 2.
Studied parameters.
| Parameter | Code | Period | Unit |
|---|---|---|---|
| Mean temperature | TA | Annual | °C |
| TS | July–August | °C | |
| TSE | June–September | °C | |
| Rainfall | RA | Annual | mm |
| RS | July–August | mm | |
| RSE | June–September | mm | |
| Potential evapotranspiration | PETA | Annual | mm |
| PETS | July–August | mm | |
| PETSE | June–September | mm | |
| Physiological drought | PDA | Annual | mm |
| PDS | July–August | mm | |
| PDSE | June–September | mm | |
| Summer drought | SD | Annual | days |
Table 3.
Simple regressions and ANOVAs for the meteorological stations and parameters analyzed.
| Par | 3195 | 3196 | 3191E | 2465 | 2462 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p | r | R2 | p | r | R2 | p | r | R2 | p | Corr | r | p | r | R2 | |
| SD | 0.4830 | 0.0852 | 0.7264 | 0.3520 | 0.1129 | 1.2750 | 0.6480 | 0.0708 | 0.5001 | 0.1552 | 0.1717 | 2.9482 | 0.4709 | 0.0933 | 0.8697 |
| TA | 0.0000 | 0.7921 | 62.7371 | 0.0000 | 0.8033 | 64.5347 | 0.0875 | 0.2606 | 6.7937 | 0.0000 | 0.7163 | 51.3114 | 0.0000 | 0.6194 | 38.3613 |
| TS | 0.0000 | 0.7527 | 56.6554 | 0.0000 | 0.7526 | 56.6352 | 0.0177 | 0.3561 | 12.6823 | 0.0032 | 0.3473 | 12.0624 | 0.0008 | 0.4140 | 17.1413 |
| TSE | 0.0000 | 0.7806 | 60.9265 | 0.0000 | 0.7705 | 59.3644 | 0.0110 | 0.3798 | 14.4259 | 0.0000 | 0.6699 | 44.8790 | 0.0002 | 0.4620 | 21.3438 |
| RA | 0.2638 | −0.1354 | 1.8325 | 0.0480 | −0.2372 | 5.6745 | 0.7948 | 0.0403 | 0.1628 | 0.1982 | −0.1557 | 2.4228 | 0.7554 | −0.0404 | 0.1630 |
| RS | 0.6845 | −0.0494 | 0.2443 | 0.5273 | −0.0768 | 0.5903 | 0.7365 | −0.0522 | 0.2723 | 0.9836 | 0.0025 | 0.0006 | 0.4111 | 0.1063 | 1.1292 |
| RSE | 0.1914 | −0.1580 | 2.4967 | 0.0696 | −0.2182 | 4.7613 | 0.3916 | −0.1324 | 1.7525 | 0.0099 | −0.3063 | 9.3802 | 0.2463 | −0.1495 | 2.2341 |
| PETA | 0.0000 | 0.7908 | 62.5410 | 0.0000 | 0.8029 | 64.4627 | 0.0000 | 0.5859 | 34.3324 | 0.0000 | 0.6123 | 37.4948 | 0.0000 | 0.5802 | 33.6659 |
| PETS | 0.0000 | 0.7417 | 55.0156 | 0.0000 | 0.7401 | 54.7804 | 0.0445 | 0.3045 | 9.2696 | 0.0000 | 0.4703 | 22.1158 | 0.0083 | 0.0332 | 11.0506 |
| PETSE | 0.0000 | 0.7735 | 59.8260 | 0.0000 | 0.7611 | 57.9236 | 0.0363 | 0.3165 | 10.0180 | 0.0000 | 0.6181 | 38.2090 | 0.0020 | 0.3850 | 14.8222 |
| PDA | 0.0005 | 0.4027 | 16.2135 | 0.0002 | 0.4258 | 18.1314 | 0.5348 | 0.0961 | 0.9242 | 0.0025 | 0.3556 | 12.6419 | 0.1916 | 0.1681 | 2.8252 |
| PDS | 0.0000 | 0.5510 | 30.3551 | 0.0000 | 0.5320 | 28.3032 | 0.1396 | 0.2263 | 5.1233 | 0.0046 | 0.3351 | 11.2291 | 0.0000 | 0.5647 | 31.8839 |
| PDSE | 0.0000 | 0.4755 | 22.6131 | 0.0000 | 0.5333 | 28.4355 | 0.0953 | 0.2547 | 6.4845 | 0.0021 | 0.3610 | 13.0348 | 0.1002 | 0.2107 | 4.4411 |
Table 4.
Multiple regressions and ANOVAs for summer and extended summer periods.
| Period | Est | 1st Regression | 2nd Regression | 3rd Regression | 4rd Regression | Remain | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p | R2adj | Out | p | R2adj | Out | p | R2adj | Out | p | R2adj | Out | |||
| Summer SD + ∑XS | 3195 | 0.0000 | 54.9726 | SD | 0.0000 | 55.6536 | PDS | 0.0000 | 56.2178 | RS | 0.0000 | 56.1576 | PETS | TS |
| 3196 | 0.0000 | 54.2578 | SD | 0.0000 | 54.9487 | PDS | 0.0000 | 55.6154 | RS | 0.0000 | 56.0413 | PETS | TS | |
| 3191E | 0.2872 | 3.2982 | RS | 0.1795 | 5.7636 | SD | 0.0967 | 8.0560 | PETS | 0.0444 | 9.9092 | PDS | TS | |
| 2465 | 0.0001 | 31.0426 | SD | 0.0000 | 32.0316 | RS | 0.0000 | 32.6101 | PDS | 0.0000 | 30.3015 | - | TS + PETS | |
| 2462 | 0.0000 | 43.8003 | SD | 0.0000 | 44.6218 | PDS | 0.0000 | 43.6710 | RS | 0.0000 | 43.2437 | - | TS + PETS | |
| Expanded summer SD + ∑XSE | 3195 | 0.0000 | 57.4294 | PDSE | 0.0000 | 58.0825 | PETSE | 0.0000 | 58.6507 | RSE | 0.0000 | 59.1684 | SD | TSE |
| 3196 | 0.0000 | 60.0179 | PETSE | 0.0000 | 60.5997 | RSE | 0.0000 | 61.1009 | PDSE | 0.0000 | 61.3526 | SD | TSE | |
| 3191E | 0.1071 | 10.0407 | PDSE | 0.0587 | 12.1870 | PETSE | 0.0388 | 12.6003 | SD | 0.0217 | 12.9929 | RSE | TSE | |
| 2465 | 0.0002 | 29.0314 | SD | 0.0001 | 30.2764 | RSE | 0.0001 | 27.3419 | PDSE | 0.0000 | 26.9267 | - | TSE + PETSE | |
| 2462 | 0.0000 | 42.6008 | RSE | 0.0000 | 43.4835 | PDSE | 0.0000 | 43.7984 | SD | 0.0000 | 44.1704 | PETSE | TSE | |
Table 5.
Simple regressions for the stations and parameters analyzed for 20-year periods.
| Par | 1962–1981 | 1982–2001 | 2002–2021 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3195 | 3196 | 2465 | 2462 | 3195 | 3196 | 3191E | 2465 | 2462 | 3195 | 3196 | 3191E | 2465 | 2462 | |
| SD | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| TA | - | - | - | - | - | 0.0154 | - | - | - | 0.0256 | 0.0235 | 0.0200 | - | 0.0142 |
| TS | - | - | 0.0353 | - | - | - | - | - | - | 0.0042 | 0.0251 | 0.0165 | - | 0.0500 |
| TSE | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| RA | - | - | - | - | - | - | - | 0.029 | - | - | - | - | - | - |
| RS | - | - | - | - | - | - | - | - | - | - | 0.0307 | - | - | - |
| RSE | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| PETA | - | - | 0.0266 | - | - | 0.0098 | - | - | - | - | - | - | - | - |
| PETS | - | - | 0.0421 | - | - | - | - | - | - | 0.0048 | 0.0284 | 0.0271 | - | - |
| PETSE | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| PDA | - | - | - | - | - | - | - | 0.0196 | - | - | - | - | - | - |
| PDS | - | - | - | - | - | 0.0195 | - | - | - | - | - | - | - | - |
| PDSE | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
Only p-values of statistically significant results are shown.
Table 6.
Expected PET values for different climate change scenarios.
| Meteorological Station | Municipality | PET (mm/Month) | |||
|---|---|---|---|---|---|
| Scenario RCP 4.5 | Scenario RCP 8.5 | ||||
| 2021 | 2100 | 2021 | 2100 | ||
| 3195, 3196 | Madrid | 66.98 | 72.37 | 68.56 | 92.03 |
| 3191E | Colmenar Viejo | 57.47 | 62.60 | 58.70 | 70.17 |
| 2465 | Segovia | 63.41 | 69.99 | 64.67 | 78.79 |
| 2462 | San Ildefonso | 63.88 | 70.03 | 65.26 | 79.54 |
Data obtained from [47].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Enríquez-de-Salamanca, Á. Effects of Climate Change on Forest Regeneration in Central Spain. Atmosphere 2022, 13, 1143. https://doi.org/10.3390/atmos13071143
AMA Style
Enríquez-de-Salamanca Á. Effects of Climate Change on Forest Regeneration in Central Spain. Atmosphere. 2022; 13(7):1143. https://doi.org/10.3390/atmos13071143
Chicago/Turabian StyleEnríquez-de-Salamanca, Álvaro. 2022. "Effects of Climate Change on Forest Regeneration in Central Spain" Atmosphere 13, no. 7: 1143. https://doi.org/10.3390/atmos13071143
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.