1. Introduction
Non-thermal pressure plasma or non-thermal plasma (NTP) generates plasma ionizing gases that damage biological targets (e.g., cancer cells) without increasing the temperature in the treated regions through the formation of free radicals, photons, positive ions, free electrons, and an electromagnetic field. Cold atmospheric plasma (CAP), which is an NTP device, is a neutral ionized gas consisting of positive charge ions, neutral particles, electrons, and UV radiation at near room temperature. NTP has been widely applied to medical therapy, biomedical materials, and food technology for decades [
1,
2,
3]. Several reactive oxygen species (ROS) and nitrogen species (RNS) are created in NTP [
4]. In food technology, NTP, including CAP, is an innovative technique used to improve the inactivation of pathogens (e.g.,
Salmonella typhimurium) [
5] or spoilage organisms (e.g.,
Saccharomyces cerevisiae) [
3], parasite control [
6], seed germination [
7,
8], and acceleration of seed growth [
8,
9]. Compared with traditional food techniques, NTP may offer several advantages, including that it can be used at room temperature in a dry environment; is a fast, safe treatment; and there are no chemical residues developed under operating conditions. It is an eco-friendly, highly sensitive, economic approach to plant health [
10,
11].
Treatment with NTP may modify the surface of the seed and notably induce the germination process, make it easy for the seed to take in nutrients and water, and change its affinity to water [
12]. NTP can damage the seed envelope, and its reactive species can penetrate the seed coat to disrupt the inner embryo and DNA [
12,
13]. Plasma techniques have been applied for agricultural purposes, especially for the growth of vegetation, for a decade. The positive impact of NTP treatment on the acceleration of germination and improvement of root, shoot, and young seedling growth has been demonstrated in previous studies [
8,
9,
12,
13,
14].
Djulis (
Chenopodium formosanum Koidz.), commonly referred to as Taiwanese red quinoa, is a traditional pseudocereal crop that is an endemic species and a local flora in Taiwan. Botanically speaking, djulis belongs to the Amaranthaceae family under the genus
Chenopodium and has a close botanical similarity to quinoa (
Chenopodium quinoa) [
15]. Djulis has been cultivated by the aborigines in southern Taiwan for over a hundred years, and, traditionally, it is used as an ingredient in local winemaking and as a major part of the aboriginal diet [
16]. Djulis has been documented to be a rich source of minerals, vitamins, and high-quality proteins and provides all eight essential amino acids [
17,
18]. However, scientific interest in djulis only surfaced after a seminal study revealed that the bioactive compound, betacyanin, present in djulis is responsible not only for the plant’s bright red color but also for its strong antioxidant capacities [
18]. In addition, other types of antioxidants, including phytosterols, flavonoids, and phenolic acids, have been isolated from djulis [
2,
15]. More recently, in vitro and in vivo studies have shown that the bioactive compounds present in djulis are responsible for its ability to lower low-density-lipoprotein cholesterol levels [
19], protect the liver from injury [
20,
21], ameliorate dermal damage [
17], and demonstrate chemopreventive effects against colon cancer [
22,
23].
This is a preliminary study intended to examine whether levels of natural bioactive compounds, particularly for the chemicals such as polyphenols, betacyanin, anthocyanin, flavonoids, and 2,2-diphenyl-1-picrylhydrazyl (DPPH)radical scavenging effects against oxidative stress and inflammation, in djulis seed are changed after the seed is treated with NTP (air and nitrogen (N2) NTP).
2. Materials and Methods
2.1. Chemicals and Reagents
Chemicals including rutin, quercetin, DPPH, epicatechin, 6-di-tert-butyl-4-methylphenol (BHT), phthaldialdehyde, and 3-mercaptopropionic acid were purchased from MilliporeSigma (Merck KGaA, Burlington, MA, USA). The methanol and ethanol were obtained from JT-Baker (Fisher Scientific, Loughborough, UK). Formic acid, 2-propanol, and acetonitrile were obtained from Honeywell Research Chemicals (Fisher Scientific).
2.2. Djulis Seeds Treated Using NTP
The djulis seed was directly harvested from the sustainable agricultural farms at National Pingtung University of Science and Technology (SAF/NPUST) or purchased from a retail store carrying products from SAF/NPUST. The atmospheric-pressure AC plasma system used as the NTP (Non-Thermal Atmospheric Pressure Plasma SAP013RT-300RB) was designed and purchased from Creating Nano Technologies Inc. (Tainan, Taiwan). The SAP013RT-300RB, including a plasma jet (torch) and the plasma input power supply (plasma input power: 600–800 W; plasma frequency: 20–40 kHz), was equipped with a three-axis automated glue dispensing robot (LI HSI Technology Co., LTD, Taipei, Taiwan) to treat 1 g (g) of djulis seed in a 60 mm Petri dish covered with a stainless-steel mesh and clipped onto a platform. Djulis seed from the NPUST retail store was treated with air NTP (air-NTP), and the seed from SAF/NPUST was treated with N
2 NTP (N
2-NTP). The high-purity working gases (>99.99%), including air and N
2, were obtained from Air Products and Chemicals, Inc. (Taiwan). The NTP-mediated gaseous ionization and plasma electronics on the seed surface were determined based on the plasma nozzle parameters (A50 (50 mm), A30 (30 mm), and A10 (10 mm)), sweep speed, or contact time (100 and 150 mm/s), input power (600 and 800 W), the distance from the seed to the plasma nozzles (5, 10, 15 mm), and the working gas (
Table 1). The spectrum of the plasma radiation and functional groups of the seed were analyzed using an optical emission spectrometer (OES) (HR 4000CG, Ocean Optics, Orlando, FL, USA) and Fourier-transform infrared spectroscopy (FTIR) (Spectrum One, Perkin Elmer, Waltham, MA, USA), respectively. The morphological properties of the surface of the djulis seed were scanned with a scanning electron microscope (SEM) (S3000N, Hitachi, Tokyo, Japan).
2.3. Analysis of Natural Bioactive Compounds
The seed powder was ground regardless of whether or not it was treated with NTP. For this, 0.1 g of powder was extracted using 10 mL distilled and deionized (DD) water or ethanol for 24 h and then centrifuged for 3500 rpm at 4 °C for 10 min. Betacyanin in the water-extracted fraction was examined with a UV–Visible Spectrometer (DR 5000TM UV–Vis, HACH, Ames, IA, USA) (UV/Vis) at 538 nm. The anthocyanin, total phenolic content (TPC), total flavonoids (TFs), and the DPPH in the ethanol-extract fraction were also determined using UV/Vis. The anthocyanin analysis followed a previous report [
24]. Briefly, 0.1 g of djulis seed powder was extracted with 10 mL of ethanol. After ethanol extraction, the djulis mixture, which was composed with 60% ethanol extract, 40% DD water, and 0.1% HCl, was made. The anthocyanin standards and djulis mixture were tested using UV/Vis at 530 or 657 nm.
The analytical method used for the TPC followed Kujala et al.’s report with minor modifications [
25]. That is, 100 μL of djulis mixture was mixed with 50 μL of Folin–Ciocalteu reagent in a 1.5 mL Eppendorf tube for 5 min, to which 1 mL of 2% sodium carbonate was subsequently added and mixed for 30 min. The standards of gallic acid (0, 50, 100, 150, 200, and 300 ppm) and 200 μL mixture solution from an Eppendorf tube were added into a medium in a 96-well plate analyzed by UV/Vis to determine the TPC. Finally, the TPC of the seed was expressed as mg of gallic acid equivalents per 100 g of the seed (mg GAE/100 g). For the TF analysis, the analytical method was slightly modified from Jia’s study [
26]. In short, 0.5 mL of the djulis mixture was placed in a 15 mL centrifuge tube and mixed with 1.5 mL 95% ethanol, 0.1 mL 10% AlCl
3·6H
2O, 0.1 mL 1 M CH
3COOK, and 2.8 mL H
2O. After mixing for 30 min, 200 μL of the mixture solution was measured using UV/Vis at 415 nm. The standard solution of quercetin (0, 50, 100, 150, 200, 300, and 500 ppm) was made for a TF calibration curve. Finally, the TFs from the djulis seed were expressed as mg of quercetin equivalents per 100 g of the seed (mg QU/100 g). The analytical method used for the DPPH was previously described [
27]. A mixture of 120 μL of the djulis mixture and 80 μL of 0.5 mM DPPH solution was well stirred prior to undergoing a dark reaction for 30 min. The reactant solution and BHT solution as the standard (0, 500, 1500, and 2000 ppm) were examined using UV/Vis at 517 nm. For calculation of the DPPH (%), the % inhibition equation was equal to [A0 − (A1/A0)] × 100%, where A0 and A1 represented the optical density (OD) of the control and the OD of the sample or standards, respectively. Finally, the DPPH (%) was determined as 100% inhibition.
2.4. Analysis of Phenolic Acids and Flavonoids by HPLC/DAD
A high-performance liquid chromatograph with a diode-array UV–VIS detector (HPLC/DAD, Hitachi Chromaster 5110 pump/Hitachi Chromaster 5420) and a reversed phase C
18 HPLC column (ZORBAX Eclipse Plus C
18, 5 μm particle size, I.D. × L 4.6 mm × 25 cm) was used to measure the levels of phenolic acids (gallic acid, chorogenic acid, and 4-hydroxy-3-methoxycinnamic acid) and flavonoids (epicatechin, quercetin, and rutin) (
Figure S1). The djulis mixture was filtered using a 0.45 μm polytetrafluoroethylene member prior to the injection of 20 μL of the extract into a sampling loop. Solvent A (99% DD water and 1% acetic acid, v/v) and solvent B (100% acetonitrile) were used as the mobile phase at a flow rate of 1 mL/min. The gradient of the two solvents in the mobile phase was maintained at 90% solvent A for 10 min, 82% solvent A for 14 min, and maintained at 82% solvent A for 3 min, followed by being decreased to 70% solvent A for 3 min, and retained at 70% solvent A for 10 min, and then increased from 70 to 90% solvent A for 3 min, and kept at 90% solvent A for 5 min. The calibration curve was made using external standards ranging from 0 to 500 ppm. Quality control followed the methods proposed by the Taiwan Food and Drug Administration, including the solvent and sample blank, limits of detection, recovery rate, and the signal-to-noise ratio.
2.5. The Tests of the Thermal Effects
The djulis seed of 1 g was heated in an oven (DO45, DENG YNG, New Taipei, Taiwan) at an oven temperature of 46, 55, or 77 °C and the contact time was set as 5, 10, or 15 s. The thermal-treated and non-thermal control (room temperature) seeds were extracted to be detected for levels of betacyanin, anthocyanin, TFCs, TFs, and DPPH.
2.6. Cell Viability Assays of the THP-1 Cells
The human monocyte cell line, THP-1, was cultured with RPMI 1640 medium (Gibco, Life Technologies, Carlsbad, CA, USA), 10% fetal bovine serum (Gibco, Life Technologies, Carlsbad, CA, USA), and 1% penicillin/streptomycin (Gibco, Life Technologies, Carlsbad, CA, USA) at 37 °C, 5% CO2, and 95% humidity. The THP-1 cells were resuspended in fresh media at a concentration of 2 × 106/mL, 0.1 mL/well in 96-well plates and reacted with 10% Alamar Blue reagent for 30 min. The THP-1 cell viability was determined after exposure of djulis and a carboxylated single-wall carbon nanotube (SWCNT-COOH), which is an engineered nanomaterial that can easily induce oxidative stress and inflammatory responses in in vitro systems, for 24 and 48 h. The fluorescence intensity of the stained cells was measured using excitation at 530–560 nm and emissions at 590 nm. Data were calculated as percent (%) viability relative to the untreated control, which was set at 100%. Three replicates were performed for each of the concentrations.
2.7. Intracellular ROS Staining of THP-1 Cells
After pretreatment with/without 100 μg of the djulis seed extract overnight, the THP-1 cells were resuspended in fresh media at a concentration of 2 × 106/mL, 0.1 mL/well in 96-well plates and reacted with 20 mM of 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) probe (Invitrogen™, Thermo Fisher Scientific, Waltham, MA, USA) for 30 min. Following treatment with SWCNT-COOH, the fluorescence intensity of the stained cells was measured at 485 nm (excitation) and 527 nm (emission) to determine the ROS levels. Cell viability was determined using an Alamar Blue assay. Following treatment with SWCNT-COOH at all concentrations for 48 h, the fluorescence intensity of the stained cells was measured based on the excitation at 485 nm and emissions with the 535 nm. Data were calculated as percent (%) relative to untreated control, which was set at 100%. Three replicates were performed for each of the concentrations.
2.8. Statistical Analysis
All of the measurements were higher than the limit of detection. Non-parametric analyses such as Wilcoxon sign rank tests were used to examine the induced folds of bioactive compounds in the NTP-treated djulis seed compared with the untreated control. The significant differences in the induced folds of bioactive compounds in the djulis seed were tested to compare the air-NTP (n = 3) and N2-NTP (n = 6) using Wilcoxon sign rank tests. All statistical analyses were performed using SPSS Version 12.0 for Windows (SPSS IBM Inc., Armonk, NY, USA).
4. Discussion
Djulis is a local flora in Taiwan. Although several Taiwanese scientists have found that the nutrition, abundant TFs and TPC (particularly rutin and gallic acid), and antioxidants are good for human health, few scientists outside Taiwan are aware of this valuable vegetation [
2,
15,
17,
18]. This study reiterates the value of djulis seed, and an attempt is made to examine the changes in the natural bioactive compounds in seed after NTP treatment.
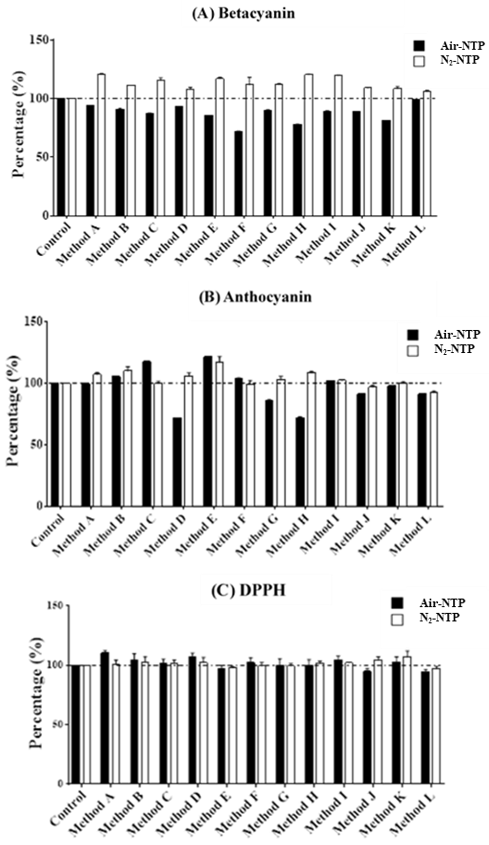
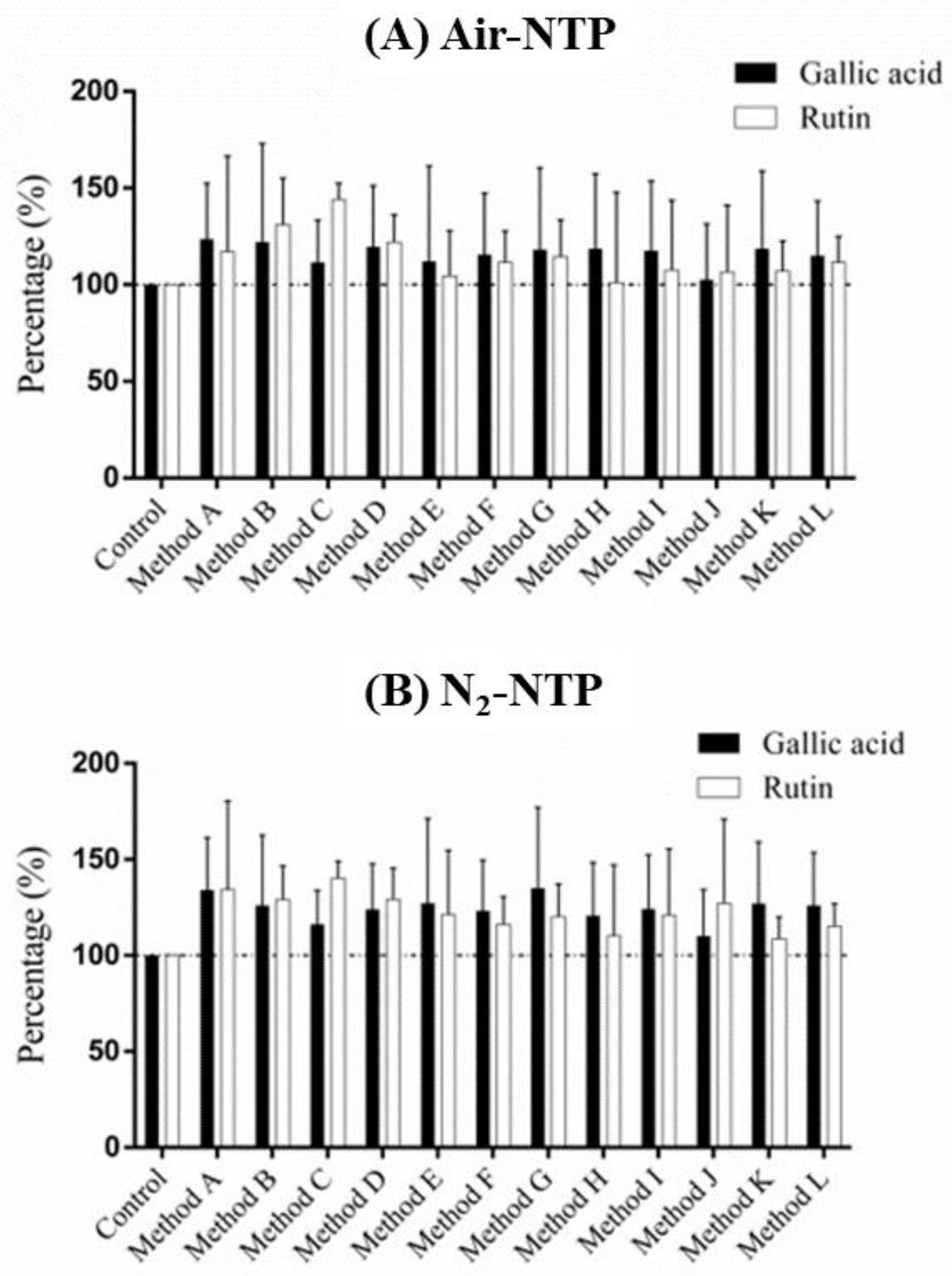
Table 2 shows levels of natural bioactive compounds in the djulis seed without NTP treatment. Few reports have focused on the bioactive compounds in djulis seed. A previous study showed a lower magnitude of TPCs (5.67–6.42 mg GAE/100 g) and TFs (0.472–0.572 mg QU/100 g) in djulis hulls [
15], compared with those from our results for djulis seed. Taiwanese nutritionists emphasize the content of betacyanin and anthocyanin in djulis seed. Tsai et al. (2010) indicated that betacyanin can be separated and identified using liquid chromatography/ion trap mass spectrometry for four species of betacyanin, including betanin, isobetanin, amaranthine, and isoamaranthin. The two bioactive compounds in betanin and isobetanin contributed 73.9% of betacyanin necessary for DPPH scavenging as the major source of the antioxidants [
18]. Huang et al. (2019) also revealed that djulis hulls are a potential source for natural antioxidants in food due to their inhibitory effects on cumulative concentrations of nitric oxide and prostaglandin estradiol [
15]. In both previous and recent studies, scientists have found various abundant bioactive compounds and their associated antioxidants in djulis.
Although treatment with CAP and NTP is common in agricultural applications, most seed studies have concentrated on the notably positive improvements in seed germination, shoot and root growth, and the growth of young seedlings after CAP or NTP treatment [
8,
9,
26,
27]. Previous reports referred to the factors influencing germination in relation to different types of plasma (e.g., the atmospheric pressure dielectric barrier charge (DBD) and low-pressure radiofrequency (RF) plasma) and the plasma working gases (e.g., air, oxygen, nitrogen, and argon) [
8,
25,
27]. An air-CAP treatment was found to notably accelerate the germination of cotton in comparison with an argon-CAP treatment, where the contact time of air-CAP was positively correlated with the hydrophilicity of the cotton seed surface and water uptake [
8]. To the best of our knowledge, few studies have considered the impact of CAP or NTP treatment on variations in bioactive compounds in seed, including djulis. In the present study, air-NTP and N
2-NTP treatment of djulis seed was carried out to measure the temperature on the djulis seed surface at temperatures ranging between 46 and 77 °C, which is slightly higher than the temperature used for CAP treatment (27 °C or 300 K). Compared with other seed studies where CAP treatment was used on seed for a few minutes [
8,
9,
26,
27], in the present study, air-NTP or N
2-NTP was used to treat djulis seed to induce changes in bioactive compounds in a few seconds. In the present study, NTP treatment affected the bioactive compound levels, such as betacyanins, anthocyanins, phenolic acids, and flavonoids, in djulis seed as compared with the untreated seed. According to our findings, the radical scavenging ability of DPPH in the untreated, air-NTP-treated, and N
2-NTP-treated seeds were at quite high levels ranging from 75% to 91%. The positive and negative effects on the bioactive compounds in the djulis seed after NTP treatment appeared to depend on the usage of plasma gas (air and N
2), the plasma input power (600 and 800 W), contact time (0.51–1.02 s), and temperature (46–77 °C) in this study (
Figure 3 and
Figure 4). The findings of the present study were mainly dependent on the plasma working gas (air and N
2) and were slightly independent of the contact temperature and time in terms of influencing the bioactive compound content in djulis seed. The results showed that bioactive compound levels, particularly betacyanin and TPCs, were significantly and positively related to N
2-NTP treatment and, in contrast, were negatively correlated with the air-NTP treatment. This is a new finding, where changes in the natural bioactive compounds of the seed could occur in a few seconds due to NTP treatment especially in the case of N
2-NTP treated djulis seed to activate the content of betacyanin, betacyanins, anthocyanins, TFCs, gallic acid, and rutin. It is still unknown why there were inversely dramatic differences in the amount of activated bioactive compounds in the air-NTP and N
2 -NTP samples.
Our results indicate that plasma etching may be related to surface damage to the djulis seed, where etching and damage were found on the pericarpal surface of air-CAP-treated quinoa seed [
25]. NTP or CAP accelerate the water uptake mainly through the generation of UV radiation, ROS and RNS, as well as specific electric currents due to plasma etching [
8,
9,
25,
26,
27]. Several reports have also shown an eroded structure or surface etching generated in NTP-treated seeds, such as brown rice [
26], cotton seed [
8], and quinoa [
25], intended to improve seed germination rates and seeding growth. It has been revealed that plasma etching on the seed surface may be associated with improvements in germination or may activate various biochemical processes that cause new functional groups on the outermost coat layers of seeds [
25,
28], which, based on our findings, may also be related to increased levels of betacyanin, anthocyanin, TPCs, rutin, and gallic acids in N
2-NTP-treated djulis seed. No thermal effects were found in the present study due to thermal treatment in a few seconds.
In the present study, bioactive compounds, with the exception of TFs, were enhanced by the N
2-NTP treatment, and these chemicals were inhibited by the air-NTP treatment under most testing conditions. According to the different patterns in the OES spectra between the air-NTP and N
2-NTP treatment groups shown in
Figure 1, these results were inferred to be due to the different reactive species generated by treatment of air-NTP and N
2-NTP. Nitrogen oxide species (NO-γ) from 200 to 300 nm possibly were generated by N
2-NTP plasma jet in the OES spectra, mainly due to the collision of the RNS in N
2-NTP with the surrounding air [
28,
29].
Figure 1 shows that differences in the intensities of NO, N
2, and N
2+ on the OES spectra were also found between the air-NTP and N
2-NTP treatment groups. The different species of reactive substances in plasma ionizing gas were induced by plasma discharge with the different working gases. Nitrogen-based reactive species, such as nitrogen dioxide (•NO
2) and nitric oxide (•NO), which are RNS, and nitrous oxide (N
2O), nitrogen trioxide (NO
3), peroxynitrite (ONOO
−), and dinitrogen tetroxide (N
2O
4), which are reactive gases, were generated by air-NTP or N
2-NTP treatment; while oxygen-based reactive species, including ROS generated by air-NTP or O
2-NTP included hydrogen peroxide (H
2O
2), ozone (O
3), hydroxyl, singlet oxygen (
1O
2), hydroxyl radical (•OH), and superoxide (O
2−•) [
30,
31].
The findings of the present study showed that air-NTP treatment inhibited the performance of certain bioactive compounds in the djulis seed, and N
2-NTP enhanced the levels of bioactive compounds. In a quinoa study [
13], ROS and RNS generated etching and damage on the seed surface, in turn modifying chemical and morphological effects on the surface and increasing absorption of nitrate functional groups on the seed. N
2-NTP is intended to generate a greater amount of UV radiation and NOx than air-NTP [
12]. In this study, various nitrogen species were produced after air-NTP, as shown in
Figure 1A. If the findings from the Gómez-Ramírez’s study are true [
13], in the present study, more abundant RNS may have been activated on the surface of the djulis seed after the N
2-NTP treatment as compared with the air-NTP treatment. A large amount of RNS and nitrogen functional groups may have been diffused into the interior of the djulis seed that in turn affected the levels of bioactive compounds and antioxidant properties. In a comet assay conducted in a previous study [
12], no significant differences in DNA damage in soybean seeds were found between a negative control (the untreated control) and an air-CAP treatment group. Inversely, N
2-CAP treatment induced notable and significant DNA damage similar to that in the positive control (the untreated control with addition of zeocin 5 mg/L for 1 h) compared with the negative control. Nitrogen species also possibly penetrated the djulis seed surface and affected the DNA on the chromosome after the NTP treatment. A Korean study revealed that PAW treatment of tomato seedlings enhanced plant growth, generated ROS and RNS, induced inflammation, activated defense hormones, such as salicylic acid and jasmonic acid, and induced pathogenesis-related genes [
10]. N
2-NTP caused DNA damage easily to further activate defense systems such as antioxidant reactions compared with air-NTP. This may explain the enhancement of bioactive compounds in the djulis seed after treatment with N
2-NTP. In a recent study, treating soybean seed with low-dose air/O
2-NTP significantly promoted the Krebs cycle and switched on oxygenic seed germination metabolism from the anoxygenic status. Inversely, high doses of N
2-NTP significantly weakened the Krebs cycle, increased lactate and alcohol dehydrogenase, and caused anoxygenic metabolism [
12]. According to a previous report [
12], N
2-NTP may activate anoxygenic metabolism and in turn, may enhance activation of betacyanin, anthocyanin, TPCs, rutin, and gallic acids due to damage to the djulis seed that activates a protection mechanism after plasma etching. Prior to the germination of seeds, it is still unknown what happens (e.g., mechanism) in terms of changes in bioactive compounds after air-NTP or N
2-NTP treatment on the seed surface due to lack of evidence in current reports.
In this study, it was posited that djulis seed extract may scavenge the free radical, ROS, and RNS in human monocytes, and these findings may be potential on djulis seed extract after air-NTP or N
2-NTP treatment.
Figures S7 and S8 show that, low doses (0.01–0.100 μg/L) of SWCNT-COOH activate the responses of oxidative stress while no cytotoxic effects are found at these doses. Our results also show that the pretreatment with djulis seed extract overnight could inhibit the expression of oxidative stress after THP-1 cells are exposed to SWCNT-COOH at low doses of 0.100 μg/L (
Figure S9). According to a previous study [
32], in vitro and in vivo exposure to SWCNT-COOH induced oxidative stress and generated ROS. Exposure of THP-1 cells to SWCNT caused cytotoxicity, genotoxicity, DNA methylation, and inflammatory responses at 10–100 μg/mL [
33,
34]. Our results show the activation of ROS production in human macrophage-differentiated THP-1 cells at low doses of 0.001–0.100 μg/L without cytotoxicity. With overnight pretreatment of djulis seed extract, the generation of ROS could be inactivated in THP-1 cells. Based on the DPPH results from air-NTP and N
2-NTP, high ROS scavenging effects occurred, and no significant differences were found among the untreated, air-NTP-treated, and N
2-NTP-treated seed. It is thus possible to infer that NTP-treated djulis seed extract could inhibit ROS production or prevent ROS generation particularly in the case of N
2-NTP. This may enhance free radical scavenging and lead to improvements in human health. Further studies are encouraged in the future.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}