
Selection Signatures Reveal Candidate Genes for the Cornish Rex Breed-Specific Phenotype


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample and Data Collection
2.3. DNA Extraction, Fragment Analysis and Sanger Sequencing
2.4. Remapping of Feline SNP Array Coordinates to F.catus_Fca126_mat1.0
2.5. Identification of Selection Signatures
2.6. Annotation of Regions under Selection
3. Results
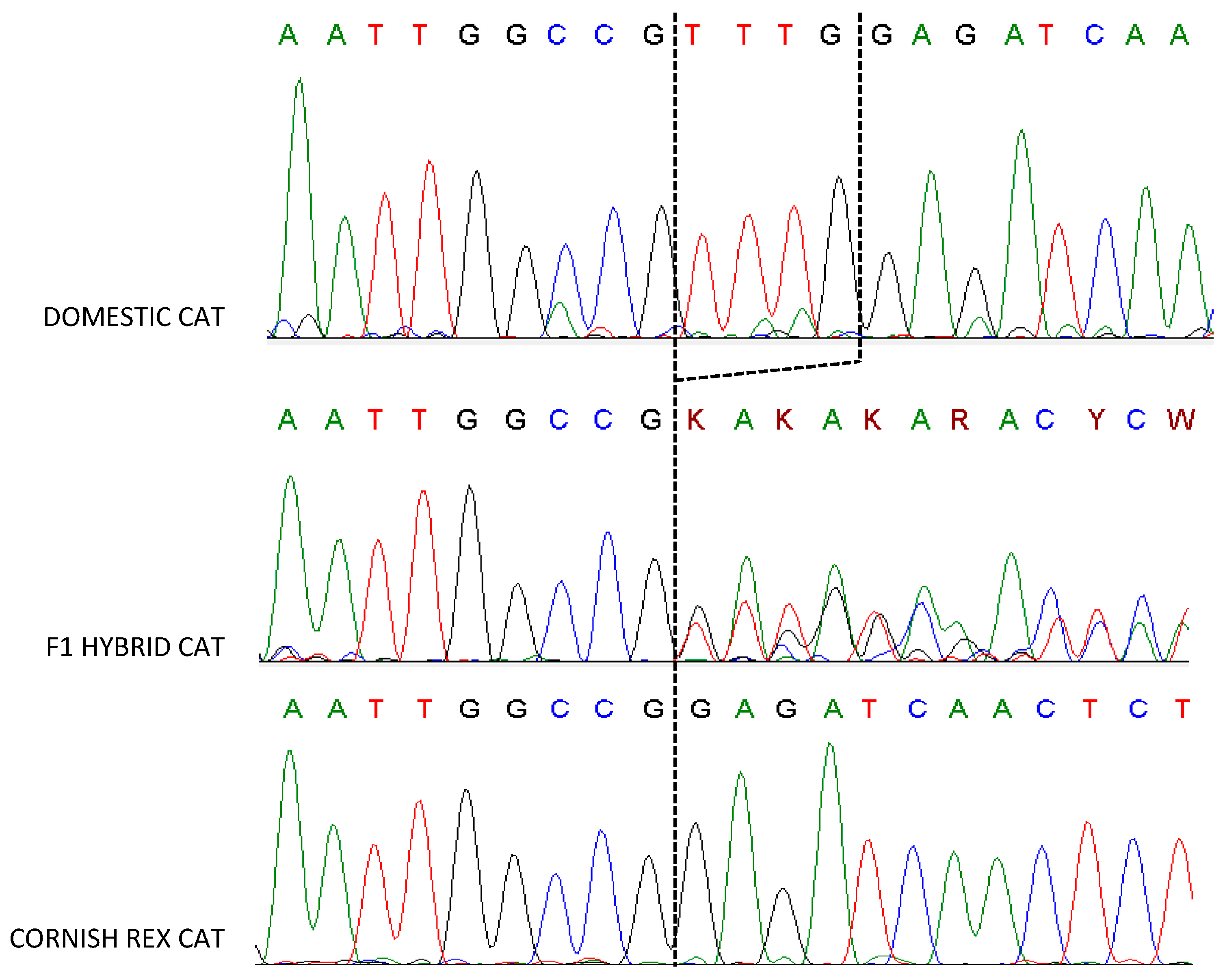
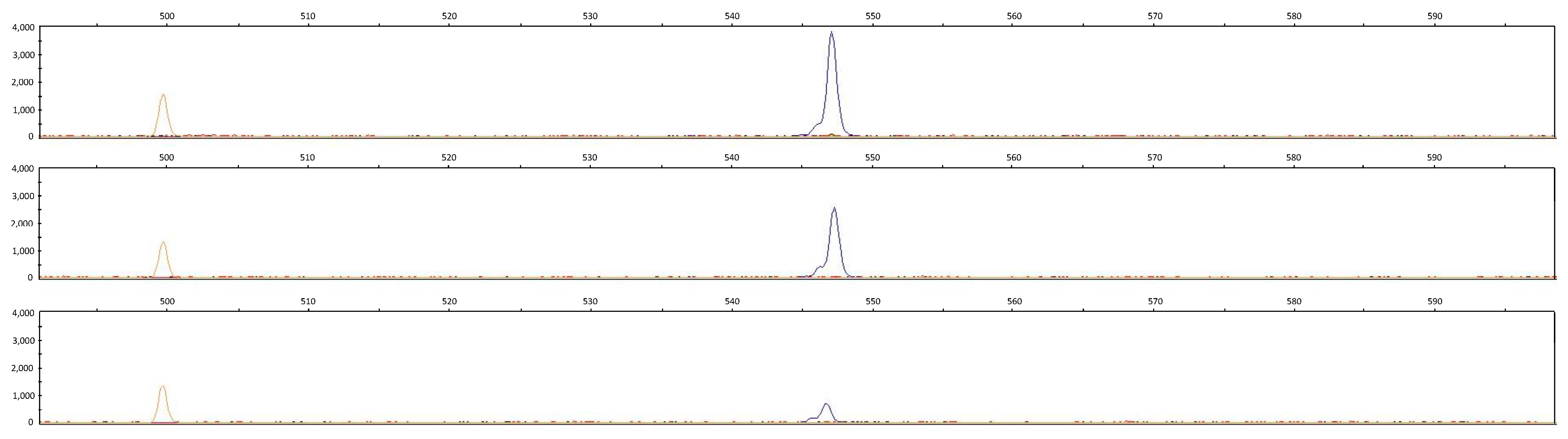
3.1. LPAR6 Mutation
3.2. ALX1 Genotype
3.3. Cornish Rex Phenotype
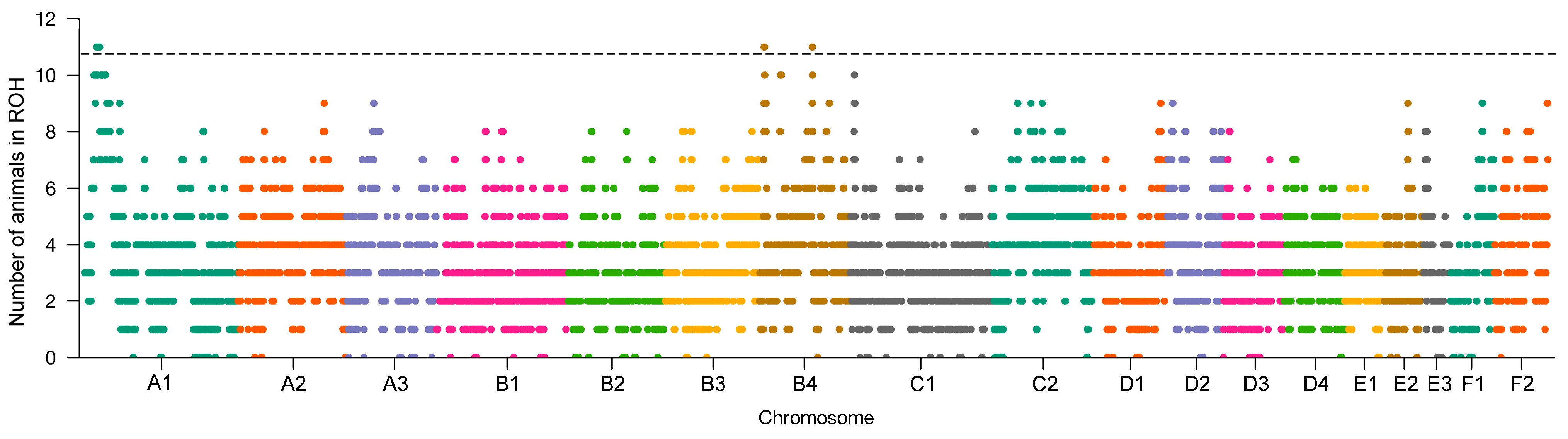
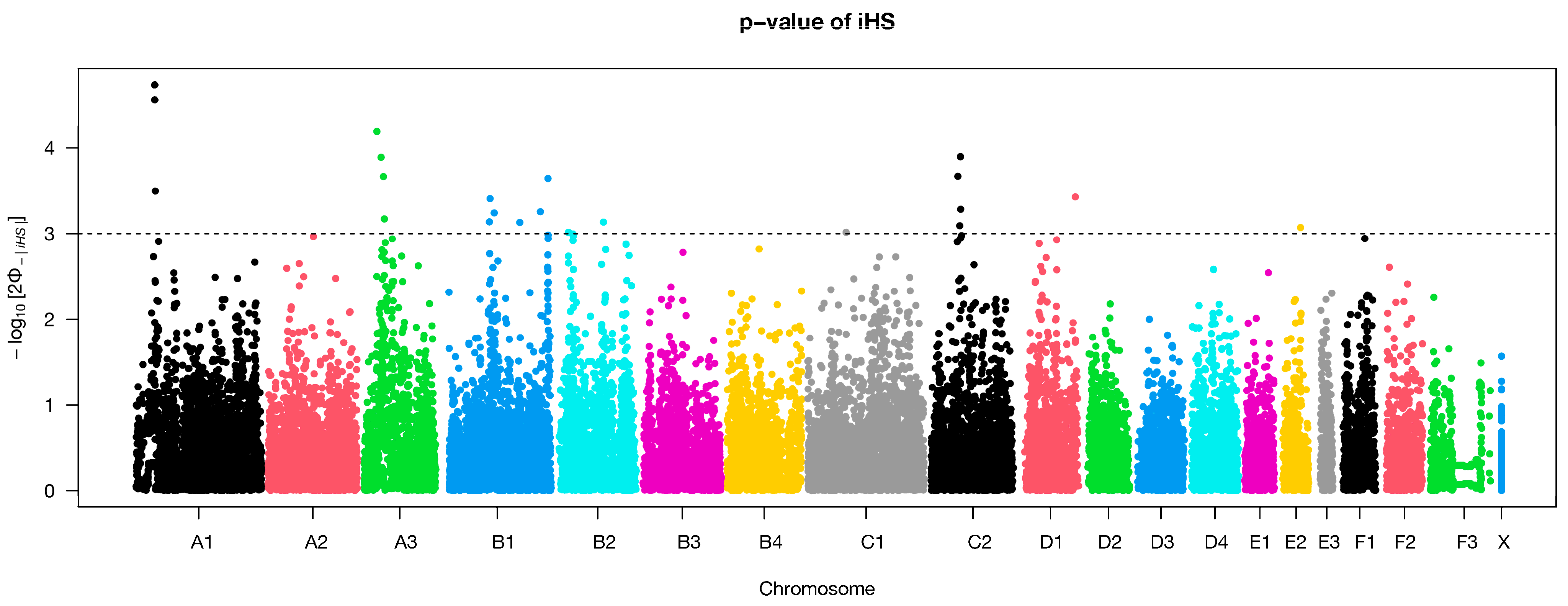
3.4. Selection Signatures
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cameron-Beaumont, C.; Lowe, S.E.; Bradshaw, J.W.S. Evidence suggesting preadaptation to domestication throughout the small Felidae. Biol. J. Linn. Soc. 2002, 75, 361–366. [Google Scholar] [CrossRef]
- Driscoll, C.A.; Macdonald, D.W.; O’Brien, S.J. From wild animals to domestic pets, an evolutionary view of domestication. Proc. Natl. Acad. Sci. USA 2009, 106 (Suppl. S1), 9971–9978. [Google Scholar] [CrossRef]
- Montague, M.J.; Li, G.; Gandolfi, B.; Khan, R.; Aken, B.L.; Searle, S.M.; Minx, P.; Hillier, L.W.; Koboldt, D.C.; Davis, B.W.; et al. Comparative analysis of the domestic cat genome reveals genetic signatures underlying feline biology and domestication. Proc. Natl. Acad. Sci. USA 2014, 111, 17230–17235. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Go, M.; Kang, D.; Kim, J. Genetic Differentiation between Domestic Cats and Wildcats. J. Anim. Breed. Genom. 2023, 7, 9–15. [Google Scholar] [CrossRef]
- Lyons, L.A. DNA mutations of the cat: The good, the bad and the ugly. J. Feline Med. Surg. 2015, 17, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Gandolfi, B.; Alhaddad, H.; Affolter, V.K.; Brockman, J.; Haggstrom, J.; Joslin, S.E.; Koehne, A.L.; Mullikin, J.C.; Outerbridge, C.A.; Warren, W.C.; et al. To the Root of the Curl: A Signature of a Recent Selective Sweep Identifies a Mutation That Defines the Cornish Rex Cat Breed. PLoS ONE 2013, 8, e67105. [Google Scholar] [CrossRef] [PubMed]
- Gandolfi, B.; Alhaddad, H.; Joslin, S.E.; Khan, R.; Filler, S.; Brem, G.; Lyons, L.A. A splice variant in KRT71 is associated with curly coat phenotype of Selkirk Rex cats. Sci. Rep. 2013, 3, 2000. [Google Scholar] [CrossRef] [PubMed]
- Castle, W.E. The rex rabbit. J. Hered. 1929, 20, 193–199. [Google Scholar] [CrossRef]
- Diribarne, M.; Mata, X.; Chantry-Darmon, C.; Vaiman, A.; Auvinet, G.; Bouet, S.; Deretz, S.; Cribiu, E.P.; de Rochambeau, H.; Allain, D.; et al. A deletion in exon 9 of the LIPH gene is responsible for the rex hair coat phenotype in rabbits (Oryctolagus cuniculus). PLoS ONE 2011, 6, e19281. [Google Scholar] [CrossRef]
- Kuramoto, T.; Hirano, R.; Kuwamura, M.; Serikawa, T. Identification of the Rat Rex Mutation as a 7-bp Deletion at Splicing Acceptor Site of the Krt71 Gene. J. Vet. Med. Sci. 2010, 72, 909–912. [Google Scholar] [CrossRef]
- Cadieu, E.; Neff, M.W.; Quignon, P.; Walsh, K.; Chase, K.; Parker, H.G.; Vonholdt, B.M.; Rhue, A.; Boyko, A.; Byers, A.; et al. Coat variation in the domestic dog is governed by variants in three genes. Science 2009, 326, 150–153. [Google Scholar] [CrossRef]
- Bauer, A.; Hadji Rasouliha, S.; Brunner, M.T.; Jagannathan, V.; Bucher, I.; Bannoehr, J.; Varjonen, K.; Bond, R.; Bergvall, K.; Welle, M.M.; et al. A second KRT71 allele in curly coated dogs. Anim. Genet. 2019, 50, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Manakhov, A.D.; Andreeva, T.V.; Rogaev, E.I. The curly coat phenotype of the Ural Rex feline breed is associated with a mutation in the lipase H gene. Anim. Genet. 2020, 51, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Daetwyler, H.D.; Capitan, A.; Pausch, H.; Stothard, P.; van Binsbergen, R.; Brøndum, R.F.; Liao, X.; Djari, A.; Rodriguez, S.C.; Grohs, C.; et al. Whole-genome sequencing of 234 bulls facilitates mapping of monogenic and complex traits in cattle. Nat. Genet. 2014, 46, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Thomer, A.; Gottschalk, M.; Christmann, A.; Naccache, F.; Jung, K.; Hewicker-Trautwein, M.; Distl, O.; Metzger, J. An epistatic effect of KRT25 on SP6 is involved in curly coat in horses. Sci. Rep. 2018, 8, 6374. [Google Scholar] [CrossRef] [PubMed]
- Whiteway, C.E.; Robinson, R. Two recessive rex coat mutants in the guinea pig. J. Hered. 1989, 80, 163–165. [Google Scholar] [CrossRef] [PubMed]
- Gandolfi, B.; Outerbridge, C.A.; Beresford, L.G.; Myers, J.A.; Pimentel, M.; Alhaddad, H.; Grahn, J.C.; Grahn, R.A.; Lyons, L.A. The naked truth: Sphynx and Devon Rex cat breed mutations in KRT71. Mamm. Genome 2010, 21, 509–515. [Google Scholar] [CrossRef]
- Robinson, R. Oregon rex—A fourth rexoid coat mutant in the cat. Genetica 1972, 43, 236–238. [Google Scholar] [CrossRef]
- Pasternack, S.M.; von Kügelgen, I.; Al Aboud, K.; Lee, Y.A.; Rüschendorf, F.; Voss, K.; Hillmer, A.M.; Molderings, G.J.; Franz, T.; Ramirez, A.; et al. G protein-coupled receptor P2Y5 and its ligand LPA are involved in maintenance of human hair growth. Nat. Genet. 2008, 40, 329–334. [Google Scholar] [CrossRef]
- Shimomura, Y.; Wajid, M.; Ishii, Y.; Shapiro, L.; Petukhova, L.; Gordon, D.; Christiano, A.M. Disruption of P2RY5, an orphan G protein–coupled receptor, underlies autosomal recessive woolly hair. Nat. Genet. 2008, 40, 335–339. [Google Scholar] [CrossRef]
- Petukhova, L.; Sousa, E.C.; Martinez-Mir, A.; Vitebsky, A.; dos Santos, L.G.; Shapiro, L.; Haynes, C.; Gordon, D.; Shimomura, Y.; Christiano, A.M. Genome-wide linkage analysis of an autosomal recessive hypotrichosis identifies a novel P2RY5 mutation. Genomics 2008, 92, 273–278. [Google Scholar] [CrossRef]
- Inoue, A.; Arima, N.; Ishiguro, J.; Prestwich, G.D.; Arai, H.; Aoki, J. LPA-producing enzyme PA-PLA1;α regulates hair follicle development by modulating EGFR signalling. Embo J. 2011, 30, 4248–4260. [Google Scholar] [CrossRef]
- Akiyama, M. Isolated autosomal recessive woolly hair/hypotrichosis: Genetics, pathogenesis and therapies. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 1788–1796. [Google Scholar] [CrossRef]
- Searle, A.G.; Jude, A.C. The ‘rex’ type op goat in the domestic cat. J. Genet. 1956, 54, 506–512. [Google Scholar] [CrossRef]
- Robinson, R. Devon rex—A third rexoid coat mutant in the cat. Genetica 1969, 40, 597–599. [Google Scholar] [CrossRef]
- Weiss, H. Rex the King. In CFA Year Book; CFA: Charlottesville, VA, USA, 1965; pp. 138–146. [Google Scholar]
- Alhaddad, H.; Khan, R.; Grahn, R.A.; Gandolfi, B.; Mullikin, J.C.; Cole, S.A.; Gruffydd-Jones, T.J.; Häggström, J.; Lohi, H.; Longeri, M.; et al. Extent of linkage disequilibrium in the domestic cat, Felis silvestris catus, and its breeds. PLoS ONE 2013, 8, e53537. [Google Scholar] [CrossRef]
- Gandolfi, B.; Alhaddad, H.; Abdi, M.; Bach, L.H.; Creighton, E.K.; Davis, B.W.; Decker, J.E.; Dodman, N.H.; Ginns, E.I.; Grahn, J.C.; et al. Applications and efficiencies of the first cat 63K DNA array. Sci. Rep. 2018, 8, 7024. [Google Scholar] [CrossRef] [PubMed]
- Pontius, J.U.; Mullikin, J.C.; Smith, D.R.; Lindblad-Toh, K.; Gnerre, S.; Clamp, M.; Chang, J.; Stephens, R.; Neelam, B.; Volfovsky, N.; et al. Initial sequence and comparative analysis of the cat genome. Genome Res. 2007, 17, 1675–1689. [Google Scholar] [CrossRef]
- Mullikin, J.C.; Hansen, N.F.; Shen, L.; Ebling, H.; Donahue, W.F.; Tao, W.; Saranga, D.J.; Brand, A.; Rubenfield, M.J.; Young, A.C.; et al. Light whole genome sequence for SNP discovery across domestic cat breeds. BMC Genom. 2010, 11, 406. [Google Scholar] [CrossRef]
- Gandolfi, B.; Gruffydd-Jones, T.J.; Malik, R.; Cortes, A.; Jones, B.R.; Helps, C.R.; Prinzenberg, E.M.; Erhardt, G.; Lyons, L.A. First WNK4-hypokalemia animal model identified by genome-wide association in Burmese cats. PLoS ONE 2012, 7, e53173. [Google Scholar] [CrossRef]
- Alhaddad, H.; Gandolfi, B.; Grahn, R.A.; Rah, H.C.; Peterson, C.B.; Maggs, D.J.; Good, K.L.; Pedersen, N.C.; Lyons, L.A. Genome-wide association and linkage analyses localize a progressive retinal atrophy locus in Persian cats. Mamm. Genome 2014, 25, 354–362. [Google Scholar] [CrossRef]
- Davis, B.W.; Seabury, C.M.; Brashear, W.A.; Li, G.; Roelke-Parker, M.; Murphy, W.J. Mechanisms Underlying Mammalian Hybrid Sterility in Two Feline Interspecies Models. Mol. Biol. Evol. 2015, 32, 2534–2546. [Google Scholar] [CrossRef]
- Kreitman, M. Methods to detect selection in populations with applications to the human. Annu. Rev. Genom. Hum. Genet. 2000, 1, 539–559. [Google Scholar] [CrossRef]
- Alhaddad, H.; Abdi, M.; Lyons, L.A. Patterns of allele frequency differences among domestic cat breeds assessed by a 63K SNP array. PLoS ONE 2021, 16, e0247092. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Lyons, L.A.; Erdman, C.A.; Grahn, R.A.; Hamilton, M.J.; Carter, M.J.; Helps, C.R.; Alhaddad, H.; Gandolfi, B. Aristaless-Like Homeobox protein 1 (ALX1) variant associated with craniofacial structure and frontonasal dysplasia in Burmese cats. Dev. Biol. 2016, 409, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Bredemeyer, K.R.; Hillier, L.; Harris, A.J.; Hughes, G.M.; Foley, N.M.; Lawless, C.; Carroll, R.A.; Storer, J.M.; Batzer, M.A.; Rice, E.S.; et al. Single-haplotype comparative genomics provides insights into lineage-specific structural variation during cat evolution. Nat. Genet. 2023, 55, 1953–1963. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J. BLAT—The BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Browning, S.R.; Browning, B.L. Rapid and accurate haplotype phasing and missing-data inference for whole-genome association studies by use of localized haplotype clustering. Am. J. Hum. Genet. 2007, 81, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M.; Vitalis, R. rehh: An R package to detect footprints of selection in genome-wide SNP data from haplotype structure. Bioinformatics 2012, 28, 1176–1177. [Google Scholar] [CrossRef]
- Salonen, M.; Vapalahti, K.; Tiira, K.; Mäki-Tanila, A.; Lohi, H. Breed differences of heritable behaviour traits in cats. Sci. Rep. 2019, 9, 7949. [Google Scholar] [CrossRef] [PubMed]
- Peltz, R. The Exotic Rex. In CFA Year Book; CFA: Charlottesville, VA, USA, 1974; pp. 280–291. [Google Scholar]
- Zhang, X.; Wang, D.; Dongye, M.; Zhu, Y.; Chen, C.; Wang, R.; Long, E.; Liu, Z.; Wu, X.; Lin, D.; et al. Loss-of-function mutations in FREM2 disrupt eye morphogenesis. Exp. Eye Res. 2019, 181, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Banlaki, Z.; Elek, Z.; Nanasi, T.; Szekely, A.; Nemoda, Z.; Sasvari-Szekely, M.; Ronai, Z. Polymorphism in the serotonin receptor 2a (HTR2A) gene as possible predisposal factor for aggressive traits. PLoS ONE 2015, 10, e0117792. [Google Scholar] [CrossRef]
- Nedic Erjavec, G.; Tudor, L.; Nikolac Perkovic, M.; Podobnik, J.; Dodig Curkovic, K.; Curkovic, M.; Svob Strac, D.; Cusek, M.; Bortolato, M.; Pivac, N. Serotonin 5-HT(2A) receptor polymorphisms are associated with irritability and aggression in conduct disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 117, 110542. [Google Scholar] [CrossRef]
- Patton, J.; Brewer, C.; Chien, W.; Johnston, J.J.; Griffith, A.J.; Biesecker, L.G. A genotypic ascertainment approach to refute the association of MYO1A variants with non-syndromic deafness. Eur. J. Hum. Genet. 2016, 25, 147–149. [Google Scholar] [CrossRef]
- Metzger, J.; Rau, J.; Naccache, F.; Bas Conn, L.; Lindgren, G.; Distl, O. Genome data uncover four synergistic key regulators for extremely small body size in horses. BMC Genom. 2018, 19, 492. [Google Scholar] [CrossRef]
- Norgard, E.A.; Lawson, H.A.; Pletscher, L.S.; Wang, B.; Brooks, V.R.; Wolf, J.B.; Cheverud, J.M. Genetic factors and diet affect long-bone length in the F34 LG,SM advanced intercross. Mamm. Genome 2011, 22, 178–196. [Google Scholar] [CrossRef]
- Geng, X.; Liu, S.; Yao, J.; Bao, L.; Zhang, J.; Li, C.; Wang, R.; Sha, J.; Zeng, P.; Zhi, D.; et al. A Genome-Wide Association Study Identifies Multiple Regions Associated with Head Size in Catfish. G3 2016, 6, 3389–3398. [Google Scholar] [CrossRef]
- Van Otterloo, E.; Williams, T.; Artinger, K.B. The old and new face of craniofacial research: How animal models inform human craniofacial genetic and clinical data. Dev. Biol. 2016, 415, 171–187. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | ||||||
|---|---|---|---|---|---|---|
| 5′UTR-194 | 5′UTR-72 | E5 63 | E5 250-253 | |||
| Breed | Coat Phenotype | N | G/A | C/T | C/T | TTTG/del |
| Cornish Rex | Curly | 20 | G | C | C | del |
| Devon Rex | Curly | 4 | G | C/T | C | TTTG/del |
| Selkirk Rex | Curly | 4 | G | C/T | C/T | TTTG |
| Sphynx | Hairless | 1 | G | C | C | TTTG |
| Don Sphynx | Hairless | 4 | G | C | C | TTTG |
| Domestic cat | Straight hair | 3 | G | T | C | TTTG |
| Cornish Rex × Domestic cat F1 | Straight hair | 4 | G | C | C | TTTG/del |
| All | 40 | |||||
| Genomic Position | Number of Animals in ROH | Protein Coding Genes in ROH Islands |
|---|---|---|
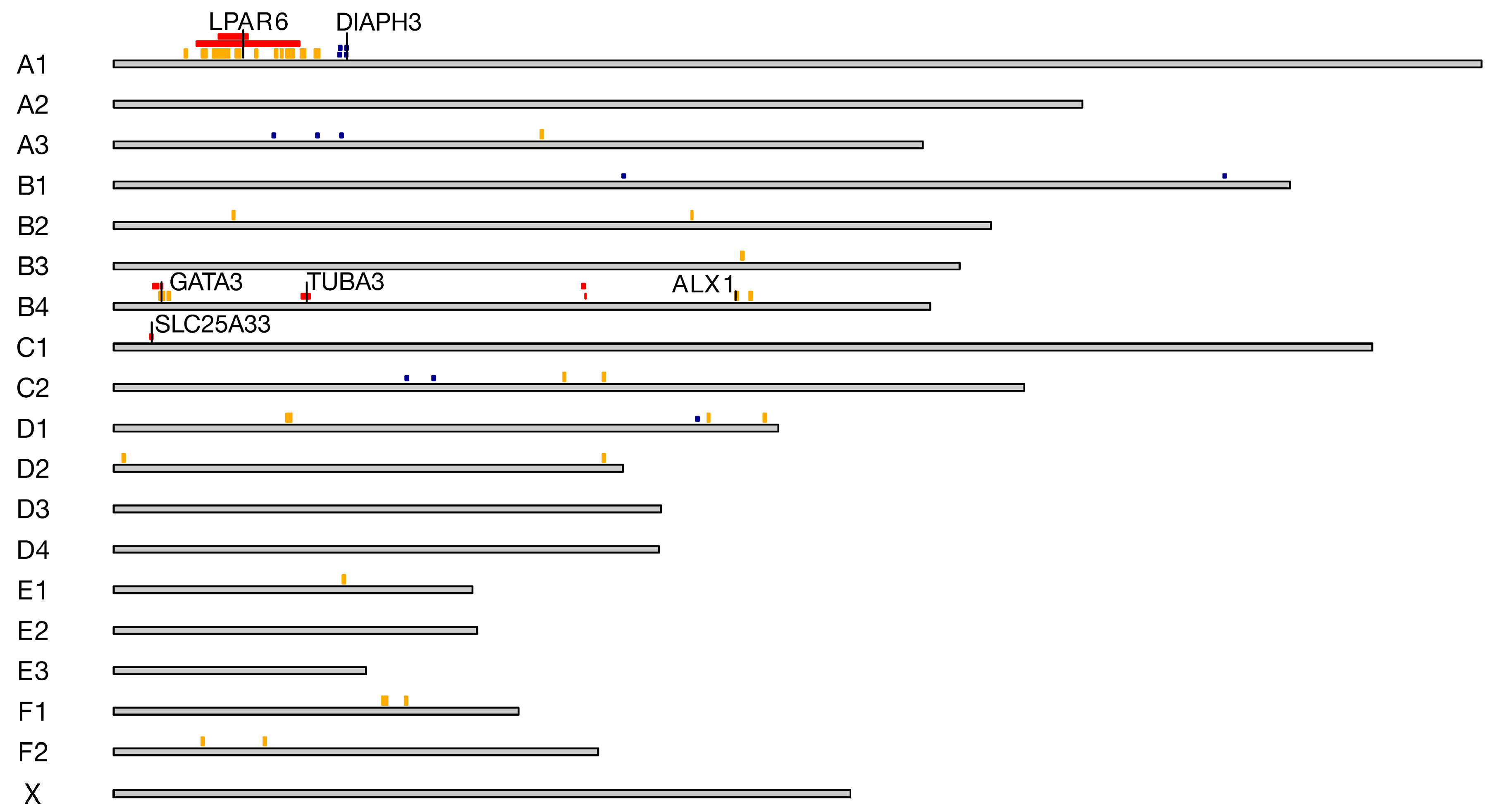
| A1: 18,541,422–23,597,569 | 11 | FOXO1, TRNAR-UCG, MRPS31, SLC25A15, THSD1, VPS36, CKAP2, NEK3, NEK5, ALG11, ATP7B, CCDC70, TMEM272, DHRS12, WDFY2, INTS6, SERPINE3, FAM124A, CA1H13orf42, RNASEH2B, DLEU7, KCNRG, TRIM13, SPRYD7, KPNA3, EBPL, ARL11, RCBTB1, PHF11, SETDB2, CAB39L, CDADC1, MLNR, FNDC3A, CYSLTR2, LOC109497974, RCBTB2, RB1, LPAR6, ITM2B, MED4, NUDT15, SUCLA2 |
| B4: 7,564,954–8,554,345 | 11 | TAF3, GATA3 |
| B4: 82,833,573–83,377,506 | 11 | STAT2, APOF, TIMELESS, MIP, SPRYD4, GLS2, RBMS2, BAZ2A, ATP5F1B, PTGES3, NACA, PRIM1, HSD17B6, SDR9C7, RDH16, GPR182, ZBTB39, TAC3 |
| A1: 14,659,146–32,635,718 | 10 | DCLK1, SOHLH2, CCDC169, SPART, CCNA1, SERTM1, RFXAP, SMAD9, ALG5, EXOSC8, SUPT20H, POSTN, TRPC4, UFM1, FREM2, STOML3, PROSER1, NHLRC3, LHFPL6, COG6, FOXO1, TRNAR-UCG, MRPS31, SLC25A15, THSD1, VPS36, CKAP2, NEK3, NEK5, ALG11, ATP7B, CCDC70, TMEM272, DHRS12, WDFY2, INTS6, SERPINE3, FAM124A, CA1H13orf42, RNASEH2B, DLEU7, KCNRG, TRIM13, SPRYD7, KPNA3, EBPL, ARL11, RCBTB1, PHF11, SETDB2, CAB39L, CDADC1, MLNR, FNDC3A, CYSLTR2, RCBTB2, RB1, LPAR6, ITM2B, MED4, NUDT15, SUCLA2, HTR2A, ESD, LRCH1, RUBCNL, LRRC63, LCP1, CPB2, ZC3H13, CBY2, SIAH3, ERICH6B, COG3, SLC25A30, TPT1, GTF2F2, KCTD4, GPALPP1, NUFIP1, TRNAE-UUC, TSC22D1, SERP2, SMIM2, LACC1, CCDC122, ENOX1, DNAJC15, EPSTI1, FAM216B, TNFSF11, AKAP11, DGKH, VWA8, RGCC, NAA16, MTRF1, KBTBD7, WBP4, ELF1, TRNAE-UUC, SUGT1, CNMD, PCDH8, OLFM4 |
| B4: 8,604,768–8,903,308 | 10 | LOC123386437, LOC123386806 |
| B4: 33,388,421–34,885,172 | 10 | USP18, TUBA8, PEX26, MICAL3, BID, BCL2L13 |
| B4: 83,404,047–83,486,525 | 10 | MYO1A, NAB2, STAT6, LRP1 |
| C1: 6,410,172–6,908,245 | 10 | H6PD, SPSB1, SLC25A33, TMEM201, PIK3CD, CLSTN1 |
| Marker | Chromosome | Position (bp) | iHS | p-Value | Protein Coding |
|---|---|---|---|---|---|
| chrA1.45401576 | A1 | 40,003,327 | −4.28 | 4.74 | DIAPH3, TDRD3, PCDH9 |
| chrA1.45490019 | A1 | 40,077,392 | 4.19 | 4.56 | TDRD3, PCDH9 |
| chrA3.333763 | A3 | 28,335,268 | −4.00 | 4.19 | SNPH, RAD21L1, SDCBP2 |
| chrC2.67784789 | C2 | 56,640,227 | −3.83 | 3.90 | CD47, IFT57, HHLA2, MYH15, DZIP3, MORC1, NECTIN3, CD96, PHLDB2 |
| chrA3.41536320 | A3 | 36,072,528 | −3.83 | 3.89 | SNAP25 |
| chrUn33.3409815 | C2 | 51,884,775 | −3.70 | 3.67 | ZPLD1, ALCAM, CBLB |
| chrA3.46750993 | A3 | 40,332,715 | −3.70 | 3.67 | MACROD2, KIF16B, SEL1L2 |
| chrB1.218769158 | B1 | 196,651,922 | −3.69 | 3.64 | KCNIP4, SLIT2, LDB2, CLNK, SLC2A9, STK32B |
| chrA1.46779148 | A1 | 41,152,849 | 3.60 | 3.50 | DIAPH3, TDRD3 |
| chrA1.46838659 | A1 | 41,215,975 | 3.60 | 3.50 | DIAPH3, TDRD3 |
| chrUn18.8527193 | D1 | 103,331,049 | −3.56 | 3.43 | CNTF, ZFP91, GLYAT, GLYATL2 |
| chrB1.111674326 | B1 | 90,268,797 | −3.55 | 3.41 | ELF2, SCLT1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zorc, M.; Horvat, T.; Tanšek, A.; Ferme, T.; Dovč, P. Selection Signatures Reveal Candidate Genes for the Cornish Rex Breed-Specific Phenotype. Genes 2024, 15, 368. https://doi.org/10.3390/genes15030368
Zorc M, Horvat T, Tanšek A, Ferme T, Dovč P. Selection Signatures Reveal Candidate Genes for the Cornish Rex Breed-Specific Phenotype. Genes. 2024; 15(3):368. https://doi.org/10.3390/genes15030368
Chicago/Turabian StyleZorc, Minja, Tajda Horvat, Anja Tanšek, Tamara Ferme, and Peter Dovč. 2024. "Selection Signatures Reveal Candidate Genes for the Cornish Rex Breed-Specific Phenotype" Genes 15, no. 3: 368. https://doi.org/10.3390/genes15030368