
Cross-Species Comparison of the Pan-RAF Inhibitor LY3009120’s Anti-Tumor Effects in Equine, Canine, and Human Malignant Melanoma Cell Lines
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Culture and Melanoma Cells Samples
2.2. Inhibitor
2.3. Sequencing
2.3.1. DNA, RNA Extraction, and cDNA Synthesis
2.3.2. PCR and Sanger Sequencing
2.3.3. Whole Exome Sequencing
2.4. Cell Counting and Viability Assays
2.4.1. Proliferation
2.4.2. Metabolic Activity
2.4.3. Biomass Quantification
2.5. Early Apoptosis and Late Apoptosis/Necrosis Measurements
2.6. Examination of Cell Morphology Changes
2.7. Statistical Analyses
3. Results
3.1. The Genotypes of BRAF, NRAS, KRAS, and KIT Mutation in Ten Melanoma Cell Lines
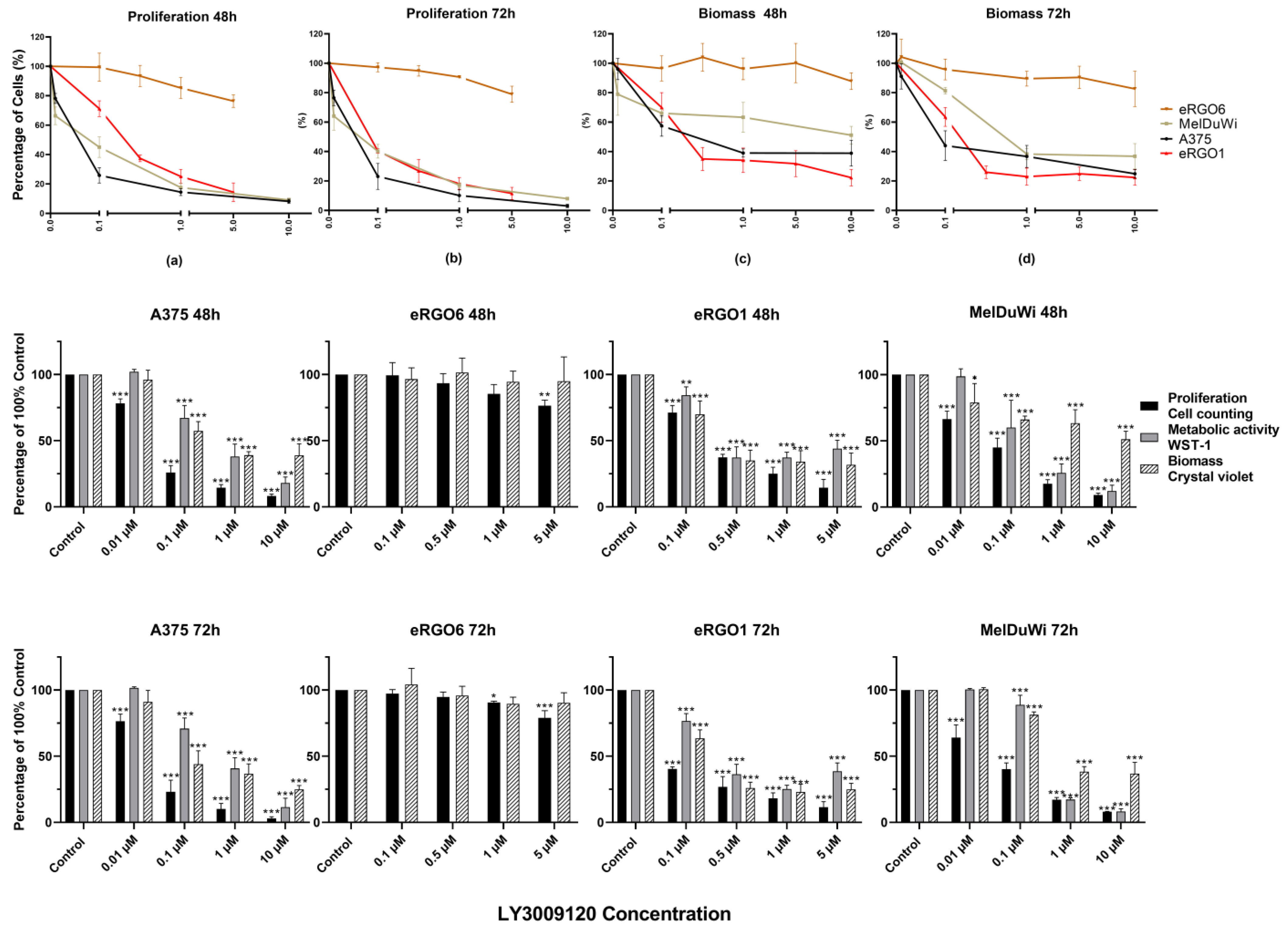
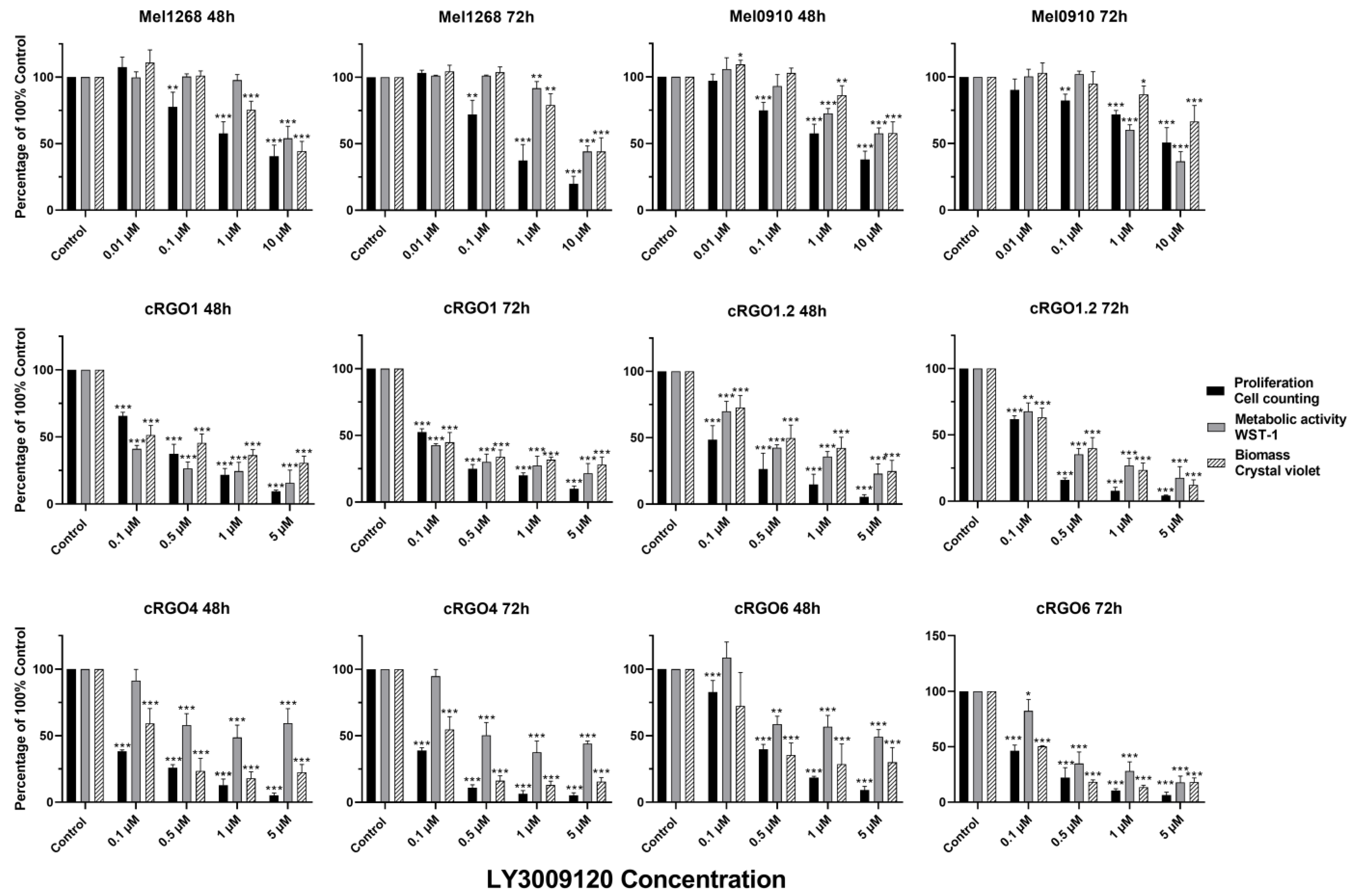
3.2. Application of Pan-RAF Inhibitor (LY3009120)
3.2.1. Effect on Viability and Proliferation
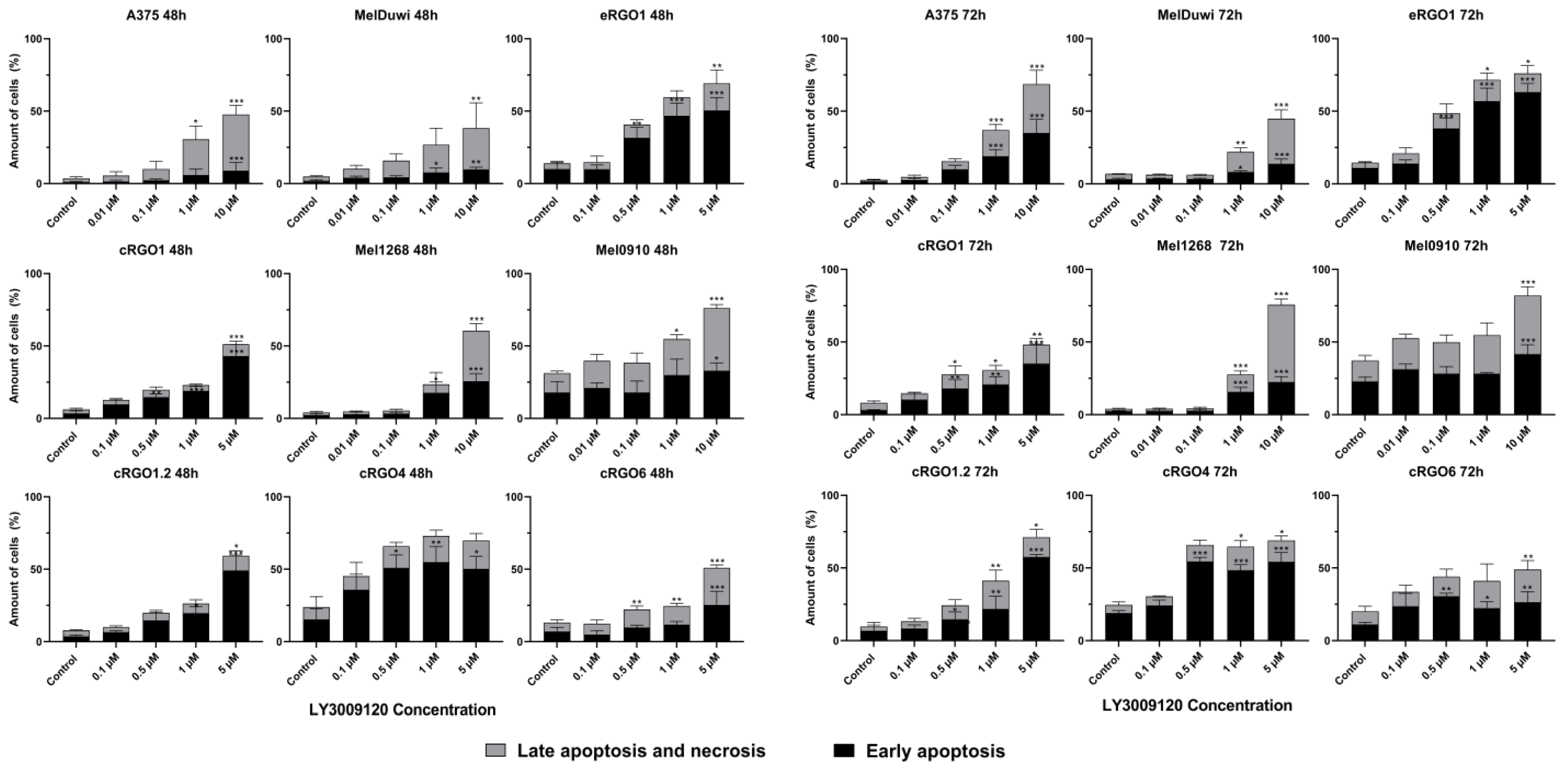
3.2.2. Application of Pan-RAF Inhibitor (LY3009120) Induces Early Apoptosis and Late Apoptosis/Necrosis of Melanoma Cell Lines
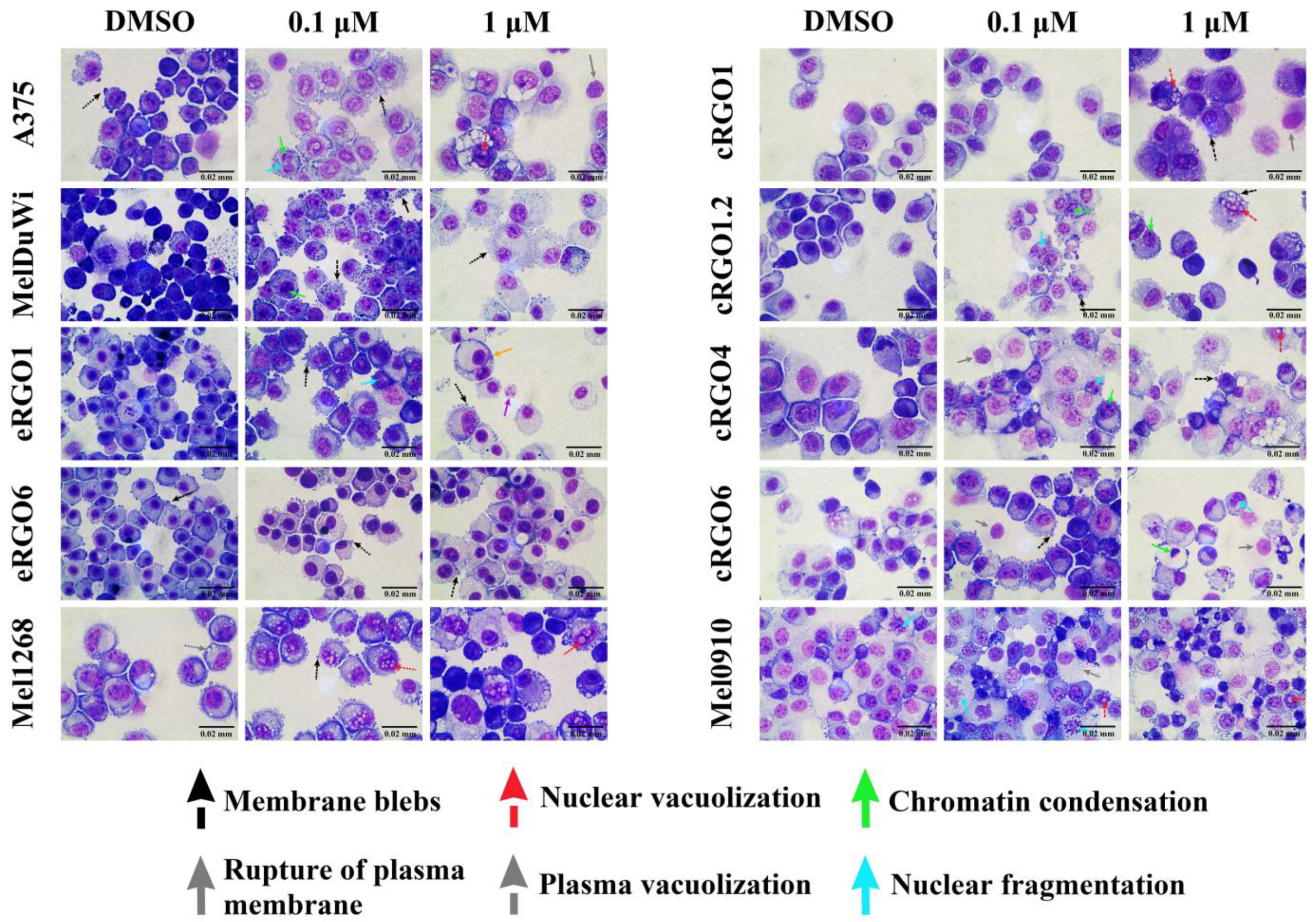
3.2.3. Morphological Changes in Melanoma Cell Lines with Application of Pan-RAF Inhibitor (LY3009120)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, J.H.; Choi, J.W.; Kim, Y.S. Frequencies of BRAF and NRAS mutations are different in histological types and sites of origin of cutaneous melanoma: A meta-analysis. Br. J. Dermatol. 2011, 164, 776–784. [Google Scholar] [CrossRef]
- Hayward, N.K.; Wilmott, J.S.; Waddell, N.; Johansson, P.A.; Field, M.A.; Nones, K.; Patch, A.-M.; Kakavand, H.; Alexandrov, L.B.; Burke, H.; et al. Whole-genome landscapes of major melanoma subtypes. Nature 2017, 545, 175–180. [Google Scholar] [CrossRef]
- Druml, T.; Brem, G.; Horna, M.; Ricard, A.; Grilz-Seger, G. DPF3, A Putative Candidate Gene For Melanoma Etiopathogenesis in Gray Horses. J. Equine Vet. Sci. 2022, 108, 103797. [Google Scholar] [CrossRef]
- Fowles, J.S.; Denton, C.L.; Gustafson, D.L. Comparative analysis of MAPK and PI3K/AKT pathway activation and inhibition in human and canine melanoma. Vet. Comp. Oncol. 2015, 13, 288–304. [Google Scholar] [CrossRef]
- Conrad, D.; Kehl, A.; Beitzinger, C.; Metzler, T.; Steiger, K.; Pfarr, N.; Fischer, K.; Klopfleisch, R.; Aupperle-Lellbach, H. Molecular Genetic Investigation of Digital Melanoma in Dogs. Vet. Sci. 2022, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.M.; Bastian, B.C.; Michael, H.T.; Webster, J.D.; Prasad, M.L.; Conway, C.M.; Prieto, V.M.; Gary, J.M.; Goldschmidt, M.H.; Esplin, D.G.; et al. Sporadic naturally occurring melanoma in dogs as a preclinical model for human melanoma. Pigment. Cell Melanoma Res. 2014, 27, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.; van der Weyden, L.; Schott, C.R.; Foote, A.; Constantino-Casas, F.; Smith, S.; Dobson, J.M.; Murchison, E.P.; Wu, H.; Yeh, I.; et al. Cross-species genomic landscape comparison of human mucosal melanoma with canine oral and equine melanoma. Nat. Commun. 2019, 10, 353. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, W.P.D.; Zismann, V.; Sivaprakasam, K.; Legendre, C.; Poorman, K.; Tembe, W.; Perdigones, N.; Kiefer, J.; Liang, W.; DeLuca, V.; et al. Somatic inactivating PTPRJ mutations and dysregulated pathways identified in canine malignant melanoma by integrated comparative genomic analysis. PLoS Genet. 2018, 14, e1007589. [Google Scholar] [CrossRef]
- Jiang, L.; Campagne, C.; Sundström, E.; Sousa, P.; Imran, S.; Seltenhammer, M.; Pielberg, G.; Olsson, M.J.; Egidy, G.; Andersson, L.; et al. Constitutive activation of the ERK pathway in melanoma and skin melanocytes in Grey horses. BMC Cancer 2014, 14, 857. [Google Scholar] [CrossRef]
- Prouteau, A.; André, C. Canine Melanomas as Models for Human Melanomas: Clinical, Histological, and Genetic Comparison. Genes 2019, 10, 501. [Google Scholar] [CrossRef]
- van der Weyden, L.; Brenn, T.; Patton, E.E.; Wood, G.A.; Adams, D.J. Spontaneously occurring melanoma in animals and their relevance to human melanoma. J. Pathol. 2020, 252, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Bax, M.J.; Brown, M.D.; Rothberg, P.G.; Laughlin, T.S.; Scott, G.A. Pigmented epithelioid melanocytoma (animal-type melanoma): An institutional experience. J. Am. Acad. Dermatol. 2017, 77, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Gao, Y.; Yu, F.; Zhu, Y.; Liu, H.; Li, J.; Escobar, H.M. Interventions for treatment of cutaneous melanoma in horses: A structured literature review. Vet. Res. Commun. 2023, 47, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Luke, J.J.; Flaherty, K.T.; Ribas, A.; Long, G.V. Targeted agents and immunotherapies: Optimizing outcomes in melanoma. Nat. Rev. Clin. Oncol. 2017, 14, 463–482. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wang, H.; Li, C. Signal pathways of melanoma and targeted therapy. Signal Transduct. Target. Ther. 2021, 6, 424. [Google Scholar] [CrossRef]
- Coelho, Y.N.B.; Soldi, L.R.; da Silva, P.H.R.; Mesquita, C.M.; Paranhos, L.R.; dos Santos, T.R.; Silva, M.J.B. Tyrosine kinase inhibitors as an alternative treatment in canine mast cell tumor. Front. Vet. Sci. 2023, 10, 1188795. [Google Scholar] [CrossRef]
- Pimenta, J.; Prada, J.; Cotovio, M. Equine Melanocytic Tumors: A Narrative Review. Animals 2023, 13, 247. [Google Scholar] [CrossRef]
- Bauquier, J.; Tudor, E.; Bailey, S. Effect of the p38 MAPK inhibitor doramapimod on the systemic inflammatory response to intravenous lipopolysaccharide in horses. J. Vet. Intern. Med. 2020, 34, 2109–2116. [Google Scholar] [CrossRef]
- Roskoski, R., Jr. Targeting oncogenic Raf protein-serine/threonine kinases in human cancers. Pharmacol. Res. 2018, 135, 239–258. [Google Scholar] [CrossRef]
- Peng, S.-B.; Henry, J.R.; Kaufman, M.D.; Lu, W.-P.; Smith, B.D.; Vogeti, S.; Rutkoski, T.J.; Wise, S.; Chun, L.; Zhang, Y.; et al. Inhibition of RAF Isoforms and Active Dimers by LY3009120 Leads to Anti-tumor Activities in RAS or BRAF Mutant Cancers. Cancer Cell. 2015, 28, 384–398. [Google Scholar] [CrossRef]
- Mendiratta, G.; Stites, E. Theoretical analysis reveals a role for RAF conformational autoinhibition in paradoxical activation. Elife 2023, 12, e82739. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-H.; Gong, X.; Zhang, Y.; Van Horn, R.D.; Yin, T.; Huber, L.; Burke, T.F.; Manro, J.; Iversen, P.W.; Wu, W.; et al. RAF inhibitor LY3009120 sensitizes RAS or BRAF mutant cancer to CDK4/6 inhibition by abemaciclib via superior inhibition of phospho-RB and suppression of cyclin D1. Oncogene 2018, 37, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Henry, J.R.; Kaufman, M.D.; Peng, S.B.; Ahn, Y.M.; Caldwell, T.M.; Vogeti, L.; Telikepalli, H.; Lu, W.P.; Hood, M.M.; Rutkoski, T.J.; et al. Discovery of 1-(3,3-dimethylbutyl)-3-(2-fluoro-4-methyl-5-(7-methyl-2-(methylamino)pyrido [2,3-d]pyrimidin-6-yl)phenyl)urea (LY3009120) as a pan-RAF inhibitor with minimal paradoxical activation and activity against BRAF or RAS mutant tumor cells. J. Med. Chem. 2015, 58, 4165–4179. [Google Scholar] [CrossRef]
- Chen, S.-H.; Zhang, Y.; Van Horn, R.D.; Yin, T.; Buchanan, S.; Yadav, V.; Mochalkin, I.; Wong, S.S.; Yue, Y.G.; Huber, L.; et al. Oncogenic BRAF Deletions That Function as Homodimers and Are Sensitive to Inhibition by RAF Dimer Inhibitor LY3009120. Cancer Discov. 2016, 6, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Vakana, E.; Pratt, S.; Blosser, W.; Dowless, M.; Simpson, N.; Yuan, X.-J.; Jaken, S.; Manro, J.; Stephens, J.; Zhang, Y.; et al. LY3009120, a panRAF inhibitor, has significant anti-tumor activity in BRAF and KRAS mutant preclinical models of colorectal cancer. Oncotarget 2017, 8, 9251–9266. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, F.; Brodesser, D.; Reifinger, M.; Forte, S.; Semp, P.; Eberspächer-Schweda, M.C.; Wolschek, M.; Brandt, S.; Kleiter, M.; Pratscher, B. Canine oral primary melanoma cells exhibit shift to mesenchymal phenotype and phagocytic behaviour. Vet. Comp. Oncol. 2019, 17, 211–220. [Google Scholar] [CrossRef]
- Weber, L.A.; Meißner, J.; Delarocque, J.; Kalbitz, J.; Feige, K.; Kietzmann, M.; Michaelis, A.; Paschke, R.; Michael, J.; Pratscher, B.; et al. Betulinic acid shows anticancer activity against equine melanoma cells and permeates isolated equine skin in vitro. BMC Vet. Res. 2020, 16, 44. [Google Scholar] [CrossRef]
- Semp, P.-G.; Brandt, S.; Burgener, I.; Kleiter, M.; Pratscher, B. Characterization of radio-sensitivity of canine oral malignant melanoma cell lines and their secretion of cytokines. Veterinary and Comparative Oncology. In Proceedings of the ESVONC Annual Congress, 2021, PP 017, virtual, 28–29 May 2021; 2021; Volume 19. [Google Scholar]
- Oh, J.H.; Cho, J.Y. Comparative oncology: Overcoming human cancer through companion animal studies. Exp. Mol. Med. 2023, 55, 725–734. [Google Scholar] [CrossRef]
- Hatzivassiliou, G.; Song, K.; Yen, I.; Brandhuber, B.J.; Anderson, D.J.; Alvarado, R.; Ludlam, M.J.C.; Stokoe, D.; Gloor, S.L.; Vigers, G.; et al. RAF inhibitors prime wild type RAF to activate the MAPK pathway and enhance growth. Nature 2010, 464, 431–435. [Google Scholar] [CrossRef]
- Jin, T.; Lavoie, H.; Sahmi, M.; David, M.; Hilt, C.; Hammell, A.; Therrien, M. RAF inhibitors promote RAS-RAF interaction by allosterically disrupting RAF autoinhibition. Nat. Commun. 2017, 8, 1211. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. The KRAS crowd targets its next cancer mutations. Nat. Rev. Drug Discov. 2023, 22, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yu, H.; Hughes, N.W.; Liu, B.; Kendirli, A.; Klein, K.; Chen, W.W.; Lander, E.S.; Sabatini, D.M. Gene Essentiality Profiling Reveals Gene Networks and Synthetic Lethal Interactions with Oncogenic Ras. Cell 2017, 168, 890–903.e15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Richmond, A.; Yan, C. Immunomodulatory Properties of PI3K/AKT/mTOR and MAPK/MEK/ERK Inhibition Augment Response to Immune Checkpoint Blockade in Melanoma and Triple-Negative Breast Cancer. Int. J. Mol. Sci. 2022, 23, 7353. [Google Scholar] [CrossRef] [PubMed]
- Lastraioli, E.; Ruffinatti, F.A.; Bagni, G.; Visentin, L.; di Costanzo, F.; Munaron, L.; Arcangeli, A. The Transcriptional Landscape of BRAF Wild Type Metastatic Melanoma: A Pilot Study. Int. J. Mol. Sci. 2022, 23, 6898. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, E.M.; Alhassan, S.A.; Errami, Y.; Elmageed, Z.Y.A.; Fang, J.S.; Wang, G.; Brooks, M.A.; Abi-Rached, J.A.; Kandil, E.; Zerfaoui, M. A Predictive Model of Adaptive Resistance to BRAF/MEK Inhibitors in Melanoma. Int. J. Mol. Sci. 2023, 24, 8407. [Google Scholar] [CrossRef]
- Bergholz, J.S.; Zhao, J.J. How Compensatory Mechanisms and Adaptive Rewiring Have Shaped Our Understanding of Therapeutic Resistance in Cancer. Cancer Res. 2021, 81, 6074–6077. [Google Scholar] [CrossRef]
- Paraiso, K.H.T.; Fedorenko, I.V.; Cantini, L.P.; Munko, A.C.; Hall, M.; Sondak, V.K.; Messina, J.L.; Flaherty, K.T.; Smalley, K.S.M. Recovery of phospho-ERK activity allows melanoma cells to escape from BRAF inhibitor therapy. Br. J. Cancer 2010, 102, 1724–1730. [Google Scholar] [CrossRef]
- Atefi, M.; Titz, B.; Avramis, E.; Ng, C.; Wong, D.J.; Lassen, A.; Cerniglia, M.; Escuin-Ordinas, H.; Foulad, D.; Comin-Anduix, B.; et al. Combination of pan-RAF and MEK inhibitors in NRAS mutant melanoma. Mol. Cancer 2015, 14, 27. [Google Scholar] [CrossRef]
- Fischer, G.M.; Vashisht Gopal, Y.N.; McQuade, J.L.; Peng, W.; DeBerardinis, R.J.; Davies, M.A. Metabolic strategies of melanoma cells: Mechanisms, interactions with the tumor microenvironment, and therapeutic implications. Pigment. Cell Melanoma Res. 2018, 31, 11–30. [Google Scholar] [CrossRef]
- Mayers, J.R.; Torrence, M.E.; Danai, L.V.; Papagiannakopoulos, T.; Davidson, S.M.; Bauer, M.R.; Lau, A.N.; Ji, B.W.; Dixit, P.D.; Hosios, A.M.; et al. Tissue of origin dictates branched-chain amino acid metabolism in mutant Kras-driven cancers. Science 2016, 353, 1161–1165. [Google Scholar] [CrossRef]
- Amendola, C.R.; Mahaffey, J.P.; Parker, S.J.; Ahearn, I.M.; Chen, W.-C.; Zhou, M.; Court, H.; Shi, J.; Mendoza, S.L.; Morten, M.J.; et al. KRAS4A directly regulates hexokinase 1. Nature 2019, 576, 482–486. [Google Scholar] [CrossRef]
- Hunter, J.C.; Manandhar, A.; Carrasco, M.A.; Gurbani, D.; Gondi, S.; Westover, K.D. Biochemical and Structural Analysis of Common Cancer-Associated KRAS Mutations. Mol. Cancer Res. 2015, 13, 1325–1335. [Google Scholar] [CrossRef]
- Wei, B.R.; Michael, H.T.; Halsey, C.H.C.; Peer, C.J.; Adhikari, A.; Dwyer, J.E.; Hoover, S.B.; El Meskini, R.; Kozlov, S.; Ohler, Z.W.; et al. Synergistic targeted inhibition of MEK and dual PI3K/mTOR diminishes viability and inhibits tumor growth of canine melanoma underscoring its utility as a preclinical model for human mucosal melanoma. Pigment Cell Melanoma Res. 2016, 29, 643–655. [Google Scholar] [CrossRef]
- Mayr, B.; Schaffner, G.; Reifinger, M.; Zwetkoff, S.; Prodinger, B. N-ras mutations in canine malignant melanomas. Vet J. 2003, 165, 169–171. [Google Scholar] [CrossRef]
- Murua Escobar, H.; Günther, K.; Richter, A.; Soller, J.T.; Winkler, S.; Nolte, I.; Bullerdiek, J. Absence of ras-gene hot-spot mutations in canine fibrosarcomas and melanomas. Anticancer Res. 2004, 24, 3027–3028. [Google Scholar]
- Shelly, S.; Chien, M.B.; Yip, B.; Kent, M.S.; Theon, A.P.; McCallan, J.L.; London, C.A. Exon 15 BRAF mutations are uncommon in canine oral malignant melanomas. Mamm Genome. 2005, 16, 211–217. [Google Scholar] [CrossRef]
- Mochizuki, H.; Kennedy, K.; Shapiro, S.G.; Breen, M. BRAF Mutations in Canine Cancers. PLoS ONE 2015, 10, e0129534. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Network. Genomic Classification of Cutaneous Melanoma. Cell 2015, 161, 1681–1696. [Google Scholar] [CrossRef]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Rutkowski, P.; Lao, C.D.; Cowey, C.L.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Five-Year Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2019, 381, 1535–1546. [Google Scholar] [CrossRef]
- Sundström, E.; Imsland, F.; Mikko, S.; Wade, C.; Sigurdsson, S.; Pielberg, G.R.; Golovko, A.; Curik, I.; Seltenhammer, M.H.; Sölkner, J.; et al. Copy number expansion of the STX17 duplication in melanoma tissue from Grey horses. BMC Genom. 2012, 13, 365. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Cell Line | Breed | Age (y) | Sex | IHC | Diagnosis | Location | Tumor Type |
|---|---|---|---|---|---|---|---|---|
| Canine | ||||||||
| 1 | Mel1268 | Mix | 10 | f | _ | OMM | Oral cavity | Primary MM |
| 2 | Mel0910 | Yorkshire Terrier | 15 | f | _ | Cutaneous MM | Ear | Primary MM |
| 3 | cRGO1 | Mix | 11 | f | Melan-A +, PNL2 +, CD146 | Malignant predominantly amelanotic melanoma | Back-end hard palate | Primary OMM |
| 4 | cRGO1.2 * | Melan-A +, S100 -, Vimentin ++, MMP2 +++, MMP9 + | Left mandibular LN | LN metastasis | ||||
| 5 | cRGO4 | Golden Retriever | 9 | m | Melan-A +, PNL2 +, CD146 | OMM | Right mandible | Primary MM |
| 6 | cRGO6 | Weimaraner | 10 | f | HMB45 -, PNL2 ++ | Amelanotic OMM (III (T2N1M0)) | Multiple skin lesions | Cutaneous metastasis |
| Equine | ||||||||
| 7 | MelDuWi | Andalusian Grey horse | 15 | m | _ | Cutaneous EMM | Skin, lip, eye, penis, and anus | Primary EMM |
| 8 | eRGO1 | Icelandic horse | 16 | m | _ | Multiple cutaneous EMM | Under the tail | Primary EMM |
| 9 | eRGO6 | Grey horse | 12 | f | _ | Metastasizing melanoma | Liver | Liver metastasis |
| Species | Gene | Primer Sequences (5′-3′) | Template | Fragment | Tm [°C] | Ta [°C] | Transcript | |
|---|---|---|---|---|---|---|---|---|
| Canine a | BRAF exon 15 | F | cacgccaagtcaatcatccacaga | cDNA | 228 bp | 62.7 | 55.3 | BRAF-202 ENSCAFT00000006306.4 |
| R | cccaaatgcgtatacatctgactgg | 63.0 | ||||||
| NRAS exon 2 | F | agcttgaggttcttgctggtgtga | cDNA | 474 bp | 62.7 | 59.3 | NRAS-202 | |
| ENSCAFT00845046556.1 | ||||||||
| R | tgtctggtcttggctgaggtttca | 62.7 | ||||||
| Equine b | BRAF exon 11 | F | tccctttcaggcatagggta | gDNA | 247 bp | 57.3 | 53.9 | BRAF-206 ENSECAT00000005257.4 |
| R | tgacatgtgacaaggtcattgtat | 57.6 | ||||||
| BRAF exon 15 | F | tcataatgcttgctctgataagaaa | gDNA | 236 bp | 56.4 | 59.3 | BRAF-206 ENSECAT00000005257.4 | |
| R | cagcatctcagggtccaaa | 56.7 | ||||||
| NRAS exon 2 | F | gtactgtagatgtggctcgc | gDNA | 224 bp | 59.4 | 62.3 | NRAS-201 ENSECAT00000014647.2 | |
| R | acggaagaaagagaggtgga | 57.3 | ||||||
| KRAS exon 2 | F | ccagactgtgtttctcccttc | gDNA | 249 bp | 59.8 | 61.1 | KRAS-203 ENSECAT00000020767.2 | |
| R | caattactcctccatgtcaattt | 55.3 | ||||||
| Mutation Genotypes | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Human | Equine | Canine | ||||||||
| Gene | A375 | MelDuWi | eRGO1 | eRGO6 | Mel1268 | Mel0910 | cRGO1 | cRGO1.2 | cRGO4 | cRGO6 |
| BRAF exon 15 | p.V600E | - | - | - | - | - | - | - | - | - |
| BRAF exon 11 | - | - | - | - | - | - | - | - | - | - |
| NRAS exon 2 | - | - | - | - | - | - | p.G13R | p.G13R | - | - |
| NRAS exon 3 | - | - | - | - | - | - | - | - | - | - |
| KRAS exon 2 | - | p.Q61H | - | - | - | - | - | - | - | - |
| KIT exon 11 | - | - | - | - | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Packeiser, E.-M.; Wendt, S.; Sekora, A.; Cavalleri, J.-M.V.; Pratscher, B.; Alammar, M.; Hühns, M.; Brenig, B.; Junghanss, C.; et al. Cross-Species Comparison of the Pan-RAF Inhibitor LY3009120’s Anti-Tumor Effects in Equine, Canine, and Human Malignant Melanoma Cell Lines. Genes 2024, 15, 202. https://doi.org/10.3390/genes15020202
Gao Y, Packeiser E-M, Wendt S, Sekora A, Cavalleri J-MV, Pratscher B, Alammar M, Hühns M, Brenig B, Junghanss C, et al. Cross-Species Comparison of the Pan-RAF Inhibitor LY3009120’s Anti-Tumor Effects in Equine, Canine, and Human Malignant Melanoma Cell Lines. Genes. 2024; 15(2):202. https://doi.org/10.3390/genes15020202
Chicago/Turabian StyleGao, Yu, Eva-Maria Packeiser, Sophia Wendt, Anett Sekora, Jessika-Maximiliane V. Cavalleri, Barbara Pratscher, Moosheer Alammar, Maja Hühns, Bertram Brenig, Christian Junghanss, and et al. 2024. "Cross-Species Comparison of the Pan-RAF Inhibitor LY3009120’s Anti-Tumor Effects in Equine, Canine, and Human Malignant Melanoma Cell Lines" Genes 15, no. 2: 202. https://doi.org/10.3390/genes15020202