
A Single-Cell Transcriptome of Bovine Milk Somatic Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bovine Milk Samples and Isolation of Cells
2.2. Single-Cell Library Preparation and Sequencing
2.3. Pre-Processing and Quality Control of scRNA-Seq Data
2.4. Identification of Cell Clusters
2.5. Annotation of Cell Types
2.6. Identification of Highly Variable Expressed Genes
3. Results
3.1. ScRNA-Seq Identifies a Diversity of Cell Types in Bovine Milk Somatic Cells
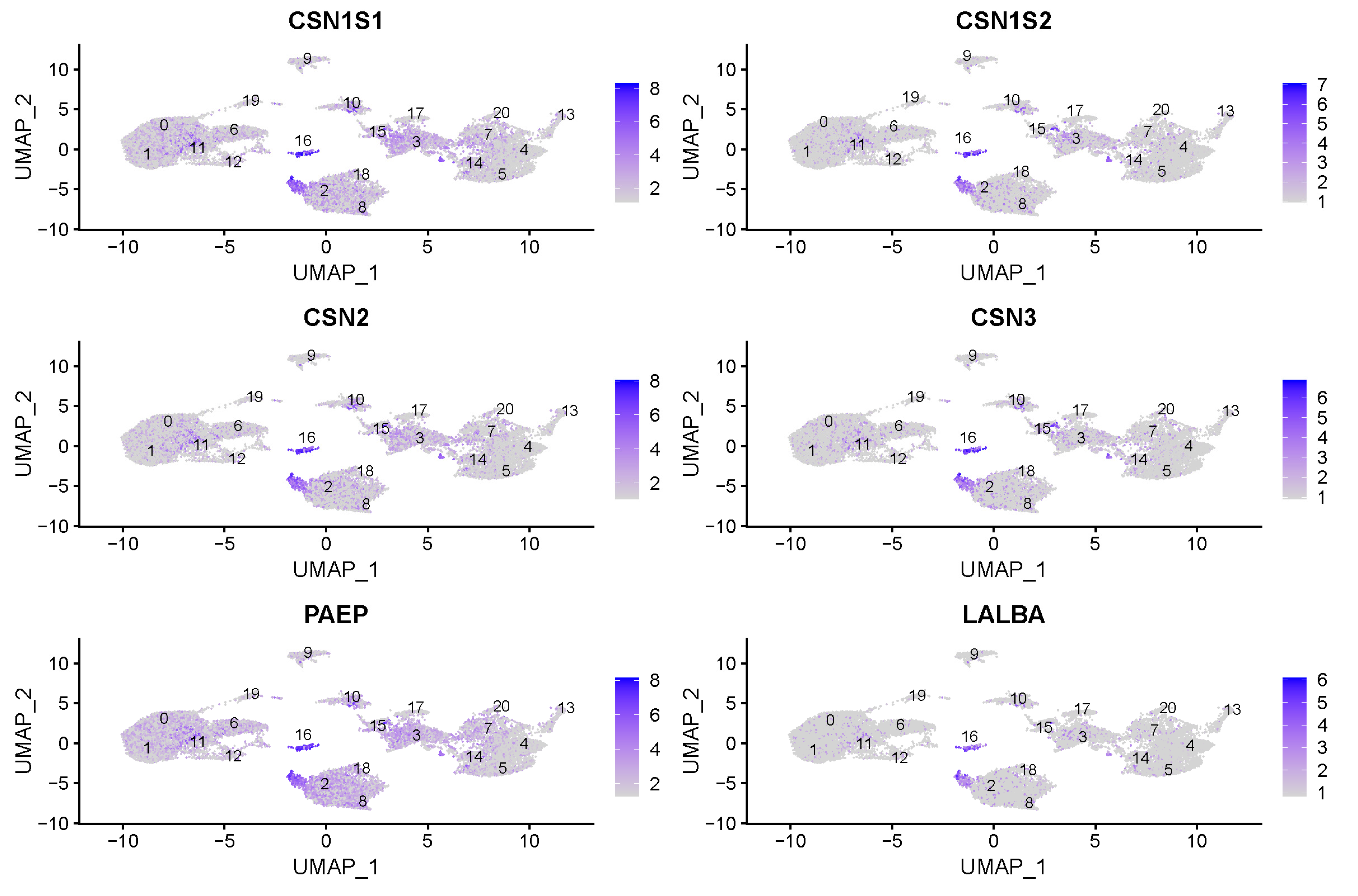
3.2. Milk Producing Cell Clusters
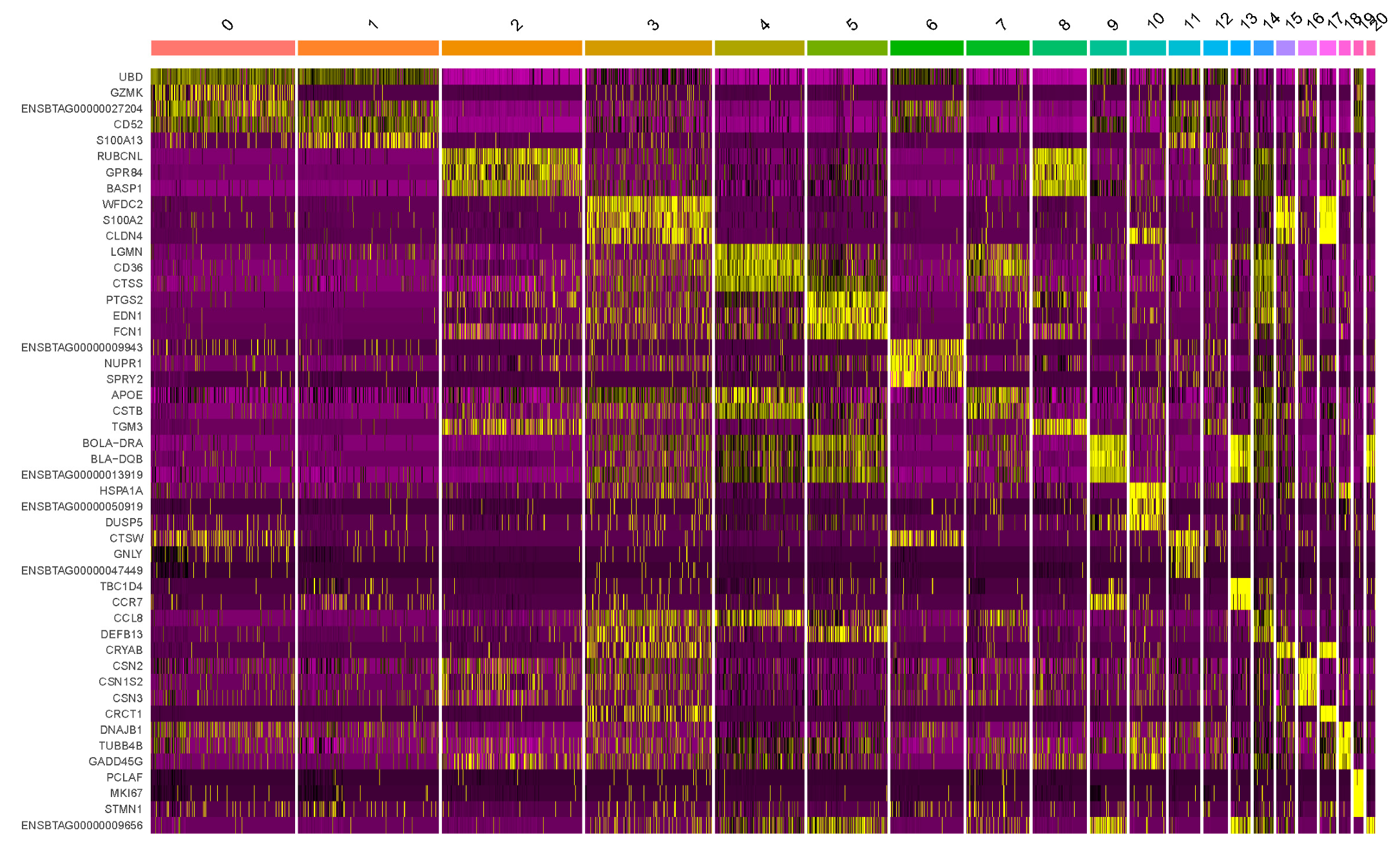
3.3. Highly Variable Expressed Genes in Bovine Milk Somatic Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inman, J.L.; Robertson, C.; Mott, J.D.; Bissell, M.J. Mammary gland development: Cell fate specification, stem cells and the microenvironment. Development 2015, 142, 1028–1042. [Google Scholar] [CrossRef]
- Neville, M.C.; McFadden, T.B.; Forsyth, I. Hormonal regulation of mammary differentiation and milk secretion. J. Mammary Gland. Biol. Neoplasia 2002, 7, 49–66. [Google Scholar] [CrossRef]
- Boutinaud, M.; Jammes, H. Potential uses of milk epithelial cells: A review. Reprod. Nutr. Dev. 2002, 42, 133–147. [Google Scholar] [CrossRef]
- Wall, S.K.; Wellnitz, O.; Bruckmaier, R.M.; Schwarz, D. Differential somatic cell count in milk before, during, and after lipopolysaccharide- and lipoteichoic-acid-induced mastitis in dairy cows. J. Dairy. Sci. 2018, 101, 5362–5373. [Google Scholar] [CrossRef]
- Biteau, B.; Hochmuth, C.E.; Jasper, H. Maintaining tissue homeostasis: Dynamic control of somatic stem cell activity. Cell Stem Cell 2011, 9, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Visvader, J.E.; Stingl, J. Mammary stem cells and the differentiation hierarchy: Current status and perspectives. Genes. Dev. 2014, 28, 1143–1158. [Google Scholar] [CrossRef] [PubMed]
- Faulkin, L.J., Jr.; Deome, K.B. Regulation of growth and spacing of gland elements in the mammary fat pad of the C3H mouse. J. Natl. Cancer Inst. 1960, 24, 953–969. [Google Scholar] [PubMed]
- Deome, K.B.; Faulkin, L.J., Jr.; Bern, H.A.; Blair, P.B. Development of mammary tumors from hyperplastic alveolar nodules transplanted into gland-free mammary fat pads of female C3H mice. Cancer Res. 1959, 19, 515–520. [Google Scholar] [PubMed]
- Shackleton, M.; Vaillant, F.; Simpson, K.J.; Stingl, J.; Smyth, G.K.; Asselin-Labat, M.L.; Wu, L.; Lindeman, G.J.; Visvader, J.E. Generation of a functional mammary gland from a single stem cell. Nature 2006, 439, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Canovas, A.; Rincon, G.; Bevilacqua, C.; Islas-Trejo, A.; Brenaut, P.; Hovey, R.C.; Boutinaud, M.; Morgenthaler, C.; VanKlompenberg, M.K.; Martin, P.; et al. Comparison of five different RNA sources to examine the lactating bovine mammary gland transcriptome using RNA-Sequencing. Sci. Rep. 2014, 4, 5297. [Google Scholar] [CrossRef] [PubMed]
- Maningat, P.D.; Sen, P.; Rijnkels, M.; Sunehag, A.L.; Hadsell, D.L.; Bray, M.; Haymond, M.W. Gene expression in the human mammary epithelium during lactation: The milk fat globule transcriptome. Physiol. Genom. 2009, 37, 12–22. [Google Scholar] [CrossRef]
- Medrano, J.F.; Rincon, G.; Islas-Trejo, A. Comparative analysis of bovine milk and mammary gland transcriptome using RNA-Seq. In Proceedings of the 9th World Congress on Genetics applied to Livestock Production, Leipzig, Germany, 1–6 August 2010. [Google Scholar]
- Wickramasinghe, S.; Rincon, G.; Islas-Trejo, A.; Medrano, J.F. Transcriptional profiling of bovine milk using RNA sequencing. BMC Genom. 2012, 13, 45. [Google Scholar] [CrossRef]
- Tang, F.; Barbacioru, C.; Wang, Y.; Nordman, E.; Lee, C.; Xu, N.; Wang, X.; Bodeau, J.; Tuch, B.B.; Siddiqui, A.; et al. mRNA-Seq whole-transcriptome analysis of a single cell. Nat. Methods 2009, 6, 377–382. [Google Scholar] [CrossRef]
- Becker, D.; Weikard, R.; Hadlich, F.; Kühn, C. Single-cell RNA sequencing of freshly isolated bovine milk cells and cultured primary mammary epithelial cells. Sci. Data 2021, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Twigger, A.-J.; Engelbrecht, L.K.; Bach, K.; Schultz-Pernice, I.; Pensa, S.; Stenning, J.; Petricca, S.; Scheel, C.H.; Khaled, W.T. Transcriptional changes in the mammary gland during lactation revealed by single cell sequencing of cells from human milk. Nat. Commun. 2022, 13, 562. [Google Scholar] [CrossRef]
- Cristea, S.; Polyak, K. Dissecting the mammary gland one cell at a time. Nat. Commun. 2018, 9, 2473. [Google Scholar] [CrossRef] [PubMed]
- Pal, B.; Chen, Y.; Vaillant, F.; Jamieson, P.; Gordon, L.; Rios, A.C.; Wilcox, S.; Fu, N.; Liu, K.H.; Jackling, F.C.; et al. Construction of developmental lineage relationships in the mouse mammary gland by single-cell RNA profiling. Nat. Commun. 2017, 8, 1627. [Google Scholar] [CrossRef]
- Bach, K.; Pensa, S.; Grzelak, M.; Hadfield, J.; Adams, D.J.; Marioni, J.C.; Khaled, W.T. Differentiation dynamics of mammary epithelial cells revealed by single-cell RNA sequencing. Nat. Commun. 2017, 8, 2128. [Google Scholar] [CrossRef]
- Nguyen, Q.H.; Pervolarakis, N.; Blake, K.; Ma, D.; Davis, R.T.; James, N.; Phung, A.T.; Willey, E.; Kumar, R.; Jabart, E.; et al. Profiling human breast epithelial cells using single cell RNA sequencing identifies cell diversity. Nat. Commun. 2018, 9, 2028. [Google Scholar] [CrossRef] [PubMed]
- Giraddi, R.R.; Chung, C.Y.; Heinz, R.E.; Balcioglu, O.; Novotny, M.; Trejo, C.L.; Dravis, C.; Hagos, B.M.; Mehrabad, E.M.; Rodewald, L.W.; et al. Single-Cell Transcriptomes Distinguish Stem Cell State Changes and Lineage Specification Programs in Early Mammary Gland Development. Cell Rep. 2018, 24, 1653–1666.e7. [Google Scholar] [CrossRef]
- Sun, H.; Miao, Z.; Zhang, X.; Chan, U.I.; Su, S.M.; Guo, S.; Wong, C.K.H.; Xu, X.; Deng, C.X. Single-cell RNA-Seq reveals cell heterogeneity and hierarchy within mouse mammary epithelia. J. Biol. Chem. 2018, 293, 8315–8329. [Google Scholar] [CrossRef] [PubMed]
- Wuidart, A.; Sifrim, A.; Fioramonti, M.; Matsumura, S.; Brisebarre, A.; Brown, D.; Centonze, A.; Dannau, A.; Dubois, C.; Van Keymeulen, A.; et al. Early lineage segregation of multipotent embryonic mammary gland progenitors. Nat. Cell Biol. 2018, 20, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Wang, R.; Zhou, Y.; Fei, L.; Sun, H.; Lai, S.; Saadatpour, A.; Zhou, Z.; Chen, H.; Ye, F.; et al. Mapping the Mouse Cell Atlas by Microwell-Seq. Cell 2018, 172, 1091–1107.e17. [Google Scholar] [CrossRef] [PubMed]
- Schaum, N.; Karkanias, J.; Neff, N.F.; May, A.P.; Quake, S.R.; Wyss-Coray, T.; Darmanis, S.; Batson, J.; Botvinnik, O.; Chen, M.B.; et al. Single-cell transcriptomics of 20 mouse organs creates a Tabula Muris. Nature 2018, 562, 367–372. [Google Scholar] [CrossRef]
- Dallas, D.C.; Murray, N.M.; Gan, J. Proteolytic Systems in Milk: Perspectives on the Evolutionary Function within the Mammary Gland and the Infant. J. Mammary Gland. Biol. Neoplasia 2015, 20, 133–147. [Google Scholar] [CrossRef]
- Bionaz, M.; Periasamy, K.; Rodriguez-Zas, S.L.; Everts, R.E.; Lewin, H.A.; Hurley, W.L.; Loor, J.J. Old and new stories: Revelations from functional analysis of the bovine mammary transcriptome during the lactation cycle. PLoS ONE 2012, 7, e33268. [Google Scholar] [CrossRef]
- Oakes, S.R.; Hilton, H.N.; Ormandy, C.J. Key stages in mammary gland development—The alveolar switch: Coordinating the proliferative cues and cell fate decisions that drive the formation of lobuloalveoli from ductal epithelium. Breast Cancer Res. 2006, 8, 207. [Google Scholar] [CrossRef]
- Chromium Next GEM Single Cell 3′ Reagent Kits v3.1. Available online: https://cdn.10xgenomics.com/image/upload/v1660261285/support-documents/CG000204_ChromiumNextGEMSingleCell3_v3.1_Rev_D.pdf (accessed on 15 January 2020).
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411. [Google Scholar] [CrossRef]
- Stuart, T.; Butler, A.; Hoffman, P.; Hafemeister, C.; Papalexi, E.; Mauck, W.M.; Stoeckius, M.; Smibert, P.; Satija, R. Comprehensive integration of single cell data. bioRxiv 2018. [Google Scholar] [CrossRef] [PubMed]
- Ianevski, A.; Giri, A.K.; Aittokallio, T. Fully-automated and ultra-fast cell-type identification using specific marker combinations from single-cell transcriptomic data. Nat. Commun. 2022, 13, 1246. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.-L.; Huang, X.-L.; Qin, W.-Y.; Liu, K.-B.; Wang, Y.; Li, M.; Ren, Y.-H.; Li, Y.-Z.; Sun, Y.-M. singleCellBase: A high-quality manually curated database of cell markers for single cell annotation across multiple species. Biomark. Res. 2023, 11, 83. [Google Scholar] [CrossRef]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, S.; Hua, S.; Rincon, G.; Islas-Trejo, A.; German, J.B.; Lebrilla, C.B.; Medrano, J.F. Transcriptome Profiling of Bovine Milk Oligosaccharide Metabolism Genes Using RNA-Sequencing. PLoS ONE 2011, 6, e18895. [Google Scholar] [CrossRef] [PubMed]
- Yip, S.H.; Sham, P.C.; Wang, J. Evaluation of tools for highly variable gene discovery from single-cell RNA-seq data. Brief. Bioinform. 2019, 20, 1583–1589. [Google Scholar] [CrossRef]
- Marenholz, I.; Zirra, M.; Fischer, D.F.; Backendorf, C.; Ziegler, A.; Mischke, D. Identification of human epidermal differentiation complex (EDC)-encoded genes by subtractive hybridization of entire YACs to a gridded keratinocyte cDNA library. Genome Res. 2001, 11, 341–355. [Google Scholar] [CrossRef]
- Alonso-Fauste, I.; Andrés, M.; Iturralde, M.; Lampreave, F.; Gallart, J.; Álava, M.A. Proteomic characterization by 2-DE in bovine serum and whey from healthy and mastitis affected farm animals. J. Proteom. 2012, 75, 3015–3030. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Boeren, S.; van Hooijdonk, A.C.M.; Vervoort, J.M.; Hettinga, K.A. A proteomic perspective on the changes in milk proteins due to high somatic cell count. J. Dairy Sci. 2015, 98, 5339–5351. [Google Scholar] [CrossRef]
- Boutinaud, M.; Rulquin, H.; Keisler, D.H.; Djiane, J.; Jammes, H. Use of somatic cells from goat milk for dynamic studies of gene expression in the mammary gland. J. Anim. Sci. 2002, 80, 1258–1269. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster | Cluster Annotation | Marker Gene |
|---|---|---|
| 0 | CD8+ T cells | GZMK, ENSBTAG00000027204, CD52, ENSBTAG00000010828, FAM162A, RGS1, CCL5, ENSBTAG00000034609, ENSBTAG00000000432 |
| 1 | CD4+ T cells | UBD, ICOS, ENSBTAG00000055140, GUCY1B1, ENSBTAG00000027204, CD4, S100A13, NCR3, ENSBTAG00000034609 |
| 2 | Neutrophils | GPR84, BASP1, PLAU, PLEK, MARCKS, IL1B, DMXL2, IL1RN, BATF3 |
| 3 | Progenitor cells | CCL2, APOE, S100A2, CLDN4, CD9, CCL8, CST6, CRYAB, CRCT1 |
| 4 | Monocytes | CD36, CTSS, CTSB, LIPA, CCL8, GRN, CNDP2, TREM2, CD9 |
| 5 | Monocytes | EDN1, FCN1, LYZ, CD14, ARAF, TNF, CXCL3, BOLA-DRA, DEFB13 |
| 6 | Mast cells | NUPR1, KIT, ENSBTAG00000055197, SPRY2, CTSW, ENSBTAG00000000144, CD7, TNFRSF9, ENSBTAG00000034609 |
| 7 | Macrophages | FABP5, CD36, CTSB, APOE, CSTB, CNDP2, CTSZ, ATOX1, CD9 |
| 8 | Neutrophils | BASP1, ENSBTAG00000048980 (Chemokine interleukin-8-like domain-containing protein), IFITM3, CXCR1, GPR84, SELL, ENSBTAG00000034366, TGM3, S100A9 |
| 9 | B cells | BLA-DQB, ENSBTAG00000013919, CD74, MS4A1, ENSBTAG00000055240, TNFRSF13C, CCR7, ENSBTAG00000009656, IRF4 |
| 10 | Progenitor cells | TACSTD2, RASD1, ENSBTAG00000050919, DUSP5, EFNB2, ARC, KLF4, HSPA2, MAFB |
| 11 | NK cells | GNLY, ENSBTAG00000047449 (Saposin B-type domain-containing protein), CD52, UBD, S100A13, PRF1, GPR183, ENSBTAG00000000144 (Ig-like domain-containing protein), ENSBTAG00000055197 (Immunoglobulin C1-set domain-containing protein) |
| 12 | Memory CD8+ T cells | RUBCNL, UBD, CD52, BASP1, PLAU, ENSBTAG00000027204, PLEK, IL1RN, ENSBTAG00000034609 |
| 13 | Dendritic cells | CCR7, GPR183, LY75, BLA-DQB, TAMALIN, PKIB, ENSBTAG00000013919, BOLA-DRA, FSCN1 |
| 14 | Monocytes | PTGS2, CCL2, CD36, CCL8, CTSS, RUBCNL, EDN1, CXCL5, DEFB13 |
| 15 | Luminal progenitor cells | CLU, CLDN3, CLDN4, CRYAB, KRT7, DSTN, WFDC2, KRT19, LTF |
| 16 | Luminal cells | CSN1S1, PAEP, CSN1S2, CSN3, GLYCAM1, LALBA, HSTN, SCGB1D, FABP3 |
| 17 | Luminal progenitor cells | CRCT1, AGPAT2, CLDN3, KRT7, CLDN4, DSTN, S100A2, CRYAB, WFDC2 |
| 18 | Neutrophils | TUBB4B, GADD45G, DDIT4, GADD45A, HSPH1, HSPA1A, IER5L, ZFAND2A, LRIF1 |
| 19 | CD4+ CD8+ T cells | PCLAF, MKI67, STMN1, TOP2A, DUT, HMGB2, CENPF, TMPO, DNMT1, UBE2C |
| 20 | Dendritic cells | BOLA-DRA, ENSBTAG00000013919 (BOLA-DRB3), ENSBTAG00000009656 (BOLA-DQA2), CD74, C3H1orf54, BLA-DQB, CST3, ENSBTAG00000037605 (BOLA-DQA1), BOLA-DMA, PLAC8A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zorc, M.; Dolinar, M.; Dovč, P. A Single-Cell Transcriptome of Bovine Milk Somatic Cells. Genes 2024, 15, 349. https://doi.org/10.3390/genes15030349
Zorc M, Dolinar M, Dovč P. A Single-Cell Transcriptome of Bovine Milk Somatic Cells. Genes. 2024; 15(3):349. https://doi.org/10.3390/genes15030349
Chicago/Turabian StyleZorc, Minja, Mateja Dolinar, and Peter Dovč. 2024. "A Single-Cell Transcriptome of Bovine Milk Somatic Cells" Genes 15, no. 3: 349. https://doi.org/10.3390/genes15030349