
Exploring the Regulation and Function of Rpl3l in the Development of Early-Onset Dilated Cardiomyopathy and Congestive Heart Failure Using Systems Genetics Approach
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse BXD Family and Heart Gene Expression Data
2.2. Array Profiling and Data Analysis
2.3. Correlation Analysis
2.4. Expression Quantitative Trait Locus (eQTL) Mapping
2.5. Candidate Gene Selection
2.6. RPL3L Expression in Human and Mouse
2.7. Differential Expression between Human Cardiomyopathy and Control Samples
2.8. Functional Enrichment Analysis
2.9. Immune Cell Infiltration Analysis
3. Results
3.1. Expression of RPL3L in Human and Mouse Heart
3.2. Phenotypes and Pathways Associated with Rpl3l-Correlated Genes
3.3. Differentially Expressed Genes and Functions between Human Cardiomyopathy and Control Samples
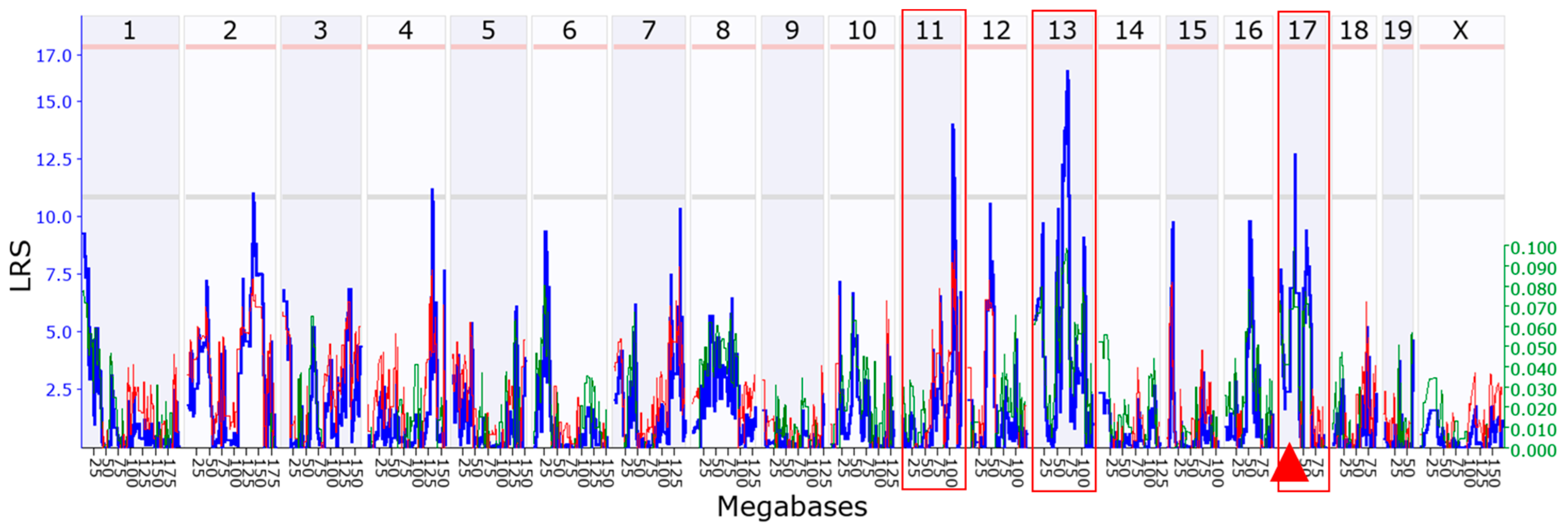
3.4. Chromosomal Loci in a Mouse Genome Regulating Rpl3l Expression
3.5. Identification of the Candidate Genes That Strongly Regulate Rpl3l Expression
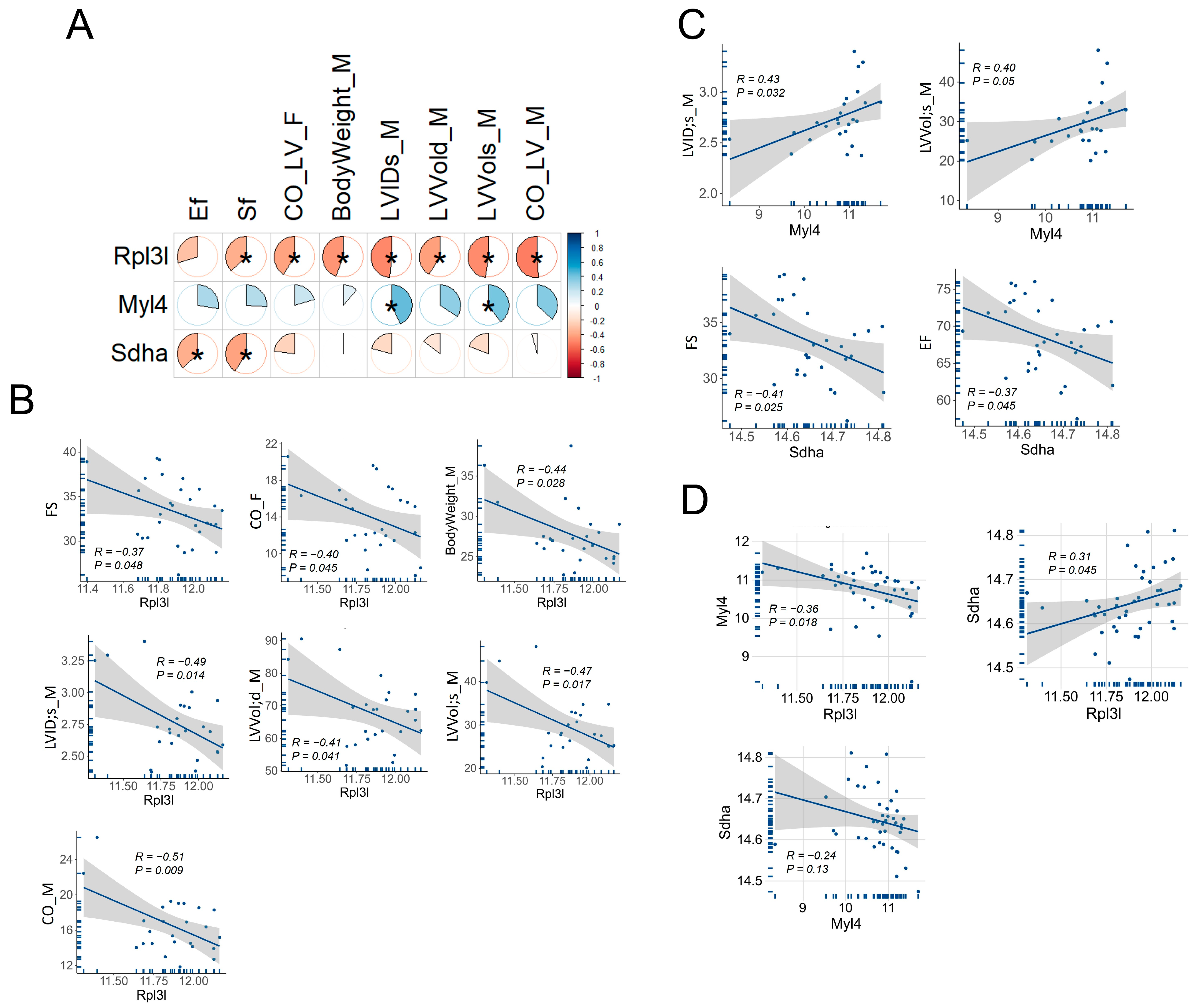
3.6. Correlation of Rpl3l, Myl4 and Sdha with Cardiac Function Traits in BXD Mice
3.7. Analysis of Immune Cell Infiltration between DCM and Normal Heart and Correlations with Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, T.M.; Hsu, D.T.; Kantor, P.; Towbin, J.A.; Ware, S.M.; Colan, S.D.; Chung, W.K.; Jefferies, J.L.; Rossano, J.W.; Castleberry, C.D.; et al. Pediatric Cardiomyopathies. Circ. Res. 2017, 121, 855–873. [Google Scholar] [CrossRef] [PubMed]
- Ganapathi, M.; Argyriou, L.; Martinez-Azorin, F.; Morlot, S.; Yigit, G.; Lee, T.M.; Auber, B.; von Gise, A.; Petrey, D.S.; Thiele, H.; et al. Bi-allelic missense disease-causing variants in RPL3L associate neonatal dilated cardiomyopathy with muscle-specific ribosome biogenesis. Hum. Genet. 2020, 139, 1443–1454. [Google Scholar] [CrossRef] [PubMed]
- Das, B.B.; Gajula, V.; Arya, S.; Taylor, M.B. Compound Heterozygous Missense Variants in RPL3L Genes Associated with Severe Forms of Dilated Cardiomyopathy: A Case Report and Literature Review. Children 2022, 9, 1495. [Google Scholar] [CrossRef] [PubMed]
- Nannapaneni, H.; Ghaleb, S.; Arya, S.; Gajula, V.; Taylor, M.B.; Das, B.B. Further Evidence of Autosomal Recessive Inheritance of RPL3L Pathogenic Variants with Rapidly Progressive Neonatal Dilated Cardiomyopathy. J. Cardiovasc. Dev. Dis. 2022, 9, 65. [Google Scholar] [CrossRef]
- Al-Hassnan, Z.N.; Almesned, A.; Tulbah, S.; Alakhfash, A.; Alhadeq, F.; Alruwaili, N.; Alkorashy, M.; Alhashem, A.; Alrashdan, A.; Faqeih, E.; et al. Categorized Genetic Analysis in Childhood-Onset Cardiomyopathy. Circ. Genom. Precis. Med. 2020, 13, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.P.; Sleeper, L.A.; Lipshultz, S.; Cinar, A.; Canter, C.; Webber, S.A.; Bernstein, D.; Pahl, E.; Alvarez, J.A.; Wilkinson, J.D.; et al. Association of left ventricular dilation at listing for heart transplant with postlisting and early posttransplant mortality in children with dilated cardiomyopathy. Circ. Heart Fail. 2009, 2, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Van Raay, T.J.; Connors, T.D.; Klinger, K.W.; Landes, G.M.; Burn, T.C. A novel ribosomal protein L3-like gene (RPL3L) maps to the autosomal dominant polycystic kidney disease gene region. Genomics 1996, 37, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, T. Ribosome specialization and its potential role in the control of protein translation and skeletal muscle size. J. Appl. Physiol. (1985) 2019, 127, 599–607. [Google Scholar] [CrossRef]
- Xue, S.; Barna, M. Specialized ribosomes: A new frontier in gene regulation and organismal biology. Nat. Rev. Mol. Cell Biol. 2012, 13, 355–369. [Google Scholar] [CrossRef]
- Milenkovic, I.; Santos Vieira, H.G.; Lucas, M.C.; Ruiz-Orera, J.; Patone, G.; Kesteven, S.; Wu, J.; Feneley, M.; Espadas, G.; Sabido, E.; et al. Dynamic interplay between RPL3- and RPL3L-containing ribosomes modulates mitochondrial activity in the mammalian heart. Nucleic Acids Res. 2023, 51, 5301–5324. [Google Scholar] [CrossRef]
- Weng, L.C.; Hall, A.W.; Choi, S.H.; Jurgens, S.J.; Haessler, J.; Bihlmeyer, N.A.; Grarup, N.; Lin, H.; Teumer, A.; Li-Gao, R.; et al. Genetic Determinants of Electrocardiographic P-Wave Duration and Relation to Atrial Fibrillation. Circ. Genom. Precis. Med. 2020, 13, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Thorolfsdottir, R.B.; Sveinbjornsson, G.; Sulem, P.; Nielsen, J.B.; Jonsson, S.; Halldorsson, G.H.; Melsted, P.; Ivarsdottir, E.V.; Davidsson, O.B.; Kristjansson, R.P.; et al. Coding variants in RPL3L and MYZAP increase risk of atrial fibrillation. Commun. Biol. 2018, 1, 68. [Google Scholar] [CrossRef] [PubMed]
- Jaouadi, H.; Chabrak, S.; Lahbib, S.; Abdelhak, S.; Zaffran, S. Identification of two variants in AGRN and RPL3L genes in a patient with catecholaminergic polymorphic ventricular tachycardia suggesting new candidate disease genes and digenic inheritance. Clin. Case Rep. 2022, 10, e05339. [Google Scholar] [CrossRef] [PubMed]
- Kao, B.R.; Malerba, A.; Lu-Nguyen, N.B.; Harish, P.; McCarthy, J.J.; Dickson, G.; Popplewell, L.J. Knockdown of Muscle-Specific Ribosomal Protein L3-Like Enhances Muscle Function in Healthy and Dystrophic Mice. Nucleic Acid Ther. 2021, 31, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, T.; Zhang, X.; McCarthy, J.J. Expression of Muscle-Specific Ribosomal Protein L3-Like Impairs Myotube Growth. J. Cell. Physiol. 2016, 231, 1894–1902. [Google Scholar] [CrossRef] [PubMed]
- Grimes, K.M.; Prasad, V.; Huo, J.; Kuwabara, Y.; Vanhoutte, D.; Baldwin, T.A.; Bowers, S.L.K.; Johansen, A.K.Z.; Sargent, M.A.; Lin, S.J.; et al. Rpl3l gene deletion in mice reduces heart weight over time. Front. Physiol. 2023, 14, 1054169. [Google Scholar] [CrossRef]
- Shiraishi, C.; Matsumoto, A.; Ichihara, K.; Yamamoto, T.; Yokoyama, T.; Mizoo, T.; Hatano, A.; Matsumoto, M.; Tanaka, Y.; Matsuura-Suzuki, E.; et al. RPL3L-containing ribosomes determine translation elongation dynamics required for cardiac function. Nat. Commun. 2023, 14, 2131. [Google Scholar] [CrossRef]
- Orgil, B.O.; Xu, F.; Munkhsaikhan, U.; Alberson, N.R.; Bajpai, A.K.; Johnson, J.N.; Sun, Y.; Towbin, J.A.; Lu, L.; Purevjav, E. Echocardiography phenotyping in murine genetic reference population of BXD strains reveals significant QTLs associated with cardiac function and morphology. Physiol. Genom. 2023, 55, 51–66. [Google Scholar] [CrossRef]
- Gu, Q.; Xu, F.; Orgil, B.O.; Khuchua, Z.; Munkhsaikhan, U.; Johnson, J.N.; Alberson, N.R.; Pierre, J.F.; Black, D.D.; Dong, D.; et al. Systems Genetics Analysis Defines Importance of TMEM43/LUMA for Cardiac and Metabolic Related Pathways. Physiol. Genom. 2021, 54, 22–35. [Google Scholar] [CrossRef]
- Bolstad, B.M.; Irizarry, R.A.; Astrand, M.; Speed, T.P. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef]
- Chesler, E.J.; Lu, L.; Shou, S.; Qu, Y.; Gu, J.; Wang, J.; Hsu, H.C.; Mountz, J.D.; Baldwin, N.E.; Langston, M.A.; et al. Complex trait analysis of gene expression uncovers polygenic and pleiotropic networks that modulate nervous system function. Nat. Genet. 2005, 37, 233–242. [Google Scholar] [CrossRef]
- Consortium, G.T. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Wang, X.; Pandey, A.K.; Mulligan, M.K.; Williams, E.G.; Mozhui, K.; Li, Z.; Jovaisaite, V.; Quarles, L.D.; Xiao, Z.; Huang, J.; et al. Joint mouse-human phenome-wide association to test gene function and disease risk. Nat. Commun. 2016, 7, 10464. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, M.K.; Mozhui, K.; Prins, P.; Williams, R.W. GeneNetwork: A Toolbox for Systems Genetics. Methods Mol. Biol. 2017, 1488, 75–120. [Google Scholar] [CrossRef]
- Smith, C.L.; Blake, J.A.; Kadin, J.A.; Richardson, J.E.; Bult, C.J.; Mouse Genome Database, G. Mouse Genome Database (MGD)-2018: Knowledgebase for the laboratory mouse. Nucleic Acids Res. 2018, 46, D836–D842. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, M.E.; Flenniken, A.M.; Ji, X.; Teboul, L.; Wong, M.D.; White, J.K.; Meehan, T.F.; Weninger, W.J.; Westerberg, H.; Adissu, H.; et al. High-throughput discovery of novel developmental phenotypes. Nature 2016, 537, 508–514. [Google Scholar] [CrossRef]
- Smith, J.R.; Hayman, G.T.; Wang, S.J.; Laulederkind, S.J.F.; Hoffman, M.J.; Kaldunski, M.L.; Tutaj, M.; Thota, J.; Nalabolu, H.S.; Ellanki, S.L.R.; et al. The Year of the Rat: The Rat Genome Database at 20: A multi-species knowledgebase and analysis platform. Nucleic Acids Res. 2020, 48, D731–D742. [Google Scholar] [CrossRef] [PubMed]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef]
- Alliance of Genome Resources, C. Harmonizing model organism data in the Alliance of Genome Resources. Genetics 2022, 220, iyac022. [Google Scholar] [CrossRef]
- Sheng, X.; Wu, J.; Sun, Q.; Li, X.; Xian, F.; Sun, M.; Fang, W.; Chen, M.; Yu, J.; Xiao, J. MTD: A mammalian transcriptomic database to explore gene expression and regulation. Brief. Bioinform. 2017, 18, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Eraslan, G.; Drokhlyansky, E.; Anand, S.; Fiskin, E.; Subramanian, A.; Slyper, M.; Wang, J.; Van Wittenberghe, N.; Rouhana, J.M.; Waldman, J.; et al. Single-nucleus cross-tissue molecular reference maps toward understanding disease gene function. Science 2022, 376, eabl4290. [Google Scholar] [CrossRef] [PubMed]
- Witt, E.; Hammer, E.; Dorr, M.; Weitmann, K.; Beug, D.; Lehnert, K.; Nauck, M.; Volker, U.; Felix, S.B.; Ameling, S. Correlation of gene expression and clinical parameters identifies a set of genes reflecting LV systolic dysfunction and morphological alterations. Physiol. Genom. 2019, 51, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.R.; Zhang, Q.; Lei, Q.; Luo, M.; Xie, G.Y.; Wang, H.; Guo, A.Y. ImmuCellAI: A Unique Method for Comprehensive T-Cell Subsets Abundance Prediction and its Application in Cancer Immunotherapy. Adv. Sci. 2020, 7, 1902880. [Google Scholar] [CrossRef]
- Kontorovich, A.R.; Patel, N.; Moscati, A.; Richter, F.; Peter, I.; Purevjav, E.; Selejan, S.R.; Kindermann, I.; Towbin, J.A.; Bohm, M.; et al. Myopathic Cardiac Genotypes Increase Risk for Myocarditis. JACC Basic Transl. Sci. 2021, 6, 584–592. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, Q.; Qin, Z.; Zhang, S.; Yi, S.; Yi, S.; Zhang, Q.; Luo, J. Novel compound heterozygous variants in the RPL3L gene causing dilated cardiomyopathy type-2D: A case report and literature review. BMC Med. Genom. 2023, 16, 127. [Google Scholar] [CrossRef]
- Zhang, W.; Elimban, V.; Nijjar, M.S.; Gupta, S.K.; Dhalla, N.S. Role of mitogen-activated protein kinase in cardiac hypertrophy and heart failure. Exp. Clin. Cardiol. 2003, 8, 173–183. [Google Scholar]
- Qin, W.; Cao, L.; Massey, I.Y. Role of PI3K/Akt signaling pathway in cardiac fibrosis. Mol. Cell. Biochem. 2021, 476, 4045–4059. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Kitakaze, M. ER stress in cardiovascular disease. J. Mol. Cell. Cardiol. 2010, 48, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Binder, P.; Fang, Q.; Wang, Z.; Xiao, W.; Liu, W.; Wang, X. Endoplasmic reticulum stress in the heart: Insights into mechanisms and drug targets. Br. J. Pharmacol. 2018, 175, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Samarel, A.M. Focal adhesion signaling in heart failure. Pflugers Arch. 2014, 466, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Franchini, K.G.; Clemente, C.F.; Marin, T.M. Focal adhesion kinase signaling in cardiac hypertrophy and failure. Braz. J. Med. Biol. Res. 2009, 42, 44–52. [Google Scholar] [CrossRef]
- de Lucia, C.; Eguchi, A.; Koch, W.J. New Insights in Cardiac β-Adrenergic Signaling During Heart Failure and Aging. Front. Pharmacol. 2018, 9, 904. [Google Scholar] [CrossRef]
- Woodall, B.P.; Gustafsson, A.B. Autophagy-A key pathway for cardiac health and longevity. Acta Physiol. 2018, 223, e13074. [Google Scholar] [CrossRef]
- Hernandez, O.M.; Jones, M.; Guzman, G.; Szczesna-Cordary, D. Myosin essential light chain in health and disease. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1643–H1654. [Google Scholar] [CrossRef]
- Peng, W.; Li, M.; Li, H.; Tang, K.; Zhuang, J.; Zhang, J.; Xiao, J.; Jiang, H.; Li, D.; Yu, Y.; et al. Dysfunction of Myosin Light-Chain 4 (MYL4) Leads to Heritable Atrial Cardiomyopathy with Electrical, Contractile, and Structural Components: Evidence from Genetically-Engineered Rats. J. Am. Heart Assoc. 2017, 6, e007030. [Google Scholar] [CrossRef]
- Zhong, Y.; Tang, K.; Nattel, S.; Zhai, M.; Gong, S.; Yu, Q.; Zeng, Y.; E, G.; Maimaitiaili, N.; Wang, J.; et al. Myosin light-chain 4 gene-transfer attenuates atrial fibrosis while correcting autophagic flux dysregulation. Redox Biol. 2023, 60, 102606. [Google Scholar] [CrossRef]
- Orr, N.; Arnaout, R.; Gula, L.J.; Spears, D.A.; Leong-Sit, P.; Li, Q.; Tarhuni, W.; Reischauer, S.; Chauhan, V.S.; Borkovich, M.; et al. A mutation in the atrial-specific myosin light chain gene (MYL4) causes familial atrial fibrillation. Nat. Commun. 2016, 7, 11303. [Google Scholar] [CrossRef]
- Renkema, G.H.; Wortmann, S.B.; Smeets, R.J.; Venselaar, H.; Antoine, M.; Visser, G.; Ben-Omran, T.; van den Heuvel, L.P.; Timmers, H.J.; Smeitink, J.A.; et al. SDHA mutations causing a multisystem mitochondrial disease: Novel mutations and genetic overlap with hereditary tumors. Eur. J. Hum. Genet. 2015, 23, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Courage, C.; Jackson, C.B.; Hahn, D.; Euro, L.; Nuoffer, J.M.; Gallati, S.; Schaller, A. SDHA mutation with dominant transmission results in complex II deficiency with ocular, cardiac, and neurologic involvement. Am. J. Med. Genet. A 2017, 173, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Levitas, A.; Muhammad, E.; Harel, G.; Saada, A.; Caspi, V.C.; Manor, E.; Beck, J.C.; Sheffield, V.; Parvari, R. Familial neonatal isolated cardiomyopathy caused by a mutation in the flavoprotein subunit of succinate dehydrogenase. Eur. J. Hum. Genet. 2010, 18, 1160–1165. [Google Scholar] [CrossRef] [PubMed]
- Cassandri, M.; Smirnov, A.; Novelli, F.; Pitolli, C.; Agostini, M.; Malewicz, M.; Melino, G.; Raschella, G. Zinc-finger proteins in health and disease. Cell Death Discov. 2017, 3, 17071. [Google Scholar] [CrossRef] [PubMed]
- Jen, J.; Wang, Y.C. Zinc finger proteins in cancer progression. J. Biomed. Sci. 2016, 23, 53. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Wang, Y.; Shaik, G.M.; Burghes, A.H.; Gangwani, L. The zinc finger protein ZPR1 is a potential modifier of spinal muscular atrophy. Hum. Mol. Genet. 2012, 21, 2745–2758. [Google Scholar] [CrossRef]
- Swirski, F.K.; Nahrendorf, M. Cardioimmunology: The immune system in cardiac homeostasis and disease. Nat. Rev. Immunol. 2018, 18, 733–744. [Google Scholar] [CrossRef]
- Gan, T.; Hu, J.; Aledan, A.K.O.; Liu, W.; Li, C.; Lu, S.; Wang, Y.; Xu, Q.; Wang, Y.; Wang, Z. Exploring the pathogenesis and immune infiltration in dilated cardiomyopathy complicated with atrial fibrillation by bioinformatics analysis. Front. Immunol. 2023, 14, 1049351. [Google Scholar] [CrossRef]
- Zhang, X.Z.; Zhang, S.; Tang, T.T.; Cheng, X. Bioinformatics and Immune Infiltration Analyses Reveal the Key Pathway and Immune Cells in the Pathogenesis of Hypertrophic Cardiomyopathy. Front. Cardiovasc. Med. 2021, 8, 696321. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chr (Mb) | Parameters Considered for Scoring | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean Expression | Coding Variant | Cis-Regulation | Sig. Corr. | RGD Causal | Function | Total Score | |||
| Myl4 | 17896 | Chr11:104.550663 | 10.8 | Nonsyn | -- | Y | Y | Y | 7 |
| Ace | 11421 | Chr11:105.967945 | 11.7 | -- | Y | Y | -- | Y | 5 |
| Kpna2 | 16647 | Chr11:106.988629 | 10.3 | Nonsyn | Y | Y | -- | -- | 5 |
| Golm1 | 105348 | Chr13:59.640163 | 9.8 | Nonsyn | Y | Y | -- | -- | 5 |
| Zfp367 | 238673 | Chr13:64.133022 | 8.3 | Nonsyn | Y | Y | -- | -- | 5 |
| Zfp712 | 78251 | Chr13:67.038594 | 8.6 | Nonsyn | Y | Y | -- | -- | 5 |
| Zfp759 | 268670 | Chr13:67.128226 | 8.7 | Nonsyn | Y | Y | -- | -- | 5 |
| Zfp874a | 238692 | Chr13: 67.426259 | 8.2 | Nonsyn | Y | Y | -- | -- | 5 |
| Zfp729a | 212281 | Chr13:67.617001 | 10.5 | Nonsyn | Y | Y | -- | -- | 5 |
| Mtrr | 210009 | Chr13:68.56078 | 9.4 | Nonsyn | Y | Y | -- | -- | 5 |
| Sdha | 66945 | Chr13:74.322254 | 14.6 | -- | -- | Y | Y | Y | 6 |
| Rhobtb3 | 73296 | Chr13:75.869537 | 10.0 | Nonsyn | Y | Y | -- | -- | 5 |
| Immune Cell Types | RPL3L | MYL4 | SDHA | |||
|---|---|---|---|---|---|---|
| R | p | R | p | R | p | |
| Macrophage | 0.30 | 1.06 × 10−1 | −0.01 | 9.55 × 10−1 | 0.13 | 4.90 × 10−1 |
| CD8 T cell | −0.52 | 3.56 × 10−3 | −0.07 | 7.20 × 10−1 | −0.45 | 1.35 × 10−2 |
| Gamma delta T cell | 0.60 | 4.06 × 10−4 | 0.36 | 5.02 × 10−2 | 0.42 | 1.93 × 10−2 |
| CD4-naive T cell | −0.42 | 2.26 × 10−2 | −0.21 | 2.75 × 10−1 | −0.44 | 1.63 × 10−2 |
| CD8-naive T cell | −0.74 | 3.56 × 10−6 | −0.19 | 3.13 × 10−1 | −0.61 | 3.72 × 10−4 |
| Cytotoxic T cell | −0.46 | 9.75 × 10−3 | −0.04 | 8.19 × 10−1 | −0.38 | 4.03 × 10−2 |
| Exhausted T cell | −0.14 | 4.54 × 10−1 | 0.18 | 3.45 × 10−1 | −0.13 | 4.87 × 10−1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bajpai, A.K.; Gu, Q.; Orgil, B.-O.; Alberson, N.R.; Towbin, J.A.; Martinez, H.R.; Lu, L.; Purevjav, E. Exploring the Regulation and Function of Rpl3l in the Development of Early-Onset Dilated Cardiomyopathy and Congestive Heart Failure Using Systems Genetics Approach. Genes 2024, 15, 53. https://doi.org/10.3390/genes15010053
Bajpai AK, Gu Q, Orgil B-O, Alberson NR, Towbin JA, Martinez HR, Lu L, Purevjav E. Exploring the Regulation and Function of Rpl3l in the Development of Early-Onset Dilated Cardiomyopathy and Congestive Heart Failure Using Systems Genetics Approach. Genes. 2024; 15(1):53. https://doi.org/10.3390/genes15010053
Chicago/Turabian StyleBajpai, Akhilesh K., Qingqing Gu, Buyan-Ochir Orgil, Neely R. Alberson, Jeffrey A. Towbin, Hugo R. Martinez, Lu Lu, and Enkhsaikhan Purevjav. 2024. "Exploring the Regulation and Function of Rpl3l in the Development of Early-Onset Dilated Cardiomyopathy and Congestive Heart Failure Using Systems Genetics Approach" Genes 15, no. 1: 53. https://doi.org/10.3390/genes15010053