
Genomics of Treatable Traits in Asthma
 , , , and
, , , and
Abstract
:1. Introduction
2. Asthma Endotypes and Phenotypes
2.1. T2-High Asthma
2.1.1. Early-Onset Atopic Asthma
2.1.2. Late-Onset Eosinophilic Asthma
2.1.3. Nonsteroidal Anti-Inflammatory Drugs-Exacerbated Respiratory Disease (NERD)
2.2. T2-Low Asthma
2.2.1. Obesity-Associated Asthma
2.2.2. Smoking-Associated Asthma Phenotype
2.2.3. Elderly-Associated Asthma Phenotype
2.3. Overlapping in Asthma Phenotypes
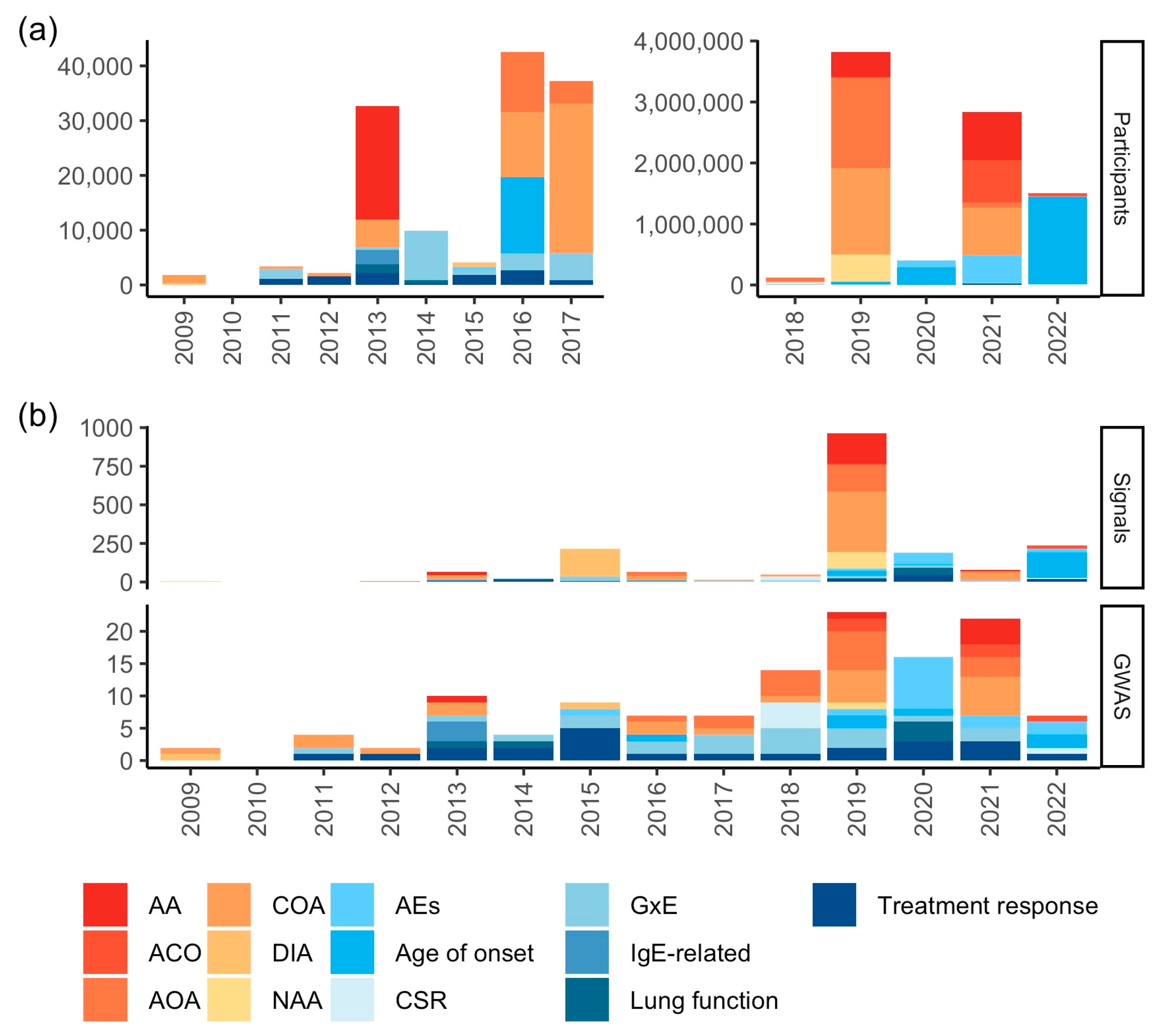
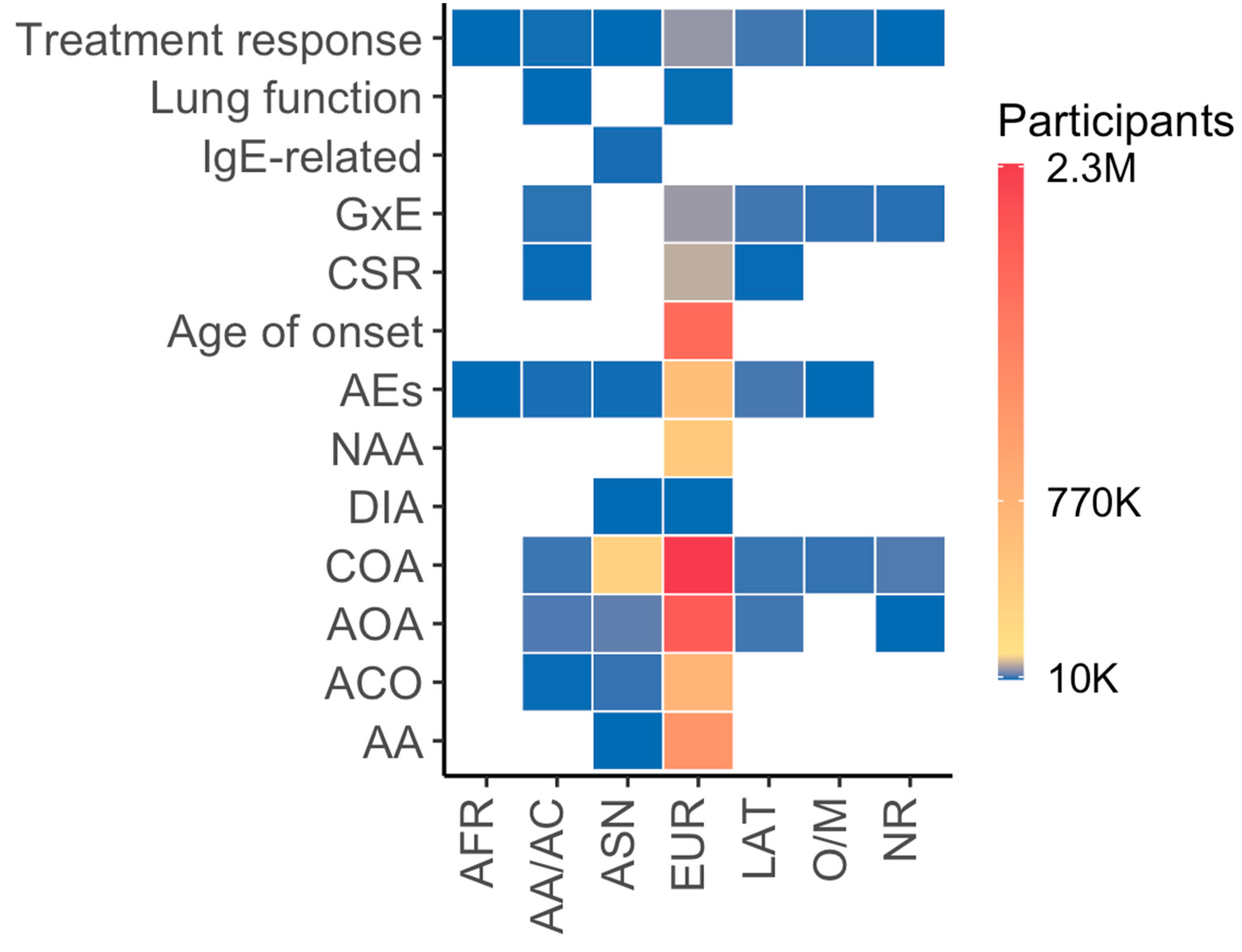
3. Genomic Studies
3.1. Genomic Studies of Asthma Phenotypes
3.2. Genomic Studies of Asthma-Related Traits

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotypes | SNP | Chr. Region a | Genomic Context | Effect Allele | Coefficient Type | Coefficient Value | p-Value | References |
|---|---|---|---|---|---|---|---|---|
| Asthma age onset | rs10208293 | 2q12 | IL1RL1 | G | HR | 1.14 | 3.1 × 10−8 | [89] |
| rs9272346 | 6p21 | HLA-DQA1 | A | HR | 1.13 | 1.6 × 10−8 | ||
| rs928413 | 9p24 | IL33 | G | HR | 1.19 | 6.5 × 10−16 | ||
| rs1861760 | 16q12 | CYLD | A | HR | 1.28 | 4.2 × 10−8 | ||
| rs9901146 | 17q12-q21 | ZPBP2/GSDMB | G | HR | 1.18 | 1.9 × 10−16 | ||
| rs61816761 | 1q21.3 | FLG | A | beta | −4.57 | 8.15 × 10−27 | [90] | |
| rs7518129 | 1q25.1 | TNFSF4 | G | beta | −0.85 | 4.89 × 10−9 | ||
| rs3771175 | 2q12.1 | IL1RL1 | T | beta | −1.73 | 7.66 × 10−17 | ||
| rs10187276 | 2q36.3 | SNRPGP8/CCL20 | T | beta | −0.87 | 1.98 × 10−8 | ||
| rs78147778 | 2q37.3 | D2HGDH | T | beta | −0.91 | 1.64 × 10−8 | ||
| rs2889896 | 3q28 | LPP | C | beta | −0.98 | 8.07 × 10−13 | ||
| rs5743618 | 4p14 | TLR1 | C | beta | −1.58 | 4.53 × 10−22 | ||
| rs4705962 | 5q31.1 | KIF3A | T | beta | −0.99 | 5.57 × 10−10 | ||
| rs12207974 | 6p21.33 | HLA-E/RANP1 | C | beta | −1.07 | 8.86 × 10−11 | ||
| rs1093 | 6p21.33 | HLA-B | G | beta | −0.99 | 8.41 × 10−10 | ||
| rs9274659 | 6p21.32 | HLA-DQB1/MTCO3P1 | A | beta | −1.25 | 6.71 × 10−18 | ||
| rs7848215 | 9p24.1 | IL33 | T | beta | −1.03 | 7.51 × 10−12 | ||
| rs117137535 | 9q34.3 | ARRDC1 | A | beta | −2.46 | 3.42 × 10−8 | ||
| rs61894547 | 11q13.5 | EMSY | T | beta | −2.23 | 4.42 × 10−15 | ||
| rs12365699 | 11q23.3 | CXCR5/DDX6 | G | beta | −1.40 | 4.42 × 10−14 | ||
| rs4795399 | 17q12 | GSDMB | T | beta | −2.29 | 6.76 × 10−65 | ||
| rs11658582 | 17q21.2 | CCR7/SMARCE1 | G | beta | −0.91 | 1.37 × 10−10 | ||
| rs4574025 | 18q21.33 | TNFRSF11A | T | beta | −0.87 | 1.61 × 10−10 | ||
| rs12964116 | 18q21.33 | SERPINB7 | G | beta | −1.92 | 4.87 × 10−8 | ||
| rs5758324 | 22q13.2 | TEF | G | HR | 1.06 | 2.39 × 10−8 | [91] | |
| rs2844649 | 6p21.33 | SFTA2/MUCL3 | A | HR | 1.08 | 4.45 × 10−8 | ||
| Asthma severity | rs560026225 | 4q27 | KIAA1109 | GATT | OR | 1.12 | 3.06 × 10−9 | [93] |
| rs10905284 | 10p14 | GATA3 | A | OR | 0.90 | 1.76 × 10−10 | ||
| rs11603634 | 11p15.5 | MUC5AC | G | OR | 1.09 | 2.32 × 10−8 | ||
| rs7523907 | 1q24.2 | CD247 | T | OR | 1.10 | 4.82 × 10−9 | ||
| rs12479210 | 2q12.1 | IL1RL1 | T | OR | 1.19 | 1.57 × 10−29 | ||
| rs34290285 | 2q37.3 | D2HGDH | A | OR | 0.84 | 2.24 × 10−23 | ||
| rs1837253 | 5q22.1 | TSLP | C | OR | 1.19 | 1.95 × 10−22 | ||
| rs1438673 | 5q22.1 | WDR36 | T | OR | 0.89 | 3.29 × 10−13 | ||
| rs3749833 | 5q31.1 | C5orf56 | C | OR | 1.14 | 5.60 × 10−14 | ||
| rs1986009 | 5q31.1 | RAD50 | A | OR | 1.17 | 2.43 × 10−15 | ||
| rs9273410 | 6p21.32 | HLA-DQB1 | A | OR | 1.21 | 5.62 × 10−32 | ||
| rs144829310 | 9p24.1 | IL33 | T | OR | 1.21 | 2.29 × 10−20 | ||
| rs7936312 | 11q13.5 | C11orf30 | T | OR | 1.17 | 6.18 × 10−24 | ||
| rs703816 | 12q13.3 | STAT6 | C | OR | 1.12 | 3.69 × 10−13 | ||
| rs10519068 | 15q22.2 | RORA | A | OR | 0.85 | 1.84 × 10−12 | ||
| rs72743461 | 15q22.33 | SMAD3 | A | OR | 1.14 | 4.52 × 10−14 | ||
| rs7203459 | 16p13.13 | CLEC16A | C | OR | 0.86 | 4.37 × 10−18 | ||
| rs2941522 | 17q12 | IKZF3 | T | OR | 1.11 | 2.32 × 10−12 | ||
| rs139210940 | 2q12.1 | IL1RL2 | AT | OR | 1.34 | 8.08 × 10−9 | [94] | |
| rs10455025 | 5q22.1 | TSLP | C | OR | 1.30 | 4.36 × 10−13 | ||
| rs17205170 | 6p21.32 | HLA-DQA1 | G | OR | 1.45 | 7.92 × 10−16 | ||
| rs2875584 | 6q15 | BACH2 | C | OR | 1.24 | 1.57 × 10−8 | ||
| rs7130588 | 11q13.5 | C11orf30 | G | OR | 1.24 | 2.46 × 10−9 | ||
| rs2104047 | 14q24.1 | RAD51B | T | OR | 1.25 | 1.28 × 10−8 | ||
| rs11631778 | 15q23 | THSD4 | G | OR | 1.23 | 3.54 × 10−8 | ||
| rs7216558 | 17q12 | GSDMB | T | OR | 1.26 | 1.91 × 10−11 | ||
| Pulmonary function | ||||||||
| FEV1 | rs559389 | 11q13.4 | - | C | beta | −0.03 | 5.28 × 10−5 | [95] |
| rs9366309 | 6p22.3 | - | T | beta | −0.03 | 3.32 × 10−5 | ||
| rs6763931 | 3q23 | ZBTB38 | A | beta | 0.03 | 5.90 × 10−5 | ||
| rs2304725 | 3p25.3 | SLC6A11 | C | beta | 0.03 | 3.87 × 10−5 | ||
| rs17161791 | 7p21.3 | - | C | beta | 0.002 | 3.01 × 10−5 | ||
| rs10795348 | 10p13 | C10orf97 | T | beta | 20.16 | 5.07 × 10−5 | [96] | |
| rs10951730 | 7p13 | HECW1 | A | beta | 20.24 | 9.58 × 10−5 | ||
| rs1291183 | 18p11.32 | YES1 | T | beta | 20.18 | 3.54 × 10−5 | ||
| rs1321267 | 6q23.2 | MOXD1 | A | beta | 20.14 | 5.46 × 10−5 | ||
| rs1843593 | 4q13.2 | GNRHR | T | beta | 20.20 | 5.30 × 10−5 | ||
| rs2040403 | 22q12.3 | SYN3 | A | beta | 0.29 | 1.96 × 10−5 | ||
| rs2063485 | 3q13.11 | ZPLD1 | T | beta | 0.23 | 7.18 × 10−5 | ||
| rs285461 | 1q23.3 | LRRC52 | T | beta | 0.22 | 9.56 × 10−5 | ||
| rs3010301 | 6q24.1 | CITED2 | T | beta | 0.17 | 2.65 × 10−5 | ||
| rs3756089 | 4q35.1 | IRF2 | T | beta | 0.26 | 8.79 × 10−5 | ||
| rs3805383 | 4q12 | NMU | A | beta | 20.17 | 2.35 × 10−5 | ||
| rs388159 | 19p13.11 | IL12RB1 | T | beta | 0.19 | 3.47 × 10−5 | ||
| rs4234121 | 2q37.3 | KIF1A | A | beta | 20.15 | 3.25 × 10−5 | ||
| rs4651208 | 1q25.3 | C1orf21 | T | beta | 20.15 | 6.34 × 10−5 | ||
| rs4735916 | 8p23.3 | ERICH1 | A | beta | 20.24 | 7.02 × 10−5 | ||
| rs5755023 | 22q12.3 | LARGE | A | beta | 20.21 | 1.75 × 10−5 | ||
| rs58667 | 22q13.31 | UPK3A | A | beta | 0.18 | 3.95 × 10−7 | ||
| rs6788848 | 3q13.11 | ZPLD1 | T | beta | 20.18 | 4.99 × 10−5 | ||
| rs7434819 | 4q32.1 | C4orf18 | A | beta | 20.14 | 7.34 × 10−5 | ||
| rs7670758 | 4q31.21 | HHIP | A | beta | 20.14 | 9.50 × 10−5 | ||
| rs7836170 | 8q13.2 | SULF1 | T | beta | 20.16 | 2.15 × 10−5 | ||
| rs925847 | 2q32.2 | STAT4 | T | beta | 0.16 | 8.17 × 10−5 | ||
| rs9364299 | 6q27 | SMOC2 | A | beta | 0.14 | 9.96 × 10−5 | ||
| rs9903394 | 17q24.3 | SOX9 | A | beta | 0.16 | 8.47 × 10−5 | ||
| FVC | rs6482071 | 10p12.31 | BCL11A | T | beta | 20.16 | 6.15 × 10−5 | |
| rs2497714 | 10q25.3 | ALS2CR4 | T | beta | 20.17 | 8.49 × 10−5 | ||
| rs2181563 | 10q25.3 | TNIK | A | beta | 0.20 | 3.50 × 10−5 | ||
| rs10466868 | 12q24.33 | YTHDC1 | T | beta | 0.35 | 1.39 × 10−5 | ||
| rs6500728 | 16p13.3 | KIAA0922 | T | beta | 20.17 | 2.02 × 10−5 | ||
| rs169660 | 16p12.2 | SULF1 | T | beta | 20.17 | 9.77 × 10−5 | ||
| rs1291183 | 18p11.32 | ABLIM1 | A | beta | 0.33 | 4.66 × 10−5 | ||
| rs11085898 | 19p13.12 | PLXDC2 | T | beta | 0.31 | 6.50 × 10−5 | ||
| rs2110565 | 2p16.1 | KIAA1600 | A | beta | 20.17 | 6.72 × 10−5 | ||
| rs1208082 | 2q33.1 | LOC338797 | T | beta | 20.25 | 6.39 × 10−5 | ||
| rs221013 | 20p12.2 | HS3ST2 | T | beta | 20.17 | 4.20 × 10−5 | ||
| rs8115491 | 20q12 | A2BP1 | T | beta | 20.17 | 7.15 × 10−5 | ||
| rs6096573 | 20q13.2 | YES1 | T | beta | 20.18 | 5.47 × 10−5 | ||
| rs9974012 | 20q13.31 | NDUFB7 | A | beta | 0.16 | 4.97 × 10−5 | ||
| rs7281703 | 21q21.1 | PAK7 | T | beta | 0.17 | 3.09 × 10−5 | ||
| rs8140240 | 22q12.3 | ATP9A | T | beta | 0.16 | 5.92 × 10−5 | ||
| rs4133045 | 3q26.2 | PTPRT | T | beta | 0.20 | 7.02 × 10−5 | ||
| rs17592868 | 4q13.2 | BMP7 | T | beta | 0.16 | 8.72 × 10−5 | ||
| rs13119846 | 4q31.3 | C21orf37 | T | beta | 0.33 | 6.98 × 10−5 | ||
| rs7836170 | 8q13.2 | EIF3S7 | A | beta | 20.31 | 9.95 × 10−5 | ||
| FEV1/FVC | rs11032873 | 11p13 | APIP | T | beta | 0.18 | 3.08 × 10−5 | |
| rs11675728 | 2q36.3 | DNER | T | beta | 20.15 | 7.22 × 10−5 | ||
| rs12659620 | 5p15.31 | ADCY2 | T | beta | 0.15 | 5.56 × 10−5 | ||
| rs1406593 | 7p15.2 | SNX10 | T | beta | 20.15 | 7.64 × 10−5 | ||
| rs1416920 | 6p22.1 | ZNF323 | T | beta | 0.16 | 9.46 × 10−5 | ||
| rs17450685 | 10q22.3 | C10orf11 | T | beta | 0.15 | 8.73 × 10−5 | ||
| rs17554448 | 2q31.1 | ZNF650 | A | beta | 0.19 | 8.56 × 10−5 | ||
| rs17646998 | 8q13.2 | SULF1 | T | beta | 20.15 | 3.61 × 10−5 | ||
| rs2063485 | 3q13.11 | ZPLD1 | T | beta | 0.25 | 1.34 × 10−5 | ||
| rs2230739 | 16p13.3 | ADCY9 | A | beta | 0.16 | 6.25 × 10−5 | ||
| rs2705044 | 8p22 | MTMR7 | A | beta | 0.21 | 7.46 × 10−5 | ||
| rs3130696 | 6p21.33 | HLA-C | A | beta | 20.17 | 7.34 × 10−5 | ||
| rs3748540 | 1q43 | GREM2 | A | beta | 0.15 | 7.88 × 10−5 | ||
| rs3809335 | 13q12.13 | MTMR6 | T | beta | 0.28 | 5.40 × 10−5 | ||
| rs4234121 | 2q37.3 | KIF1A | A | beta | 20.15 | 5.32 × 10−5 | ||
| rs5767064 | 22q13.32 | LOC388915 | A | beta | 0.16 | 2.79 × 10−5 | ||
| rs7663065 | 4p15.1 | FLJ45721 | A | beta | 20.15 | 5.43 × 10−5 | ||
| rs8030494 | 15q24.1 | TBC1D21 | A | beta | 0.15 | 7.83 × 10−5 | ||
| rs823673 | 1p34.2 | NFYC | A | beta | 0.19 | 3.94 × 10−5 | ||
| rs9287995 | 2q31.1 | HNRPA3 | T | beta | 0.15 | 6.02 × 10−5 | ||
| rs9362054 | 6q14.3 | C6orf84 | T | beta | 0.14 | 8.68 × 10−5 | ||
| rs9574386 | 13q31.1 | C13orf10 | A | beta | 20.27 | 7.72 × 10−5 | ||
| Asthma remission | [97] | |||||||
| Clinical remission | rs7240102 | 18q21.2 | LOC100130003/C18orf26 | G | OR | 1.99 | 7.9 × 10−5 | |
| Complete remission | rs6581895 | 12q15 | YEATS4/FRS2 | G | OR | 3.83 | 1.3 × 10−5 | |
| rs12405429 | 1q42.2 | FAM89A/TRIM67 | G | OR | 3.10 | 1.0 × 10−4 | ||
| rs1420101 | 2q12.1 | IL18R1/IL1RL1 | A | OR | 0.44 | 3.4 × 10−4 | ||
| T2-low asthma | rs117639512 | 19q13.41 | KLK5 | A | OR | 0.33 | 2.75 × 10−8 | [101] |
| Total IgE | rs10404342 | 19q13.43 | ZNF71 | C | NA | NA | 7.60 × 10−6 | [102] |
| rs4879926 | 9p13.3 | TLN1 | C | NA | NA | 7.74 × 10−60 | ||
| Eosinophil-specific proteins | [104] | |||||||
| EDN | rs72677651 | 1p31.3 | JAK1/AK4 | T | beta | 0.54 | 2.0 × 10−8 | |
| rs76335186 | 7p21.3 | NDUFA4 | G | beta | −0.55 | 4.9 × 10−8 | ||
| rs67049014 | 14q11.2 | RNASE2/METTL17 | A | beta | −0.32 | 3 × 10−12 | ||
| ECP | rs56675562 | 9q22.1 | CDK20/SPATA31C2 | G | beta | −0.56 | 5.1 × 10−9 | |
| ECP-EDN | rs116571378 | 2p13.3 | ARHGAP25 | T | beta | NA | 4.2 × 10−10 | |
| rs67049014 | 14q11.2 | RNASE2/METTL17 | A | beta | NA | 1 × 10−13 | ||
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix A.1. Literature and Database Mining
| Asthma-Related Traits | Asthma Phenotype |
|---|---|
| AES | AA |
| Asthma exacerbations | Atopic asthma |
| Asthma with exacerbation (PheCode 495.2) | Asthma and hay fever |
| Childhood asthma exacerbations in long-acting beta2-agonist treatment | Asthma and eczema |
| Childhood asthma with severe exacerbations | ICD10 J45.0: Predominantly allergic asthma |
| Severe exacerbations in childhood asthma | ICD10 J45.0: Predominantly allergic asthma (Gene-based burden) |
| Exacerbations requiring hospitalization in asthma | ACO |
| Asthma exacerbations in inhaled corticosteroid treatment | Asthma–COPD overlap syndrome |
| Asthma with severe exacerbations | Asthma–chronic obstructive pulmonary disease overlap syndrome in asthma |
| Age of onset | Asthma–COPD overlap syndrome (Gene-based burden) |
| Asthma (time to childhood onset) in early life tobacco smoke exposure) | Asthma–chronic obstructive pulmonary disease overlap syndrome |
| Asthma (time to onset) | AOA |
| Asthma (age of onset) | Adult-onset asthma in non-smokers |
| Age of onset of childhood-onset asthma | Adult-onset asthma in ever-smokers |
| Age of onset of adult-onset asthma | Adult asthma (Gene-based burden) |
| Asthma (time to event) | Asthma (adult onset) |
| CSR | Adult asthma |
| Asthma (moderate or severe) | COA |
| Clinical remission in asthma | Pediatric asthma |
| Complete remission in asthma | Self-reported childhood asthma in adult smokers |
| Asthma control | Asthma (childhood onset) |
| GxE | Childhood asthma |
| Asthma (sex interaction) | Asthma onset (childhood vs. adult) |
| Adult onset asthma (smoking interaction) | DIA |
| Childhood asthma x sex interaction | Asthma (toluene diisocyanate-induced) |
| Asthma (SNP x SNP interaction) | Diisocyanate-induced asthma |
| Asthma control x inhaled corticosteroid treatment interaction (1df) | NAA |
| Asthma control x inhaled corticosteroid treatment interaction (2df) | Nonatopic asthma |
| Asthma x Hispanic interaction (2df) | |
| Bronchodilator response x age interaction in asthma | |
| Bone mineral accretion in asthma (oral corticosteroid dose interaction) | |
| Response to zileuton treatment in asthma (FEV1 change interaction) | |
| Asthma (time to childhood onset) x early life tobacco smoke interaction | |
| Adult onset asthma (smoking interaction) | |
| Asthma (sex interaction) | |
| Asthma (SNP x SNP interaction) | |
| Asthma or atopy (farm exposure interaction) | |
| Asthma x air pollution interaction (2df) | |
| Asthma x Hispanic interaction (2df) | |
| Childhood asthma x sex interaction | |
| Childhood onset asthma (traffic air pollution exposure interaction) | |
| Lung function (FEV1) in asthma (dust mite allergen exposure interaction) | |
| Lung function (FEV1/FVC) in asthma (dust mite allergen exposure interaction) | |
| Adverse response to inhaled corticosteroid treatment x age interaction in asthma | |
| Bronchodilator response in asthma (inhaled corticosteroid treatment interaction) | |
| Post-bronchodilator FEV1 x air pollution (CO) interaction in childhood asthma) | |
| Asthma x air pollution interaction (2df) | |
| Post-bronchodilator FEV1 x air pollution (NO2) interaction in childhood asthma, | |
| Childhood onset asthma (traffic air pollution exposure interaction) | |
| IgE-related | |
| IgE levels in asthmatics | |
| IgE levels in asthmatics (D.f. specific) | |
| IgE levels in asthmatics (D.p. specific) | |
| Lung function | |
| Pulmonary function in asthmatics, | |
| Lung function (FEV1) in asthma | |
| Lung function (FVC) in asthma | |
| Lung function (FEV1/FVC) in asthma | |
| Treatment response | |
| Post-bronchodilator lung function in asthma (FEV1) | |
| Post-bronchodilator lung function in asthma (FVC) | |
| Post-bronchodilator lung function in asthma (FEV1/FVC) | |
| Asthma treatment response | |
| Asthma (bronchodilator response) | |
| Asthma (corticosteroid response) | |
| Response to inhaled corticosteroid treatment in asthma (change in FEV1) | |
| Response to inhaled glucocorticoid treatment in asthma (change in FEV1) | |
| Bronchodilator response in asthma | |
| Response to montelukast in asthma (change in FEV1) | |
| Response to mepolizumab in severe asthma | |
| Response to placebo treatment in childhood asthma (FVC change) | |
| Oral corticosteroid burst in asthma | |
| Recent medication for asthma (UKB data field 22167) | |
| Recent medication for asthma (UKB data field 22167) (Gene-based burden) | |
| Subjective response to placebo treatment in childhood asthma (change in cough/wheeze) |
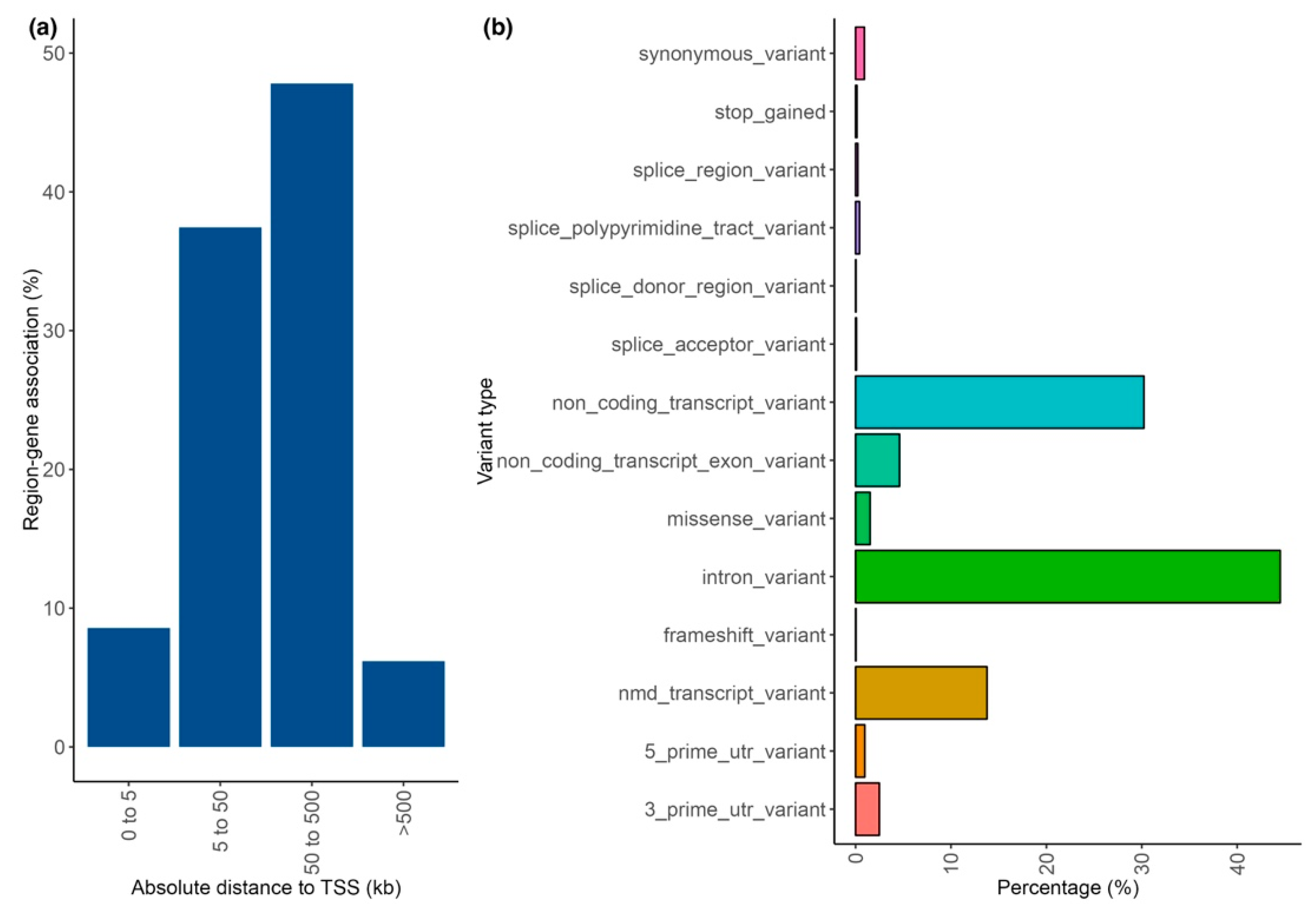
Appendix A.2. Variant Annotation
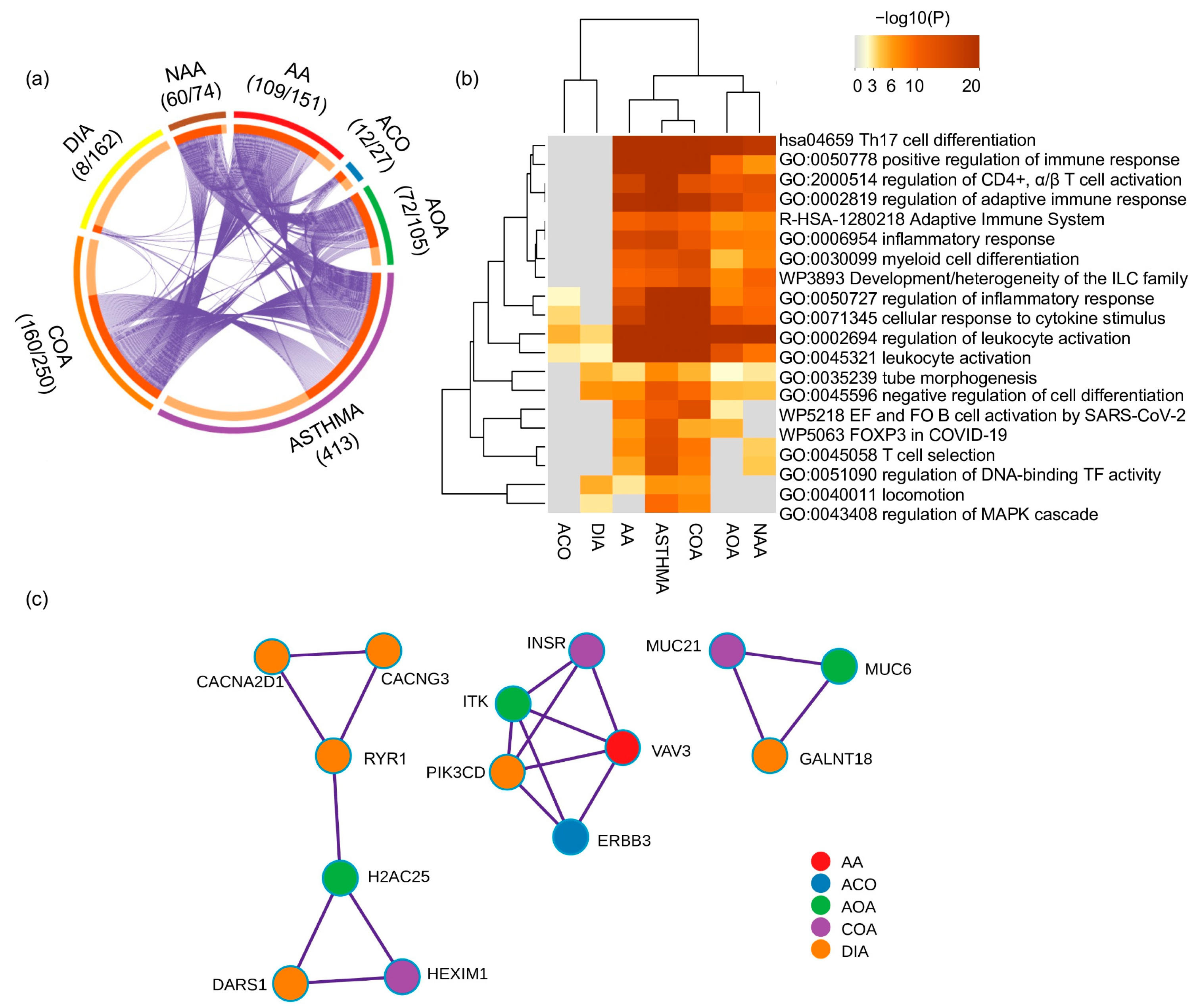
Appendix A.3. Enrichment and Protein–Protein Interaction Network Analyses
References
- Ferkol, T.; Schraufnagel, D. The Global Burden of Respiratory Disease. Ann. Am. Thorac. Soc. 2014, 11, 404–406. [Google Scholar] [CrossRef] [PubMed]
- Global Initiative for Asthma. Global Strategy for Asthma Management and Prevention. 2023. Available online: https://ginasthma.org/ (accessed on 27 August 2023).
- Price, D.; Fletcher, M.; Molen, T. van der Asthma Control and Management in 8000 European Patients: The REcognise Asthma and Link to Symptoms and Experience (REALISE) Survey. NPJ Prim. Care Respir. Med. 2014, 24, 14009. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.L. The National Review of Asthma Deaths: What Did We Learn and What Needs to Change? Breathe Sheff 2015, 11, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Sadatsafavi, M.; Rousseau, R.; Chen, W.; Zhang, W.; Lynd, L.; FitzGerald, J.M. The Preventable Burden of Productivity Loss Due to Suboptimal Asthma Control: A Population-Based Study. Chest 2014, 145, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Vicente, C.T.; Revez, J.A.; Ferreira, M.A.R. Lessons from Ten Years of Genome-Wide Association Studies of Asthma. Clin. Transl. Immunol. 2017, 6, e165. [Google Scholar] [CrossRef] [PubMed]
- Sakula, A. Henry Hyde Salter (1823-71): A Biographical Sketch. Thorax 1985, 40, 887–888. [Google Scholar] [CrossRef]
- Wenzel, S.E. Asthma Phenotypes: The Evolution from Clinical to Molecular Approaches. Nat. Med. 2012, 18, 716–725. [Google Scholar] [CrossRef]
- Rackemann, F.M. A Working Classification of Asthma. Am. J. Med. 1947, 3, 601–606. [Google Scholar] [CrossRef]
- McFadden, E.R. A Century of Asthma. Am. J. Respir. Crit. Care Med. 2004, 170, 215–221. [Google Scholar] [CrossRef]
- Hinks, T.S.C.; Levine, S.J.; Brusselle, G.G. Treatment Options in Type-2 Low Asthma. Eur. Respir. J. 2021, 57, 2000528. [Google Scholar] [CrossRef]
- Woodruff, P.G.; Modrek, B.; Choy, D.F.; Jia, G.; Abbas, A.R.; Ellwanger, A.; Koth, L.L.; Arron, J.R.; Fahy, J.V. T-Helper Type 2-Driven Inflammation Defines Major Subphenotypes of Asthma. Am. J. Respir. Crit. Care Med. 2009, 180, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Busse, W.W.; Holgate, S.T.; Wenzel, S.W.; Klekotka, P.; Chon, Y.; Feng, J.; Ingenito, E.P.; Nirula, A. Biomarker Profiles in Asthma with High vs Low Airway Reversibility and Poor Disease Control. Chest 2015, 148, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The Cytokines of Asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef] [PubMed]
- Svenningsen, S.; Nair, P. Asthma Endotypes and an Overview of Targeted Therapy for Asthma. Front. Med. Lausanne 2017, 4, 158. [Google Scholar] [CrossRef] [PubMed]
- Moore, W.C.; Meyers, D.A.; Wenzel, S.E.; Teague, W.G.; Li, H.; Li, X.; Jr, R.D.; Castro, M.; Curran-Everett, D.; Fitzpatrick, A.M.; et al. Identification of Asthma Phenotypes Using Cluster Analysis in the Severe Asthma Research Program. Am. J. Respir. Crit. Care Med. 2010, 181, 315–323. [Google Scholar] [CrossRef]
- Shaw, D.E.; Sousa, A.R.; Fowler, S.J.; Fleming, L.J.; Roberts, G.; Corfield, J.; Pandis, I.; Bansal, A.T.; Bel, E.H.; Auffray, C.; et al. Clinical and Inflammatory Characteristics of the European U-BIOPRED Adult Severe Asthma Cohort. Eur. Respir. J. 2015, 46, 1308–1321. [Google Scholar] [CrossRef]
- Loza, M.J.; Djukanovic, R.; Chung, K.F.; Horowitz, D.; Ma, K.; Branigan, P.; Barnathan, E.S.; Susulic, V.S.; Silkoff, P.E.; Sterk, P.J.; et al. Validated and Longitudinally Stable Asthma Phenotypes Based on Cluster Analysis of the ADEPT Study. Respir. Res. 2016, 17, 165. [Google Scholar] [CrossRef]
- Haldar, P.; Pavord, I.D.; Shaw, D.E.; Berry, M.A.; Thomas, M.; Brightling, C.E.; Wardlaw, A.J.; Green, R.H. Cluster Analysis and Clinical Asthma Phenotypes. Am. J. Respir. Crit. Care Med. 2008, 178, 218–224. [Google Scholar] [CrossRef]
- Kuruvilla, M.E.; Lee, F.E.-H.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef]
- Anderson, G.P. Endotyping Asthma: New Insights into Key Pathogenic Mechanisms in a Complex, Heterogeneous Disease. Lancet 2008, 372, 1107–1119. [Google Scholar] [CrossRef]
- Ray, A.; Camiolo, M.; Fitzpatrick, A.; Gauthier, M.; Wenzel, S.E. Are We Meeting the Promise of Endotypes and Precision Medicine in Asthma? Physiol. Rev. 2020, 100, 983–1017. [Google Scholar] [CrossRef]
- Locksley, R.M. Asthma and Allergic Inflammation. Cell 2010, 140, 777–783. [Google Scholar] [CrossRef]
- Chung, K.F. Asthma Phenotyping: A Necessity for Improved Therapeutic Precision and New Targeted Therapies. J. Intern. Med. 2016, 279, 192–204. [Google Scholar] [CrossRef]
- Akar-Ghibril, N.; Casale, T.; Custovic, A.; Phipatanakul, W. Allergic Endotypes and Phenotypes of Asthma. J. Allergy Clin. Immunol. Prac. 2020, 8, 429–440. [Google Scholar] [CrossRef]
- Jartti, T.; Gern, J.E. Role of Viral Infections in the Development and Exacerbation of Asthma in Children. J. Allergy Clin. Immunol. 2017, 140, 895–906. [Google Scholar] [CrossRef]
- Foronjy, R.; D’Armiento, J. The Effect of Cigarette Smoke-Derived Oxidants on the Inflammatory Response of the Lung. Clin. Appl. Immunol. Rev. 2006, 6, 53–72. [Google Scholar] [CrossRef]
- Martinez, F.D. Childhood Asthma Inception and Progression: Role of Microbial Exposures, Susceptibility to Viruses and Early Allergic Sensitization. Immunol. Allergy Clin. N. Am. 2019, 39, 141–150. [Google Scholar] [CrossRef]
- Ayers, A.B.; Henry, K.; Russell, S.B.; Steiner, R.E. The Microvasculature of the Spleen. Clin. Radiol. 1976, 27, 259–264. [Google Scholar] [CrossRef]
- Fahy, J.V. Type 2 Inflammation in Asthma--Present in Most, Absent in Many. Nat. Rev. Immunol. 2015, 15, 57–65. [Google Scholar] [CrossRef]
- Peters, M.C.; Kerr, S.; Dunican, E.M.; Woodruff, P.G.; Fajt, M.L.; Levy, B.D.; Israel, E.; Phillips, B.R.; Mauger, D.T.; Comhair, S.A.; et al. Refractory Airway Type 2 Inflammation in a Large Subgroup of Asthmatic Patients Treated with Inhaled Corticosteroids. J. Allergy Clin. Immunol. 2019, 143, 104–113.e14. [Google Scholar] [CrossRef]
- Amelink, M.; de Groot, J.C.; de Nijs, S.B.; Lutter, R.; Zwinderman, A.H.; Sterk, P.J.; Brinke, A.T.; Bel, E.H. Severe Adult-Onset Asthma: A Distinct Phenotype. J. Allergy Clin. Immunol. 2013, 132, 336–341. [Google Scholar] [CrossRef]
- White, A.A.; Doherty, T.A. Role of Group 2 Innate Lymphocytes in Aspirin-Exacerbated Respiratory Disease Pathogenesis. Am. J. Rhinol. Allergy 2018, 32, 7–11. [Google Scholar] [CrossRef]
- Laidlaw, T.M.; Mullol, J.; Woessner, K.M.; Amin, N.; Mannent, L.P. Chronic Rhinosinusitis with Nasal Polyps and Asthma. J. Allergy Clin. Immunol. Prac. 2021, 9, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Swierczynska, M.; Nizankowska-Mogilnicka, E.; Zarychta, J.; Gielicz, A.; Szczeklik, A. Nasal versus bronchial and nasal response to oral aspirin challenge: Clinical and biochemical differences between patients with aspirin-induced asthma/rhinitis. J. Allergy Clin. Immunol. 2003, 112, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Daffern, P.J.; Muilenburg, D.; Hugli, T.E.; Stevenson, D.D. Association of urinary leukotriene E4 excretion during aspirin challenges with severity of respiratory responses. J. Allergy Clin. Immunol. 1999, 104, 559–564. [Google Scholar] [CrossRef]
- Micheletto, C.; Tognella, S.; Visconti, M.; Trevisan, F.; Dal Negro, R.W. Changes in urinary LTE4 and nasal functions following nasal provocation test with ASA in ASA-tolerant and -intolerant asthmatics. Respir. Med. 2006, 100, 2144–2150. [Google Scholar] [CrossRef]
- Laidlaw, T.M.; Boyce, J.A. Pathogenesis of aspirin-exacerbated respiratory disease and reactions. Immunol. Allergy Clin. N. Am. 2013, 33, 195–210. [Google Scholar] [CrossRef]
- Cavagnero, K.J.; Doherty, T.A. Lipid-Mediated Innate Lymphoid Cell Recruitment and Activation in Aspirin-Exacerbated Respiratory Disease. Ann. Allergy Asthma Immunol. 2021, 126, 135–142. [Google Scholar] [CrossRef]
- Eastman, J.J.; Cavagnero, K.J.; Deconde, A.S.; Kim, A.S.; Karta, M.R.; Broide, D.H.; Zuraw, B.L.; White, A.A.; Christiansen, S.C.; Doherty, T.A. Group 2 Innate Lymphoid Cells Are Recruited to the Nasal Mucosa in Patients with Aspirin-Exacerbated Respiratory Disease. J. Allergy Clin. Immunol. 2017, 140, 101–108.e3. [Google Scholar] [CrossRef]
- Fitzpatrick, A.M.; Chipps, B.E.; Fernando, P.G.H. Woodruff T2-“Low” Asthma: Overview and Management Strategies. J. Allergy Clin. Immunol. Prac. 2020, 8, 452–463. [Google Scholar] [CrossRef]
- Hudey, S.N.; Ledford, D.K.; Cardet, J.C. Mechanisms of Non-Type 2 Asthma. Curr. Opin. Immunol. 2020, 66, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.L.; Scott, R.; Boyle, M.J.; Gibson, P.G. Inflammatory Subtypes in Asthma: Assessment and Identification Using Induced Sputum. Respirology 2006, 11, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Carr, T.F. Treatment Approaches for the Patient with T2 Low Asthma. Ann. Allergy Asthma Immunol. 2021, 127, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Hastie, A.T.; Moore, W.C.; Meyers, D.A.; Vestal, P.L.; Li, H.; Peters, S.P.; Bleecker, E.R.; Lung, N.H.; Program, B.I.S.A.R. Analyses of Asthma Severity Phenotypes and Inflammatory Proteins in Subjects Stratified by Sputum Granulocytes. J. Allergy Clin. Immunol. 2010, 125, 1028–1036.e13. [Google Scholar] [CrossRef] [PubMed]
- Rossios, C.; Pavlidis, S.; Hoda, U.; Kuo, C.-H.; Wiegman, C.; Russell, K.; Sun, K.; Loza, M.J.; Baribaud, F.; Durham, A.L.; et al. Sputum Transcriptomics Reveal Upregulation of IL-1 Receptor Family Members in Patients with Severe Asthma. J. Allergy Clin. Immunol. 2018, 141, 560–570. [Google Scholar] [CrossRef]
- Peters, M.C.; McGrath, K.W.; Hawkins, G.A.; Hastie, A.T.; Levy, B.D.; Israel, E.; Phillips, B.R.; Mauger, D.T.; Comhair, S.A.; Erzurum, S.C.; et al. Plasma Interleukin-6 Concentrations, Metabolic Dysfunction, and Asthma Severity: A Cross-Sectional Analysis of Two Cohorts. Lancet Respir. Med. 2016, 4, 574–584. [Google Scholar] [CrossRef]
- Ntontsi, P.; Loukides, S.; Bakakos, P.; Kostikas, K.; Papatheodorou, G.; Papathanassiou, E.; Hillas, G.; Koulouris, N.; Papiris, S.; Papaioannou, A.I. Clinical, Functional and Inflammatory Characteristics in Patients with Paucigranulocytic Stable Asthma: Comparison with Different Sputum Phenotypes. Allergy 2017, 72, 1761–1767. [Google Scholar] [CrossRef]
- Tliba, O.; Jr, R.A.P. Paucigranulocytic Asthma: Uncoupling of Airway Obstruction from Inflammation. J. Allergy Clin. Immunol. 2019, 143, 1287–1294. [Google Scholar] [CrossRef]
- Ogawa, H.; Azuma, M.; Umeno, A.; Shimizu, M.; Murotomi, K.; Yoshida, Y.; Nishioka, Y.; Tsuneyama, K. Singlet Oxygen -Derived Nerve Growth Factor Exacerbates Airway Hyperresponsiveness in a Mouse Model of Asthma with Mixed Inflammation. Allergol. Int. 2022, 71, 395–404. [Google Scholar] [CrossRef]
- Braun, A.; Quarcoo, D.; Schulte-Herbrüggen, O.; Lommatzsch, M.; Hoyle, G.; Renz, H. Nerve Growth Factor Induces Airway Hyperresponsiveness in Mice. Int. Arch. Allergy Immunol. 2001, 124, 205–207. [Google Scholar] [CrossRef]
- Kyriakopoulos, C.; Gogali, A.; Bartziokas, K.; Kostikas, K. Identification and Treatment of T2-Low Asthma in the Era of Biologics. ERJ Open Res. 2021, 7, 309–2020. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.; O’Byrne, P.M. Measuring Eosinophils to Make Treatment Decisions in Asthma. Chest 2016, 150, 485–487. [Google Scholar] [CrossRef] [PubMed]
- Thomson, N.C. Novel Approaches to the Management of Noneosinophilic Asthma. Ther. Adv. Respir. Dis. 2016, 10, 211–234. [Google Scholar] [CrossRef]
- Mohan, A.; Grace, J.; Wang, B.R.; Lugogo, N. The Effects of Obesity in Asthma. Curr. Allergy Asthma Rep. 2019, 19, 49. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.E.; Que, L.G. Obesity and Asthma. Semin. Respir. Crit. Care Med. 2022, 43, 662–674. [Google Scholar] [CrossRef]
- Ignacio, R.M.C.; Kim, C.-S.; Kim, S.-K. Immunological Profiling of Obesity. J. Lifestyle Med. 2014, 4, 1–7. [Google Scholar] [CrossRef]
- Himmerich, H.; Fulda, S.; Linseisen, J.; Seiler, H.; Wolfram, G.; Stephanie, K.H.; Gedrich; Pollmächer, T. TNF-α, Soluble TNF Receptor and Interleukin-6 Plasma Levels in the General Population. Eur. Cytokine Netw. 2006, 17, 196–201. [Google Scholar] [PubMed]
- Björkander, S.; Klevebro, S.; Hernandez-Pacheco, N.; Kere, M.; Ekström, S.; Mikus, M.S.; van Hage, M.; James, A.; Kull, I.; Bergström, A.; et al. Obese Asthma Phenotypes Display Distinct Plasma Biomarker Profiles. Clin. Transl. Allergy 2023, 13, e12238. [Google Scholar] [CrossRef]
- Li, X.; Hastie, A.T.; Peters, M.C.; Hawkins, G.A.; Phipatanakul, W.; Li, H.; Moore, W.C.; Busse, W.W.; Castro, M.; Erzurum, S.C.; et al. Investigation of the Relationship between IL-6 and Type 2 Biomarkers in Patients with Severe Asthma. J. Allergy Clin. Immunol. 2020, 145, 430–433. [Google Scholar] [CrossRef]
- Holguin, F.; Comhair, S.A.A.; Hazen, S.L.; Powers, R.W.; Khatri, S.S.; Bleecker, E.R.; Busse, W.W.; Calhoun, W.J.; Castro, M.; Fitzpatrick, A.M.; et al. An Association between L-Arginine/Asymmetric Dimethyl Arginine Balance, Obesity, and the Age of Asthma Onset Phenotype. Am. J. Respir. Crit. Care Med. 2013, 187, 153–159. [Google Scholar] [CrossRef]
- Barcala, F.J.G.; de la Fuente-Cid, R.; Alvarez-Gil, R.; Tafalla, M.; Nuevo, J.; Caamaño-Isorna, F. Factores Asociados Con El Control Del Asma En Pacientes de Atención Primaria En España: El Estudio CHAS. Arch. Bronconeumol. 2010, 46, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Gorgojo, I.; Antolín-Amérigo, D.; Domínguez-Ortega, J.; Quirce, S. Non-Eosinophilic Asthma: Current Perspectives. J. Asthma Allergy 2018, 11, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Boulet, L.-P.; Lemière, C.; Archambault, F.; Carrier, G.; Claire, F.D.M. Deschesnes Smoking and Asthma: Clinical and Radiologic Features, Lung Function, and Airway Inflammation. Chest 2006, 129, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Kiljander, T.; Poussa, T.; Helin, T.; Jaakkola, A.; Venho, K.; Lehtimäki, L. Symptom Control among Asthmatics with a Clinically Significant Smoking History: A Cross-Sectional Study in Finland. BMC Pulm. Med. 2020, 20, 88. [Google Scholar] [CrossRef]
- Tiotiu, A.; Ioan, I.; Wirth, N.; Romero-Fernandez, R.; González-Barcala, F.-J. The Impact of Tobacco Smoking on Adult Asthma Outcomes. Int. J. Environ. Res. Public Health 2021, 18, 992. [Google Scholar] [CrossRef]
- Polosa, R.; Thomson, N.C. Smoking and Asthma: Dangerous Liaisons. Eur. Respir. J. 2013, 41, 716–726. [Google Scholar] [CrossRef]
- Cazzola, M.; Polosa, R. Anti-TNF-α and Th1 Cytokine-Directed Therapies for the Treatment of Asthma. Curr. Opin. Allergy Clin. Immunol. 2006, 6, 43–50. [Google Scholar] [CrossRef]
- Bujarski, S.; Parulekar, A.D.; Amir, N.A.S. Hanania the Asthma COPD Overlap Syndrome (ACOS). Curr. Allergy Asthma Rep. 2015, 15, 509. [Google Scholar] [CrossRef]
- Gibson, P.G.; McDonald, V.M.; Marks, G.B. Asthma in Older Adults. Lancet 2010, 376, 803–813. [Google Scholar] [CrossRef]
- Ducharme, M.-E.; Prince, P.; Hassan, N.; Nair, P.; Boulet, L.-P. Expiratory Flows and Airway Inflammation in Elderly Asthmatic Patients. Respir. Med. 2011, 105, 1284–1289. [Google Scholar] [CrossRef]
- Dunn, R.M.; Busse, P.J.; Wechsler, M.E. Asthma in the Elderly and Late-Onset Adult Asthma. Allergy 2018, 73, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, V.; Rink, L.; Uciechowski, P. The Th17/Treg Balance Is Disturbed during Aging. Exp. Gerontol. 2013, 48, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Guzman, E.; Mannino, D.M. Airway Obstructive Diseases in Older Adults: From Detection to Treatment. J. Allergy Clin. Immunol 2010, 126, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.Y.; Zhang, X.; Wang, J.; Wang, G.; Oliver, B.G.; Zhang, H.P.; Kang, D.Y.; Wang, L.; Qiu, Z.X.; Li, W.M.; et al. Multidimensional Assessment of Asthma Identifies Clinically Relevant Phenotype Overlap: A Cross-Sectional Study. J. Allergy Clin. Immunol. Prac. 2021, 9, 349–362.e18. [Google Scholar] [CrossRef] [PubMed]
- Quirce, S.; Sastre, J. Occupational asthma: Clinical phenotypes, biomarkers, and management. Curr. Opin. Pulm. Med. 2019, 25, 59–63. [Google Scholar] [CrossRef]
- Vandenplas, O.; Godet, J.; Hurdubaea, L.; Rifflart, C.; Suojalehto, H.; Wiszniewska, M.; Munoz, X.; Sastre, J.; Klusackova, P.; Moore, V.; et al. Are high- and low-molecular-weight sensitizing agents associated with different clinical phenotypes of occupational asthma? Allergy 2019, 74, 261–272. [Google Scholar] [CrossRef]
- Ricciardolo, F.L.M.; Guida, G.; Bertolini, F.; Stefano, A.D.; Carriero, V. Phenotype Overlap in the Natural History of Asthma. Eur. Respir. Rev. 2023, 32, 220201. [Google Scholar] [CrossRef]
- Frey, U.; Suki, B. Complexity of Chronic Asthma and Chronic Obstructive Pulmonary Disease: Implications for Risk Assessment, and Disease Progression and Control. Lancet 2008, 372, 1088–1099. [Google Scholar] [CrossRef]
- González-Pérez, R.; Poza-Guedes, P.; Mederos-Luis, E.; Sánchez-Machín, I. Real-Life Performance of Mepolizumab in T2-High Severe Refractory Asthma with the Overlapping Eosinophilic-Allergic Phenotype. Biomedicines 2022, 10, 2635. [Google Scholar] [CrossRef]
- Kim, K.W.; Ober, C. Lessons Learned from GWAS of Asthma. Allergy Asthma Immunol. Res. 2019, 11, 170–187. [Google Scholar] [CrossRef]
- Sollis, E.; Mosaku, A.; Abid, A.; Buniello, A.; Cerezo, M.; Gil, L.; Groza, T.; Güneş, O.; Hall, P.; Hayhurst, J.; et al. The NHGRI-EBI GWAS Catalog: Knowledgebase and Deposition Resource. Nucleic Acids Res. 2023, 51, D977–D985. [Google Scholar] [CrossRef] [PubMed]
- Sakaue, S.; Kanai, M.; Tanigawa, Y.; Karjalainen, J.; Kurki, M.; Koshiba, S.; Narita, A.; Konuma, T.; Yamamoto, K.; Akiyama, M.; et al. A Cross-Population Atlas of Genetic Associations for 220 Human Phenotypes. Nat. Genet. 2021, 53, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Tsuo, K.; Zhou, W.; Wang, Y.; Kanai, M.; Namba, S.; Gupta, R.; Majara, L.; Nkambule, L.L.; Morisaki, T.; Okada, Y.; et al. Multi-Ancestry Meta-Analysis of Asthma Identifies Novel Associations and Highlights the Value of Increased Power and Diversity. Cell Genom. 2022, 2, 100212. [Google Scholar] [CrossRef]
- Herrera-Luis, E.; Forno, E.; Celedón, J.C.; Pino-Yanes, M. Asthma Exacerbations: The Genes Behind the Scenes. J. Investig. Allergol. Clin. Immunol. 2023, 33, 76–94. [Google Scholar] [CrossRef]
- Hernandez-Pacheco, N.; Pino-Yanes, M.; Flores, C. Genomic Predictors of Asthma Phenotypes and Treatment Response. Front. Pediatr. 2019, 7, 6. [Google Scholar] [CrossRef]
- Perez-Garcia, J.; Herrera-Luis, E.; Lorenzo-Diaz, F.; González, M.; Sardón, O.; Villar, J.; Pino-Yanes, M. Precision Medicine in Childhood Asthma: Omic Studies of Treatment Response. Int. J. Mol. Sci. 2020, 21, 2908. [Google Scholar] [CrossRef]
- Hernandez-Pacheco, N.; Kere, M.; Melén, E. Gene-Environment Interactions in Childhood Asthma Revisited; Expanding the Interaction Concept. Pediatr. Allergy Immunol. Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2022, 33, e13780. [Google Scholar] [CrossRef] [PubMed]
- Sarnowski, C.; Sugier, P.-E.; Granell, R.; Jarvis, D.; Dizier, M.-H.; Ege, M.; Imboden, M.; Laprise, C.; Khusnutdinova, E.K.; Freidin, M.B.; et al. Identification of a New Locus at 16q12 Associated with Time to Asthma Onset. J. Allergy Clin. Immunol. 2016, 138, 1071–1080. [Google Scholar] [CrossRef]
- Pividori, M.; Schoettler, N.; Nicolae, D.L.; Ober, C.; Im, H.K. Shared and Distinct Genetic Risk Factors for Childhood-Onset and Adult-Onset Asthma: Genome-Wide and Transcriptome-Wide Studies. Lancet Respir. Med. 2019, 7, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Bi, W.; Fritsche, L.G.; Mukherjee, B.; Kim, S.; Lee, S. A Fast and Accurate Method for Genome-Wide Time-to-Event Data Analysis and Its Application to UK Biobank. Am. J. Hum. Genet. 2020, 107, 222–233. [Google Scholar] [CrossRef]
- Li, N.; Zhai, Z.; Chen, Y.; Li, X. Transcriptomic and Immunologic Implications of the Epithelial-Mesenchymal Transition Model Reveal a Novel Role of SFTA2 in Prognosis of Non-Small-Cell Lung Carcinoma. Front. Genet. 2022, 13, 911801. [Google Scholar] [CrossRef] [PubMed]
- Shrine, N.; Portelli, M.A.; John, C.; Soler Artigas, M.; Bennett, N.; Hall, R.; Lewis, J.; Henry, A.P.; Billington, C.K.; Ahmad, A.; et al. Moderate-to-Severe Asthma in Individuals of European Ancestry: A Genome-Wide Association Study. Lancet Respir. Med. 2019, 7, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.; Hunkapiller, J.; Bhangale, T.; Reeder, J.; Mukhyala, K.; Tom, J.; Cowgill, A.; Vogel, J.; Forrest, W.F.; Khan, Z.; et al. A Whole Genome Sequencing Study of Moderate to Severe Asthma Identifies a Lung Function Locus Associated with Asthma Risk. Sci. Rep. 2022, 12, 5574. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Gamazon, E.R.; Im, H.K.; Geeleher, P.; White, S.R.; Solway, J.; Clemmer, G.L.; Weiss, S.T.; Tantisira, K.G.; Cox, N.J.; et al. Genome-Wide Interrogation of Longitudinal FEV1 in Children with Asthma. Am. J. Respir. Crit. Care Med. 2014, 190, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hawkins, G.A.; Ampleford, E.J.; Moore, W.C.; Li, H.; Hastie, A.T.; Howard, T.D.; Boushey, H.A.; Busse, W.W.; Calhoun, W.J.; et al. Genome-Wide Association Study Identifies TH1 Pathway Genes Associated with Lung Function in Asthmatic Patients. J. Allergy Clin. Immunol. 2013, 132, 313–320.e15. [Google Scholar] [CrossRef]
- Vonk, J.M.; Nieuwenhuis, M.A.E.; Dijk, F.N.; Boudier, A.; Siroux, V.; Bouzigon, E.; Probst-Hensch, N.; Imboden, M.; Keidel, D.; Sin, D.; et al. Novel Genes and Insights in Complete Asthma Remission: A Genome-Wide Association Study on Clinical and Complete Asthma Remission. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2018, 48, 1286–1296. [Google Scholar] [CrossRef]
- Teumer, A.; Li, Y.; Ghasemi, S.; Prins, B.P.; Wuttke, M.; Hermle, T.; Giri, A.; Sieber, K.B.; Qiu, C.; Kirsten, H.; et al. Genome-Wide Association Meta-Analyses and Fine-Mapping Elucidate Pathways Influencing Albuminuria. Nat. Commun. 2019, 10, 4130. [Google Scholar] [CrossRef]
- Oelsner, E.C.; Balte, P.P.; Grams, M.E.; Cassano, P.A.; Jacobs, D.R.; Barr, R.G.; Burkart, K.M.; Kalhan, R.; Kronmal, R.; Loehr, L.R.; et al. Albuminuria, Lung Function Decline, and Risk of Incident Chronic Obstructive Pulmonary Disease. The NHLBI Pooled Cohorts Study. Am. J. Respir. Crit. Care Med. 2019, 199, 321–332. [Google Scholar] [CrossRef]
- Bolotova, E.V.; Dudnikova, A.V.; Chulkova, A.O. Frequency of Albuminuria in Patients with Bronchial Obstructive Diseases. Adv. Gerontol. Uspekhi Gerontol. 2021, 34, 272–276. [Google Scholar]
- Jackman, J.K.; Stockwell, A.; Choy, D.F.; Xie, M.M.; Lu, P.; Jia, G.; Li, H.; Abbas, A.R.; Bronson, P.G.; Lin, W.-Y.; et al. Genome-Wide Association Study Identifies Kallikrein 5 in Type 2 Inflammation-Low Asthma. J. Allergy Clin. Immunol. 2022, 150, 972–978.e7. [Google Scholar] [CrossRef]
- Kim, J.-H.; Cheong, H.S.; Park, J.S.; Jang, A.-S.; Uh, S.-T.; Kim, Y.-H.; Kim, M.-K.; Choi, I.S.; Cho, S.H.; Choi, B.W.; et al. A Genome-Wide Association Study of Total Serum and Mite-Specific IgEs in Asthma Patients. PLoS ONE 2013, 8, e71958. [Google Scholar] [CrossRef] [PubMed]
- Yick, C.Y.; Zwinderman, A.H.; Kunst, P.W.; Grünberg, K.; Mauad, T.; Fluiter, K.; Bel, E.H.; Lutter, R.; Baas, F.; Sterk, P.J. Glucocorticoid-Induced Changes in Gene Expression of Airway Smooth Muscle in Patients with Asthma. Am. J. Respir. Crit. Care Med. 2013, 187, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Vernet, R.; Matran, R.; Zerimech, F.; Madore, A.-M.; Lavoie, M.-E.; Gagnon, P.-A.; Mohamdi, H.; Margaritte-Jeannin, P.; Siroux, V.; Dizier, M.-H.; et al. Identification of Novel Genes Influencing Eosinophil-Specific Protein Levels in Asthma Families. J. Allergy Clin. Immunol. 2022, 150, 1168–1177. [Google Scholar] [CrossRef]
- Mills, M.C.; Rahal, C. A Scientometric Review of Genome-Wide Association Studies. Commun. Biol. 2019, 2, 9. [Google Scholar] [CrossRef]
- Martin, A.R.; Kanai, M.; Kamatani, Y.; Okada, Y.; Neale, B.M.; Daly, M.J. Clinical Use of Current Polygenic Risk Scores May Exacerbate Health Disparities. Nat. Genet. 2019, 51, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.M.; Gui, H.; Hernandez-Pacheco, N.; Yang, M.; Xiao, S.; Yang, J.J.; Hochstadt, S.; Barczak, A.J.; Eckalbar, W.L.; Rynkowski, D.; et al. Integrative Approach Identifies Corticosteroid Response Variant in Diverse Populations with Asthma. J. Allergy Clin. Immunol. 2019, 143, 1791–1802. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Luis, E.; Mak, A.C.Y.; Perez-Garcia, J.; Martin-Gonzalez, E.; Eng, C.; Beckman, K.B.; Huntsman, S.; Hu, D.; González-Pérez, R.; Hernández-Pérez, J.M.; et al. Admixture Mapping of Severe Asthma Exacerbations in Hispanic/Latino Children and Youth. Thorax 2023, 78, 233–241. [Google Scholar] [CrossRef]
- Spear, M.L.; Hu, D.; Pino-Yanes, M.; Huntsman, S.; Eng, C.; Levin, A.M.; Ortega, V.E.; White, M.J.; McGarry, M.E.; Thakur, N.; et al. A Genome-Wide Association and Admixture Mapping Study of Bronchodilator Drug Response in African Americans with Asthma. Pharmacogenomics J. 2019, 19, 249–259. [Google Scholar] [CrossRef]
- Ortega, V.E.; Daya, M.; Szefler, S.J.; Bleecker, E.R.; Chinchilli, V.M.; Phipatanakul, W.; Mauger, D.; Martinez, F.D.; Herrera-Luis, E.; Pino-Yanes, M.; et al. Pharmacogenetic Studies of Long-Acting β Agonist and Inhaled Corticosteroid Responsiveness in Randomised Controlled Trials of Individuals of African Descent with Asthma. Lancet Child Adolesc. Health 2021, 5, 862–872. [Google Scholar] [CrossRef]
- Gudbjartsson, D.F.; Bjornsdottir, U.S.; Halapi, E.; Helgadottir, A.; Sulem, P.; Jonsdottir, G.M.; Thorleifsson, G.; Helgadottir, H.; Steinthorsdottir, V.; Stefansson, H.; et al. Sequence Variants Affecting Eosinophil Numbers Associate with Asthma and Myocardial Infarction. Nat. Genet. 2009, 41, 342–347. [Google Scholar] [CrossRef]
- Schofield, J.P.R.; Burg, D.; Nicholas, B.; Strazzeri, F.; Brandsma, J.; Staykova, D.; Folisi, C.; Bansal, A.T.; Xian, Y.; Guo, Y.; et al. Stratification of Asthma Phenotypes by Airway Proteomic Signatures. J. Allergy Clin. Immunol. 2019, 144, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Pacheco, N.; Gorenjak, M.; Jurgec, S.; Corrales, A.; Jorgensen, A.; Karimi, L.; Vijverberg, S.J.; Berce, V.; Schieck, M.; Acosta-Herrera, M.; et al. Combined Analysis of Transcriptomic and Genetic Data for the Identification of Loci Involved in Glucocorticosteroid Response in Asthma. Allergy 2021, 76, 1238–1243. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Sharma, A.; Jha, R.; Arora, S.; Ahmad, R.; Rahmani, A.H.; Almatroodi, S.A.; Dohare, R.; Syed, M.A. Transcriptomic Analysis Delineates Potential Signature Genes and MiRNAs Associated with the Pathogenesis of Asthma. Sci. Rep. 2020, 10, 13354. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Zhang, X.; Chen, X.; Brown, A.P.; Weirauch, M.T.; Guilbert, T.W.; Khurana Hershey, G.K.; Biagini, J.M.; Ji, H. Nasal DNA Methylation Differentiates Severe from Non-Severe Asthma in African-American Children. Allergy 2021, 76, 1836–1845. [Google Scholar] [CrossRef]
- Milger, K.; Götschke, J.; Krause, L.; Nathan, P.; Alessandrini, F.; Tufman, A.; Fischer, R.; Bartel, S.; Theis, F.J.; Behr, J.; et al. Identification of a Plasma MiRNA Biomarker Signature for Allergic Asthma: A Translational Approach. Allergy 2017, 72, 1962–1971. [Google Scholar] [CrossRef]
- Jia, G.; Zhong, X.; Im, H.K.; Schoettler, N.; Pividori, M.; Hogarth, D.K.; Sperling, A.I.; White, S.R.; Naureckas, E.T.; Lyttle, C.S.; et al. Discerning Asthma Endotypes through Comorbidity Mapping. Nat. Commun. 2022, 13, 6712. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, M.J.; Zhang, Y.; Durvasula, A.; Inouye, M.; Holmes, C.; Price, A.L.; McVean, G. Age-dependent topic modelling of comorbidities in UK Biobank identifies disease subtypes with differential genetic risk. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Siroux, V.; González, J.R.; Bouzigon, E.; Curjuric, I.; Boudier, A.; Imboden, M.; Anto, J.M.; Gut, I.; Jarvis, D.; Lathrop, M.; et al. Genetic Heterogeneity of Asthma Phenotypes Identified by a Clustering Approach. Eur. Respir. J. 2014, 43, 439–452. [Google Scholar] [CrossRef]
- Dahl, A.; Zaitlen, N. Genetic Influences on Disease Subtypes. Annu. Rev. Genom. Hum. Genet. 2020, 21, 413–435. [Google Scholar] [CrossRef]
- Guo, H.; An, J.; Yu, Z. Identifying Shared Risk Genes for Asthma, Hay Fever, and Eczema by Multi-Trait and Multiomic Association Analyses. Front. Genet. 2020, 11, 270. [Google Scholar] [CrossRef]
- Zhu, Z.; Lee, P.H.; Chaffin, M.D.; Chung, W.; Loh, P.-R.; Lu, Q.; Christiani, D.C.; Liang, L. A Genome-Wide Cross-Trait Analysis from UK Biobank Highlights the Shared Genetic Architecture of Asthma and Allergic Diseases. Nat. Genet. 2018, 50, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Shirai, Y.; Nakanishi, Y.; Suzuki, A.; Konaka, H.; Nishikawa, R.; Sonehara, K.; Namba, S.; Tanaka, H.; Masuda, T.; Yaga, M.; et al. Multi-Trait and Cross-Population Genome-Wide Association Studies across Autoimmune and Allergic Diseases Identify Shared and Distinct Genetic Component. Ann. Rheum. Dis. 2022, 81, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Ziyatdinov, A.; Hobbs, B.D.; Kanaan-Izquierdo, S.; Moll, M.; Sakornsakolpat, P.; Shrine, N.; Chen, J.; Song, K.; Bowler, R.P.; Castaldi, P.J.; et al. Identifying COPD Subtypes Using Multi-Trait Genetics. medRxiv 2023. [Google Scholar] [CrossRef]
- Knapp, E.A.; Kress, A.M.; Parker, C.B.; Page, G.P.; McArthur, K.; Gachigi, K.K.; Alshawabkeh, A.N.; Aschner, J.L.; Bastain, T.M.; Breton, C.V.; et al. The Environmental Influences on Child Health Outcomes (ECHO)-Wide Cohort. Am. J. Epidemiol. 2023, 192, 1249–1263. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, A.H.; Sulieman, L.; Schlueter, D.J.; Halvorson, A.; Qian, J.; Ratsimbazafy, F.; Loperena, R.; Mayo, K.; Basford, M.; Deflaux, N.; et al. The All of Us Research Program: Data quality, utility, and diversity. Patterns 2022, 3, 100570. [Google Scholar] [CrossRef]
- Forno, E.; Wang, T.; Yan, Q.; Brehm, J.; Acosta-Perez, E.; Colon-Semidey, A.; Alvarez, M.; Boutaoui, N.; Cloutier, M.M.; Alcorn, J.F.; et al. A Multiomics Approach to Identify Genes Associated with Childhood Asthma Risk and Morbidity. Am. J. Respir. Cell Mol. Biol. 2017, 57, 439–447. [Google Scholar] [CrossRef]
- Soliai, M.M.; Kato, A.; Helling, B.A.; Stanhope, C.T.; Norton, J.E.; Naughton, K.A.; Klinger, A.I.; Thompson, E.E.; Clay, S.M.; Kim, S.; et al. Multi-omics colocalization with genome-wide association studies reveals a context-specific genetic mechanism at a childhood onset asthma risk locus. Genome Med. 2021, 13, 157. [Google Scholar] [CrossRef]
- Gautam, Y.; Johansson, E.; Mersha, T.B. Multi-Omics Profiling Approach to Asthma: An Evolving Paradigm. J. Pers. Med. 2022, 12, 66. [Google Scholar] [CrossRef]
- Zheng, Z.; Huang, D.; Wang, J.; Zhao, K.; Zhou, Y.; Guo, Z.; Zhai, S.; Xu, H.; Cui, H.; Yao, H.; et al. QTLbase: An integrative resource for quantitative trait loci across multiple human molecular phenotypes. Nucleic Acids Res. 2020, 48, D983–D991. [Google Scholar] [CrossRef]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of Published Genome-Wide Association Studies, Targeted Arrays and Summary Statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef]
- Mills, M.C.; Rahal, C. The GWAS Diversity Monitor Tracks Diversity by Disease in Real Time. Nat. Genet. 2020, 52, 242–243. [Google Scholar] [CrossRef] [PubMed]
- Fantini, D. easyPubMed: Search and Retrieve Scientific Publication Records from PubMed. 2019. R Package Version 2.13. Available online: https://CRAN.R-project.org/package=easyPubMed (accessed on 27 August 2023).
- McLean, C.Y.; Bristor, D.; Hiller, M.; Clarke, S.L.; Schaar, B.T.; Lowe, C.B.; Wenger, A.M.; Bejerano, G. GREAT Improves Functional Interpretation of Cis-Regulatory Regions. Nat. Biotechnol. 2010, 28, 495–501. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein-Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Stark, C.; Breitkreutz, B.-J.; Reguly, T.; Boucher, L.; Breitkreutz, A.; Tyers, M. BioGRID: A General Repository for Interaction Datasets. Nucleic Acids Res. 2006, 34, D535–D539. [Google Scholar] [CrossRef]
- Bader, G.D.; Hogue, C.W.V. An Automated Method for Finding Molecular Complexes in Large Protein Interaction Networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef]




| Endotype | Phenotype | Clinical Features | Molecular Mechanism | References |
|---|---|---|---|---|
| T2-high asthma | Early-onset atopic asthma | Trigger-induced phenotypes. Steroid-sensitive. Preserved lung function | Allergy to aeroallergens | [8,13,14,19,25,26,27,28,29] |
| Late-onset eosinophilic asthma | CRSwNP frequently associated. Steroid-refractory | Staphilococcus aureus enterotoxin | [11,12,30,31,32] | |
| NERD | Samter´s Syndrome. Adult onset. Trigger-induced phenotypes | Arachidonic acid dysregulation | [33,34,35,36,37,38,39,40] | |
| T2-low asthma | Non-atopic asthma | Neutrophilic or paucigranulocytic. | Th1/Th17 inflammation | [22,41,42,43,44,45,46,47,48,49,50,51,52,53,54] |
| Adult onset | ||||
| Smoking-associated asthma | Adult onset. Lower lung function | Oxidative stress | [62,63,64,65,66,67] | |
| Obesity-associated asthma | Metabolic syndrome. Females. Preserved lung function | Th1/Th17 inflammation. Oxidative stress. IL-6 | [55,56,57,58,59,60,61] | |
| Elderly-related asthma | Very late onset. Declined lung function | Th1/Th17 inflammation | [70,71,72,73,74] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espuela-Ortiz, A.; Martin-Gonzalez, E.; Poza-Guedes, P.; González-Pérez, R.; Herrera-Luis, E. Genomics of Treatable Traits in Asthma. Genes 2023, 14, 1824. https://doi.org/10.3390/genes14091824
Espuela-Ortiz A, Martin-Gonzalez E, Poza-Guedes P, González-Pérez R, Herrera-Luis E. Genomics of Treatable Traits in Asthma. Genes. 2023; 14(9):1824. https://doi.org/10.3390/genes14091824
Chicago/Turabian StyleEspuela-Ortiz, Antonio, Elena Martin-Gonzalez, Paloma Poza-Guedes, Ruperto González-Pérez, and Esther Herrera-Luis. 2023. "Genomics of Treatable Traits in Asthma" Genes 14, no. 9: 1824. https://doi.org/10.3390/genes14091824