
Children with Chronic Immune Thrombocytopenia Exhibit High Expression of Human Endogenous Retroviruses TRIM28 and SETDB1
 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Populations
2.2. RNA Extraction and Retrotranscription
2.3. Transcription Levels of Pol Genes of HERV-H, -K, and -W; Env Genes of SYN1, SYN2, and HERV-W; as well as of TRIM28 and SETB1 by Real-Time PCR Assay
2.4. Statistical Analysis
3. Results
3.1. Study Populations
3.2. Expression Levels of Housekeeping Gene
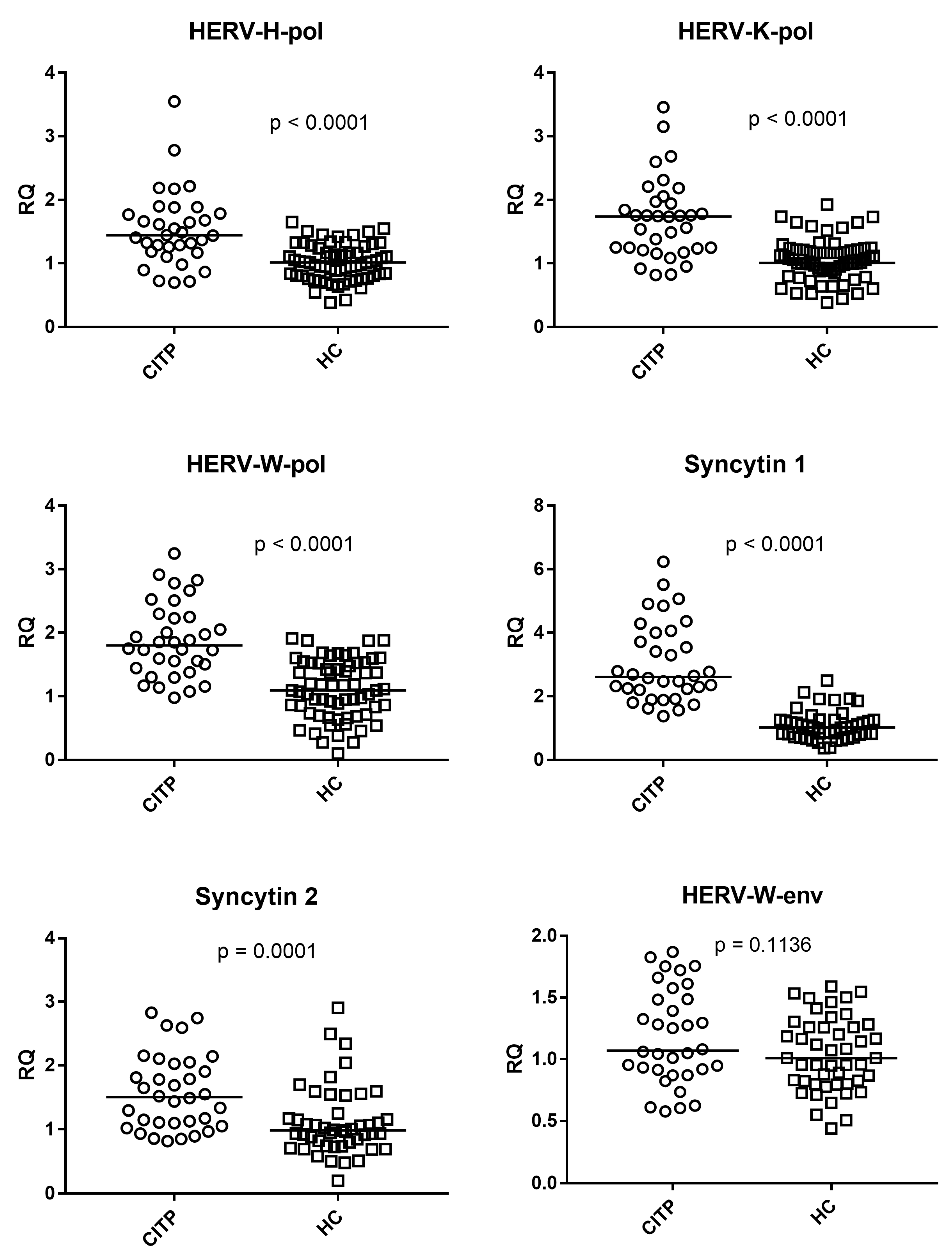
3.3. Expressions of HERV-H-pol, HERV-K-pol, HERV-W-pol, SYN1-env, SYN2-env, and HERV-W-env
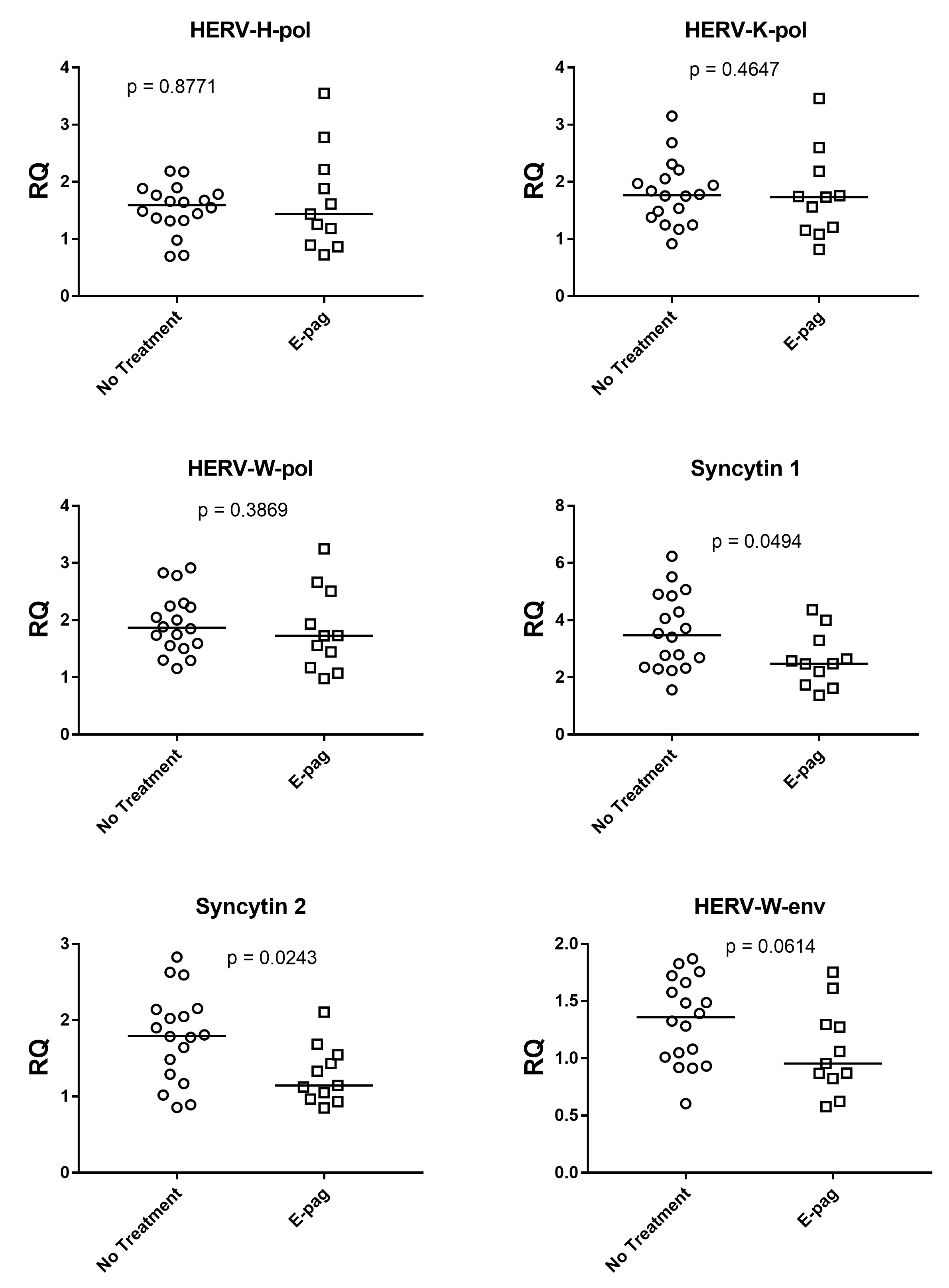
3.4. HERV Expressions in Patients with and without Eltrombopag Treatment
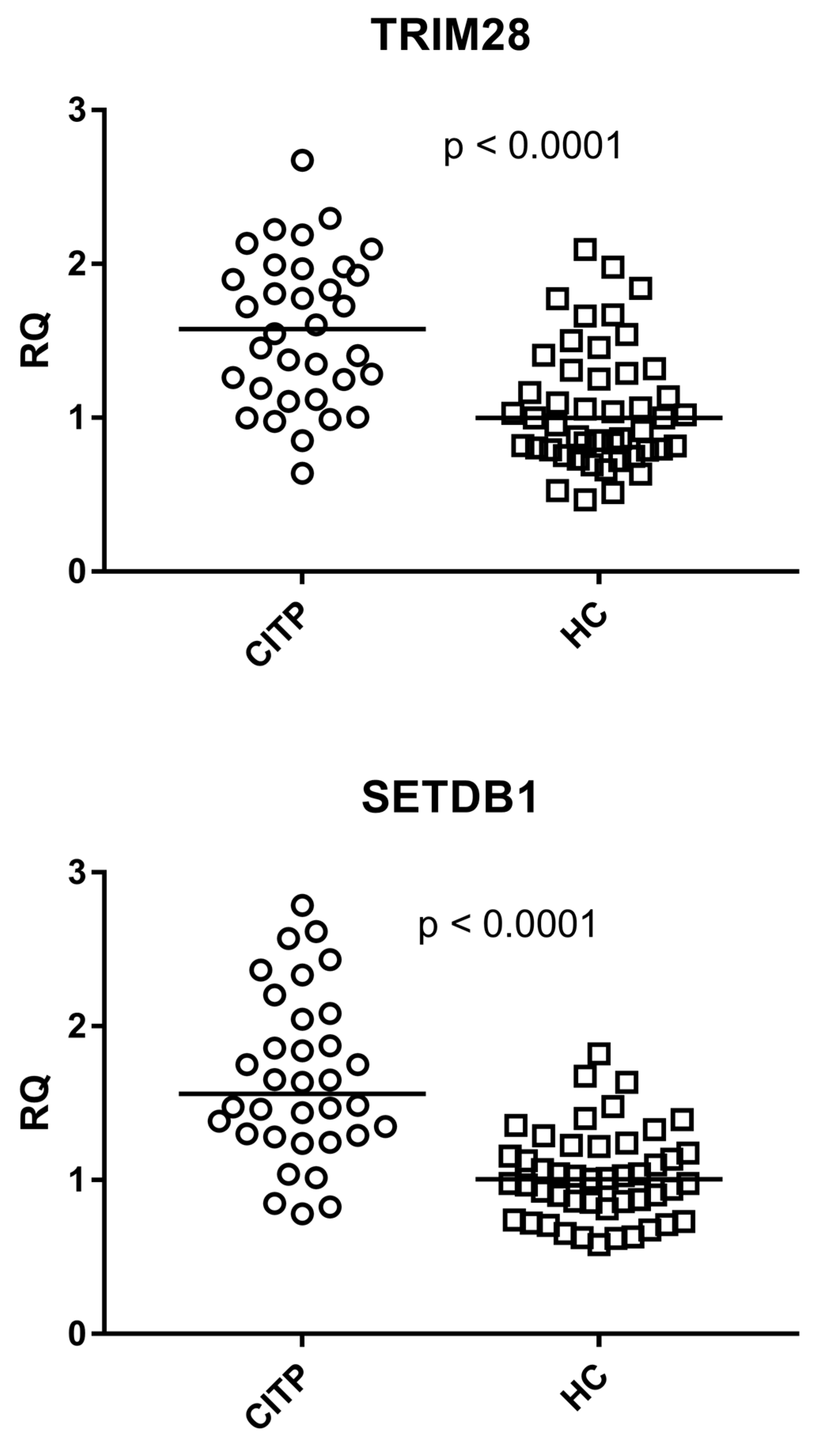
3.5. Expressions of TRIM28 and SETDB1
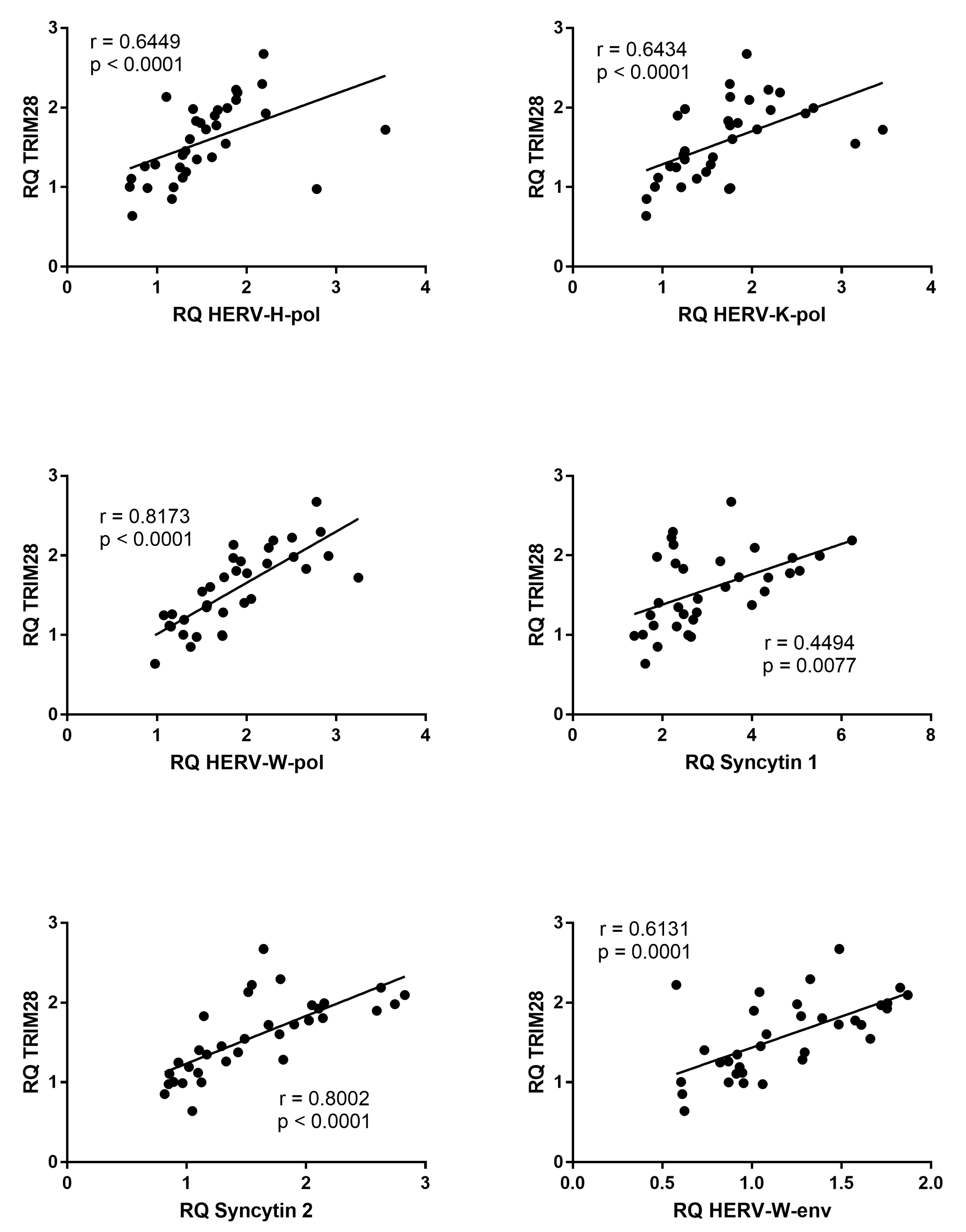
3.6. Correlations between Expressions of TRIM28 and HERVs
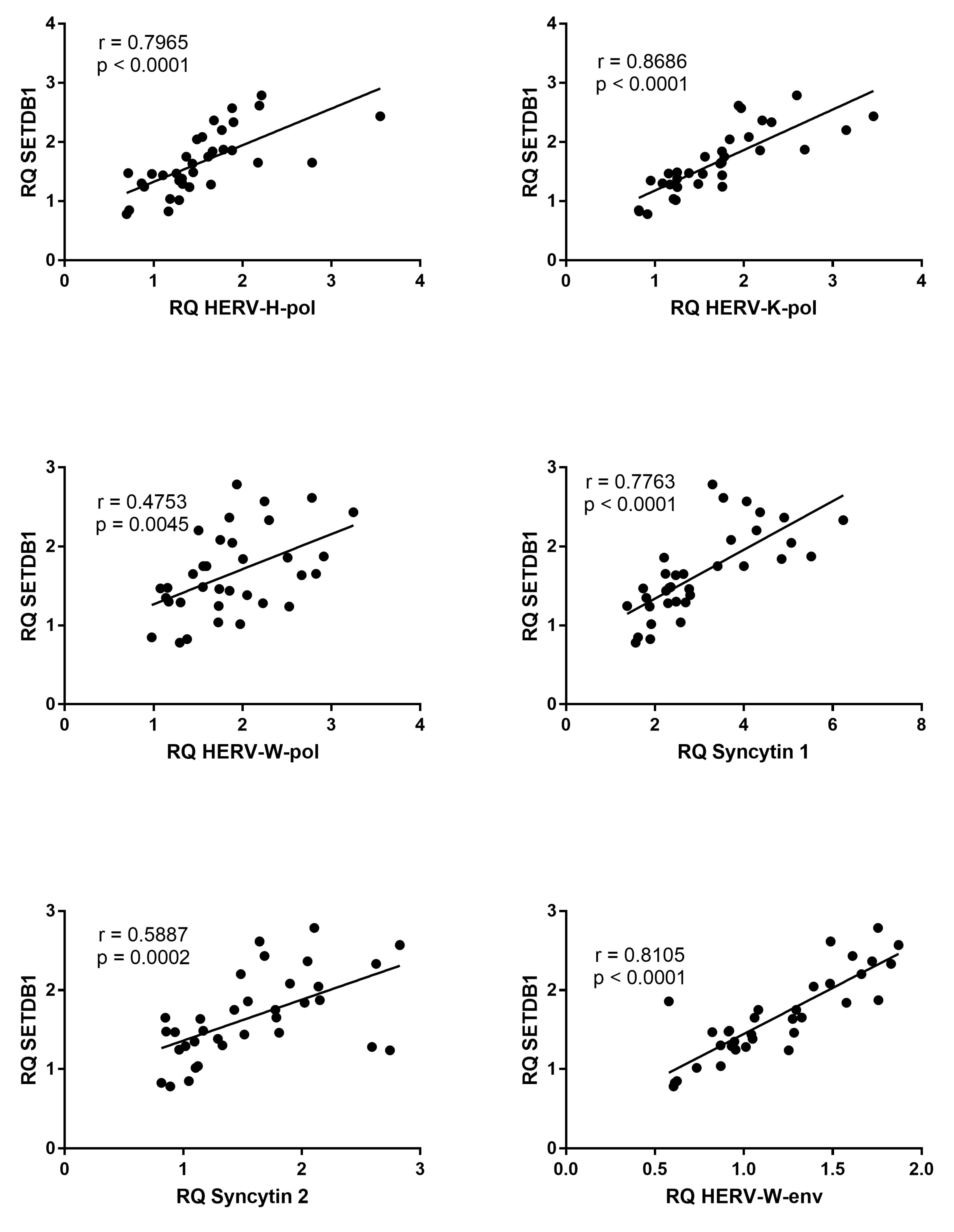
3.7. Correlations between Expressions of SETDB1 and HERVs
3.8. Expressions of TRIM28 and SETDB1 in Patients with and without Eltrombopag Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rodeghiero, F.; Stasi, R.; Gernsheimer, T.; Michel, M.; Provan, D.; Arnold, D.M.; Bussel, J.B.; Cines, D.B.; Chong, B.H.; Cooper, N.; et al. Standardization of terminology, definitions and outcome criteria in immune thrombocytopenic purpura of adults and children: Report from an international working group. Blood 2009, 113, 2386–2393. [Google Scholar] [PubMed]
- Terrell, D.R.; Beebe, L.A.; Vesely, S.K.; Neas, B.R.; Segal, J.B.; George, J.N. The incidence of immune thrombocytopenic purpura in children and adults: A critical review of published reports. Am. J. Hematol. 2010, 85, 174–180. [Google Scholar]
- Provan, D.; Stasi, R.; Newland, A.C.; Blanchette, V.S.; Bolton-maggs, P.; Bussel, J.B.; Chong, H.; Cines, D.B.; Gernsheimer, T.B.; Godeau, B.; et al. Review article International consensus report on the investigation and management of primary immune thrombocytopenia. Blood 2010, 115, 168–186. [Google Scholar] [CrossRef] [PubMed]
- Imbach, P.; Kuhne, T.; Muller, D.; Berchtold, W.; Zimmerman, S.; Elalfy, M.; Buchanan, G.R. ITP: 12 months follow-up data from the prospective registry I of the Intercontinental Childhood ITP Study Group (ICIS). Pediatr. Blood Cancer 2006, 46, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Glanz, J.; France, E.; Xu, S.; Hayes, T.; Hambidge, S. A population-based, multisite cohort study of the predictors of chronic idiopathic thrombocytopenic purpura in children. Pediatrics 2008, 121, e506–e512. [Google Scholar] [CrossRef]
- Schmidt, D.E.; Wendtland Edslev, P.; Heitink-Pollé, K.M.J.; Mertens, B.; Bruin, M.C.A.; Kapur, R.; Vidarsson, G.; van der Schoot, C.E.; Porcelijn, L.; van der Bom, J.G.; et al. A clinical prediction score for transient versus persistent childhood immune thrombocytopenia. J. Thromb. Haemost. 2021, 19, 121–130. [Google Scholar] [CrossRef]
- Cines, D.B.; Bussel, J.B.; Liebman, H.A.; Luning Prak, E.T. The ITP syndrome: Pathogenic and clinical diversity. Blood 2009, 113, 6511–6521. [Google Scholar] [CrossRef] [Green Version]
- Provan, D.; Semple, J.W. Recent advances in the mechanisms and treatment of immune thrombocytopenia. EBioMedicine 2022, 76, 103820. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, X.; Li, H.; Xue, F.; Zhang, L.; Yang, R. Association of FOXP3 gene polymorphisms with chronic immune thrombocytopenia in a Chinese Han population. Int. J. Lab. Hematol. 2021, 43, 1104–1109. [Google Scholar] [CrossRef]
- Swikels, M.; Rijkers, M.; Voorberg, J.; Vidarsson, G.; Leebeek, F.W.G.; Jansen, A.J.G. Emerging concepts in immune thrombocytopenia. Front. Immunol. 2018, 9, 880. [Google Scholar] [CrossRef]
- Zhao, Y.; Cui, S.; Wang, Y.; Xu, R. The extensive regulation of microRNA in immune thrombocytopenia. Clin. Appl. Thromb. Hemost. 2022, 28, 10760296221093595. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Xue, F.; Xu, J.; Fang, Z. Aberrant histone methylation in the patients with immune thrombocytopenia. Platelets 2014, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Gkoutsias, A.; Makis, A. The role of epigenetics in childhood autoimmune diseases with hematological manifestations. Pediatr. Investig. 2022, 6, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.E. Origins and evolutionary consequences of ancient endogenous retroviruses. Nat. Rev. Microbiol. 2019, 17, 355–370. [Google Scholar] [CrossRef]
- Grow, E.J.; Flynn, R.A.; Chavez, S.L.; Bayless, N.L.; Wossidlo, M.; Wesche, D.J.; Martin, L.; Ware, C.B.; Blish, C.A.; Chang, H.Y.; et al. Intrinsic retroviral reactivation in human preimplantation embryos and pluripotent cells. Nature 2015, 52, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Heidmann, O.; Béguin, A.; Paternina, J.; Berthier, R.; Deloger, M.; Bawa, O.; Heidmann, T. HEMO, an ancestral endogenous retroviral envelope protein shed in the blood of pregnant women and expressed in pluripotent stem cells and tumors. Proc. Natl. Acad. Sci. USA 2017, 114, E6642–E6651. [Google Scholar] [CrossRef]
- Blond, J.L.; Lavillette, D.; Cheynet, V.; Bouton, O.; Oriol, G.; Chapel-Fernandes, S.; Mandrand, B.; Mallet, F.; Cosset, F.L. An envelope glycoprotein of the human endogenous HERV-W is expressed in the human placenta and fuses cells expressing the type D mammalian retrovirus receptor. J. Virol. 2000, 74, 3321–3329. [Google Scholar] [CrossRef] [Green Version]
- Blaise, L.S.; de Parseval, N.; Benit, L.; Heidmann, T. Genomewide screening for fusogenic human endogenous retrovirus envelopes identifies syncytin 2, a gene conserved on primate evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 13013–13018. [Google Scholar] [CrossRef]
- Garcia-Montojo, M.; Rodriguez-Martin, E.; Ramos-Mozo, P.; Ortega-Madueño, I.; Dominguez-Mozo, M.I.; Arias-Leal, A.; García-Martínez, M.Á.; Casanova, I.; Galan, V.; Arroyo, R.; et al. Syncytin-1/HERV-W envelope is an early activation marker of leukocytes and is upregulated in multiple sclerosis patients. Eur. J. Immunol. 2020, 50, 685–694. [Google Scholar] [CrossRef]
- Lokossou, A.G.; Toudic, C.; Nguyen, P.T.; Elisseeff, X.; Vargas, A.; Rassart, É.; Lafond, J.; Leduc, L.; Bourgault, S.; Gilbert, C.; et al. Endogenous retrovirus-encoded Syncytin-2 contributes to exosome-mediated immunosuppression of T cells. Biol. Reprod. 2020, 102, 185–198. [Google Scholar] [CrossRef]
- Ukadike, K.C.; Mustelin, T. Implications of endogenous retroelements in the etiopathogenesis of systemic lupus erythematosus. J. Clin. Med. 2021, 10, 856. [Google Scholar] [CrossRef] [PubMed]
- Perron, H.; Dougier-Reynaud, H.L.; Lomparski, C.; Popa, I.; Firouzi, R.; Bertrand, J.B.; Marusic, S.; Portoukalian, J.; Jouvin-Marche, E.; Villiers, C.L.; et al. Human endogenous retrovirus protein activates innate immunity and promotes experimental allergic encephalomyelitis in mice. PLoS ONE 2013, 8, e80128. [Google Scholar]
- Tovo, P.A.; Rabbone, I.; Tinti, D.; Galliano, I.; Trada, M.; Daprà, V.; Cerutti, F.; Bergallo, M. Enhanced expression of human endogenous retroviruses in new-onset type 1 diabetes: Potential pathogenetic and therapeutic implications. Autoimmunity 2020, 53, 283–288. [Google Scholar] [PubMed]
- Tovo, P.A.; Opramolla, A.; Pizzol, A.; Calosso, G.; Daprà, V.; Galliano, I.; Calvi, C.; Pinon, M.; Cisarò, F.; Rigazio, C.; et al. Overexpression of endogenous retroviruses in children with celiac disease. Eur. J. Pediatr. 2021, 180, 2429–2434. [Google Scholar] [CrossRef]
- Isbel, L.; Whitelaw, E. Endogenous retroviruses in mammals: An emerging picture of how ERVs modify expression of adjacent genes. Bioessays 2012, 34, 734–738. [Google Scholar] [PubMed]
- Chuong, E.B.; Elde, N.C.; Feschotte, C. Regulatory evolution of innate immunity through co-option of endogenous retroviruses. Science 2016, 351, 1083–1087. [Google Scholar] [CrossRef] [Green Version]
- Rolland, A.; Jouvin-Marche, E.; Viret, C.; Faure, M.; Perron, H.; Marche, P.N. The envelope protein of a human endogenous retrovirus-W family activates innate immunity through CD14/TLR4 and promotes Th1-like responses. J. Immunol. 2006, 176, 7636–7644. [Google Scholar] [CrossRef]
- Dembny, P.; Newman, A.G.; Singh, M.; Hinz, M.; Szczepek, M.; Krüger, C.; Adalbert, R.; Dzaye, O.; Trimbuch, T.; Wallach, T.; et al. Human endogenous retrovirus HERV-K(HML-2) RNA causes neurodegeneration through Toll-like receptors. JCI Insight 2020, 5, e131093. [Google Scholar] [CrossRef] [Green Version]
- Brudek, T.; Christensen, T.; Aagaard, L.; Petersen, T.; Hansen, H.J.; Møller-Larsen, A. B cells and monocytes from patients with active multiple sclerosis exhibit increased surface expression of both HERV-H env and HERV-W env, accompanied by increased seroreactivity. Retrovirology 2009, 6, 104. [Google Scholar] [CrossRef] [Green Version]
- Nelson, P.N.; Roden, D.; Nevill, A.; Freimanis, G.L.; Trela, M.; Ejtehadi, H.D.; Bowman, S.; Axford, J.; Veitch, A.M.; Tugnet, N.; et al. Rheumatoid arthritis is associated with IgG antibodies to human endogenous retrovirus gag matrix: A potential pathogenic mechanism of disease? J. Rheumatol. 2014, 41, 1952–1960. [Google Scholar]
- Mameli, G.; Erre, G.L.; Caggiu, E.; Mura, S.; Cossu, D.; Bo, M.; Cadoni, M.L.; Piras, A.; Mundula, N.; Colombo, E.; et al. Identification of a HERV-K env surface peptide highly recognized in rheumatoid arthritis (RA) patients: A cross-sectional case–control study. Clin. Exp. Immunol. 2017, 189, 127–131. [Google Scholar] [PubMed] [Green Version]
- Perl, A.; Nagy, G.; Koncz, A.; Gergely, P.; Fernandez, D.; Doherty, E.; Telarico, T.; Bonilla, E.; Phillips, P.E. Molecular mimicry and immunomodulation by the HRES-1 endogenous retrovirus in SLE. Autoimmunity 2008, 41, 287–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tovo, P.A.; Garazzino, S.; Daprà, V.; Alliaudi, C.; Silvestro, E.; Calvi, C.; Montanari, P.; Galliano, I.; Bergallo, M. Chronic HCV infection is associated with overexpression of human endogenous retroviruses that persists after drug-induced viral clearance. Int. J. Mol. Sci. 2020, 21, 3980. [Google Scholar]
- Tovo, P.A.; Garazzino, S.; Daprà, V.; Pruccoli, G.; Calvi, C.; Mignone, F.; Alliaudi, C.; Denina, M.; Scolfaro, C.; Zoppo, M.; et al. COVID-19 in children: Expressions of type I/II/III interferons, TRIM28, SETDB1, and endogenous retroviruses in mild and severe cases. Int. J. Mol. Sci. 2021, 22, 7481. [Google Scholar]
- Friedman, J.R.; Fredericks, W.J.; Jensen, D.E.; Speicher, D.W.; Huang, X.P.; Neilson, E.G.; Rauscher, F.J. 3rd. KAP-1, a novel corepressor for the highly conserved KRAB repression domain. Genes Dev. 1996, 10, 2067–2078. [Google Scholar] [CrossRef] [Green Version]
- Sobocinska, J.; Molenda, S.; Machnik, M.; Oleksiewicz, U. KRAB-ZFP transcriptional regulators acting as oncogenes and tumor suppressors: An Overview. Int. J. Mol. Sci. 2021, 23, 2212. [Google Scholar]
- Schultz, D.C.; Ayyanathan, K.; Negorev, D.; Maul, G.G.; Rauscher, F.J. 3rd. SETDB1: A novel KAP-1-associated histone H3, lysine 9-specific methyltransferase that contributes to HP1-mediated silencing of euchromatic genes by KRAB zinc-finger proteins. Genes Dev. 2002, 16, 919–932. [Google Scholar] [PubMed] [Green Version]
- Turelli, P.; Castro-Diaz, N.; Marzetta, F.; Kapopoulou, A.; Raclot, C.; Duc, J.; Tieng, V.; Quenneville, S.; Trono, D. Interplay of TRIM28 and DNA methylation in controlling human endogenous retroelements. Genome Res. 2014, 24, 1260–1270. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Liu, Y.; Lu, H.; Sun, S.C.; Jin, W.; Wang, X.; Dong, C. Epigenetic activation during T helper 17 cell differentiation is mediated by Tripartite motif containing 28. Nat. Commun. 2018, 12, 1424. [Google Scholar] [CrossRef] [Green Version]
- Gehrmann, U.; Carpier, J.M.; Burgdorf, N.; Hoyler, T.; Suarez, G.; Joannas, L.; Heurtebise-Chrétien, S.; Durand, S.; Panes, R.; Bellemare-Pelletier, A.; et al. Critical role for TRIM28 and HP1/in the epigenetic control of T cell metabolic reprograming and effector differentiation. Proc. Natl. Acad. Sci. USA 2019, 116, 25839–25849. [Google Scholar] [CrossRef] [Green Version]
- Adoue, V.; Binet, B.; Malbec, A.; Fourquet, J.; Romagnoli, P.; van Meerwijk, J.P.M.; Amigorena, S.; Joffre, O.P. The histone methyltransferase SETDB1 controls T helper cell lineage integrity by repressing endogenous retroviruses. Immunity 2019, 50, 629–644. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgenb, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, I.; Fuscaldo, A.A.; Erlick, B.J.; Kingsbury, E.W.; Schultz, G.M.; Fuscaldo, K.E. Analysis of platelets from patients with thrombocythemia for reverse transcriptase and virus-like particles. J. Natl. Cancer Inst. 1975, 55, 1069. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, I.; Fuscaldo, A.A.; Erlick, B.J.; Fuscaldo, K.E. Effect of busulfan on oncornavirus-like activity in platelets and chromosomes in polycythemia vera and essential thrombocythemia. J. Natl. Cancer Inst. 1977, 59, 61. [Google Scholar] [CrossRef]
- Boyd, M.T.; Foley, B.; Brodsky, I. Evidence for copurification of HERV-K-related transcripts and a reverse transcriptase activity in human platelets from patients with essential thrombocythemia. Blood 1997, 90, 4022–4030. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.; Brodsky, I. Human endogenous retrovirus (HERV-K)eg particles in megakaryocytes cultured from essential thromocythemia peripheral blood stem cells. Exp. Hematol. 2004, 32, 520–525. [Google Scholar] [CrossRef]
- Miyazawa, M.; Tabata, N.; Fujisawa, R.; Hashimoto, K.; Shiwaku, H.; Takei, Y.A. Roles of endogenous retroviruses and platelets in the development of vascular injury in spontaneous mouse models of autoimmune diseases. Int. J. Cardiol. 2000, 75 (Suppl. 1), S65–S73. [Google Scholar] [CrossRef]
- Tovo, P.A.; Monti, G.; Daprà, V.; Montanari, P.; Calvi, C.; Alliaudi, C.; Sardo, A.; Galliano, I.; Bergallo, M. Enhanced expression of endogenous retroviruses and of TRIM28 and SETDB1 in children with food allergy. Clin. Transl. Allergy 2022, 12, e12124. [Google Scholar] [CrossRef]
- Matsui, T.; Leung, D.; Miyashita, H.; Maksakova, I.A.; Miyachi, H.; Kimura, H.; Tachibana, M.; Lorincz, M.C.; Shinkai, Y. Proviral silencing in embryonic stem cells requires the histone methyltransferase ESET. Nature 2010, 464, 927–931. [Google Scholar] [CrossRef] [Green Version]
- Rowe, H.M.; Kapopoulou, A.; Corsinotti, A.; Fasching, L.; Macfarlan, T.; Tarabay, Y.; Viville, S.; Jakobsson, J.; Pfaff, S.L.; Trono, D. TRIM28 repression of retrotransposon-based enhancers is necessary to preserve transcriptional dynamics in embryonic stem cells. Genome Res. 2013, 23, 452–461. [Google Scholar] [CrossRef] [Green Version]
- Wiznerowicz, M.; Jakobsson, J.; Szulc, J.; Liao, S.; Quazzola, A.; Beermann, F.; Aebischer, P.; Trono, D. The Kruppel-associated box repressor domain can trigger de novo promoter methylation during mouse early embryogenesis. J. Biol. Chem. 2007, 282, 34535–34541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoni de Sio, F.R.; Barde, I.; Offner, S.; Kapopoulou, A.; Corsinotti, A.; Bojkowska, K.; Genolet, R.; Thomas, J.H.; Luescher, I.F.; Pinschewer, D.; et al. KAP1 regulates gene networks controlling T-cell development and responsiveness. FASEB J. 2012, 26, 4561–4575. [Google Scholar]
- Randolph, K.; Hyder, U.; D’Orso, I. KAP1/TRIM28: Transcriptional activator and/or repressor of viral and cellular programs? Front. Cell Infect. Microbiol. 2022, 12, 834636. [Google Scholar]
- Chabert, A.; Hamzeh-Cognasse, H.; Pozzetto, B.; Cognasse, F.; Schattner, M.; Gomez, R.M.; Garraud, O. Human platelets and their capacity of binding viruses: Meaning and challenges? BMC Immunol. 2015, 16, 26. [Google Scholar]
- Raadsen, M.; Du Toit, J.; Langerak, T.; van Bussel, B.; van Gorp, E.; Goeijenbier, M. Thrombocytopenia in virus infections. J. Clin. Med. 2021, 10, 877. [Google Scholar] [CrossRef]
- Mameli, G.; Poddighe, L.; Mei, A.; Uleri, E.; Sotgiu, S.; Serra, C.; Manetti, R.; Dolei, A. Expression and activation by Epstein Barr virus of human endogenous retroviruses-W in blood cells and astrocytes: Inference for multiple sclerosis. PLoS ONE 2012, 7, e44991. [Google Scholar]
- Assinger, A.; Yaiw, K.C.; Göttesdorfer, I.; Leib-Mösch, C.; Söderberg-Nauclér, C. Human cytomegalovirus (HCMV) induces human endogenous retrovirus (HERV) transcription. Retrovirology 2013, 10, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Kuyl, A.C. HIV infection and HERV expression: A review. Retrovirology 2012, 9, 6. [Google Scholar]
- Liu, H.; Bergant, V.; Frishman, G.; Ruepp, A.; Pichlmair, A.; Vincendeau, M.; Frishman, D. Influenza A virus infection reactivates human endogenous retroviruses associated with modulation of antiviral immunity. Viruses 2022, 14, 1591. [Google Scholar] [CrossRef]
- Marston, J.L.; Greenig, M.; Singh, M.; Bendall, M.L.; Duarte, R.R.R.; Feschotte, C.; Iñiguez, L.P.; Nixon, D.F. SARS-CoV-2 infection mediates differential expression of human endogenous retroviruses and long interspersed nuclear elements. JCI Insight 2021, 6, e147170. [Google Scholar] [CrossRef]
- Manghera, M.; Ferguson-Parry, J.; Lin, R.; Douville, R.N. NF-κB and IRF1 induce endogenous retrovirus K expression via interferon-stimulated response elements in its 5' long terminal repeat. J. Virol. 2016, 90, 9338–9349. [Google Scholar] [CrossRef] [Green Version]
- Khodadi, E.; Asnafi, A.A.; Shahrabi, S.; Shahjahani, M.; Saki, N. Bone marrow niche in immune thrombocytopenia: A focus on megakaryopoiesis. Ann. Hematol. 2016, 95, 1765–1776. [Google Scholar] [CrossRef]
- Ito, K.; Baudino, L.; Kihara, M.; Leroy, V.; Vyse, T.J.; Evans, L.H.; Izui, S. Three Sgp loci act independently as well as synergistically to elevate the expression of specific endogenous retroviruses implicated in murine lupus. J. Autoimmun. 2013, 43, 10–17. [Google Scholar]
- Bashratyan, R.; Regn, D.; Rahman, M.J.; Marquardt, K.; Fink, E.; Hu, W.Y.; Elder, J.H.; Binley, J.; Sherman, L.A.; Dai, Y.D. Type 1 diabetes pathogenesis is modulated by spontaneous autoimmune responses to endogenous retrovirus antigens in NOD mice. Eur. J. Immunol. 2017, 47, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Freimanis, G.; Hooley, P.; Ejtehadi, H.D.; Ali, H.A.; Veitch, A.; Rylance, P.B.; Alawi, A.; Axford, J.; Nevill, A.; Murray, P.G.; et al. A role for human endogenous retrovirus-K (HML-2) in rheumatoid arthritis: Investigating mechanisms of pathogenesis. Clin. Exp. Immunol. 2010, 160, 340–347. [Google Scholar]
- Sicat, J.; Sutkowski, N.; Huber, B.T. Expression of human endogenous retrovirus HERV-K18 superantigen is elevated in juvenile rheumatoid arthritis. J. Rheumatol. 2005, 32, 1821–1831. [Google Scholar]
- Tolosa, J.M.; Parsons, K.S.; Hansbro, P.M.; Smith, R.; Wark, P.A. The placental protein syncytin-1 impairs antiviral responses and exaggerates inflammatory responses to influenza. PLoS ONE 2015, 10, e0118629. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Z.; Wang, P.; Li, S.; Zeng, J.; Tu, X.; Yan, Q.; Xiao, Z.; Pan, M.; Zhu, F. Syncytin-1, an endogenous retroviral protein, triggers the activation of CRP via TLR3 signal cascade in glial cells. Brain Behav. Immun. 2018, 67, 324–334. [Google Scholar] [CrossRef]
- Bussel, J.B.; Cheng, G.; Saleh, M.N.; Psaila, B.; Kovaleva, L.; Meddeb, B.; Kloczko, J.; Hassani, H.; Mayer, B.; Stone, N.L.; et al. Eltrombopag for the treatment of chronic idiopathic thrombocytopenic purpura. N. Engl. J. Med. 2007, 357, 2237–2247. [Google Scholar] [CrossRef]
- Giordano, P.; Lassandro, G.; Barone, A.; Cesaro, S.; Fotzi, I.; Giona, F.; Ladogana, S.; Miano, M.; Marzollo, A.; Nardi, M.; et al. Use of Eltrombopag in children with chronic immune thrombocytopenia (ITP): A real life retrospective multicenter experience of the Italian Association of Pediatric Hematology and Oncology (AIEOP). Front. Med. 2020, 7, 66. [Google Scholar]
- Liang, Q.; Deng, H.; Li, X.; Wu, X.; Tang, Q.; Chang, T.H.; Peng, H.; Rauscher, F.J., 3rd; Ozato, K.; Zhu, F. Tripartite motif-containing protein 28 is a small ubiquitin-related modifier E3 ligase and negative regulator of IFN regulatory factor 7. J. Immunol. 2011, 187, 4754–4763. [Google Scholar]
- Krischuns, T.; Günl, F.; Henschel, L.; Binder, M.; Willemsen, J.; Schloer, S.; Rescher, U.; Gerlt, V.; Zimmer, G.; Nordhoff, C.; et al. Phosphorylation of TRIM28 enhances the expression of IFN-β and proinflammatory cytokines during HPAIV infection of human lung epithelial cells. Front. Immunol. 2018, 9, 2229. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.F.; Yu, J.; Chang, M.; Zhang, M.; Zhou, D.; Cammas, F.; Sun, S.C. TRIM28 mediates chromatin modifications at the TCRα enhancer and regulates the development of T and natural killer T cells. Proc. Natl. Acad. Sci. USA 2012, 109, 20083–20088. [Google Scholar] [CrossRef]
- Takikita, S.; Muro, R.; Takai, T.; Otsubo, T.; Kawamura, Y.I.; Dohi, T.; Oda, H.; Kitajima, M.; Oshima, K.; Hattori, M.; et al. A Histone methyltransferase ESET is critical for T cell development. J. Immunol. 2016, 197, 2269–2279. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Martin, S.; Pfleger, C.; Du, J.; Buckner, J.H.; Bluestone, J.A.; Riley, J.L.; Ziegler, S.F. Cutting Edge: A novel, human-specific interacting protein couples FOXP3 to a chromatin-remodeling complex that contains KAP1/TRIM28. J. Immunol. 2013, 190, 4470–4473. [Google Scholar]
- Chikuma, S.; Yamanaka, S.; Nakagawa, S.; Ueda, M.T.; Hayabuchi, H.; Tokifuji, Y.; Kanayama, M.; Okamura, T.; Arase, H.; Yoshimura, A. TRIM28 expression on dendritic cells prevents excessive T cell priming by silencing endogenous retrovirus. J. Immunol. 2021, 206, 1528–1539. [Google Scholar] [CrossRef]
- Hartung, H.P.; Derfuss, T.; Cree, B.A.; Sormani, M.P.; Selmaj, K.; Stutters, J.; Prados, F.; MacManus, D.; Schneble, H.M.; Lambert, E.; et al. Efficacy and safety of temelimab in multiple sclerosis: Results of a randomized phase 2b and extension study. Mult. Scler. 2021, 9, 422–440. [Google Scholar]
- Giménez-Orenga, K.; Oltra, E. Human endogenous retrovirus as therapeutic targets in neurologic disease. Pharmaceuticals 2021, 14, 495. [Google Scholar] [CrossRef]
- Baldwin, E.T.; Götte, M.; Tchesnokov, E.P.; Arnold, E.; Hagel, M.; Nichols, C.; Dossang, P.; Lamers, M.; Wan, P.; Steinbacher, S.; et al. Human endogenous retrovirus-K (HERV-K) reverse transcriptase (RT) structure and biochemistry reveals remarkable similarities to HIV-1 RT and opportunities for HERV-K-specific inhibition. Proc. Natl. Acad. Sci. USA 2022, 119, e2200260119. [Google Scholar]
- Laderoute, M.P.; Giulivi, A.; Larocque, L.; Bellfoy, D.; Hou, Y.; Wu, H.X.; Fowke, K.; Wu, J.; Diaz-Mitoma, F. The replicative activity of human endogenous retrovirus K102 (HERV-K102) with HIV viremia. AIDS 2007, 21, 2417–2424. [Google Scholar] [CrossRef]
- Bowen, L.N.; Tyagi, R.; Li, W.; Alfahad, T.; Smith, B.; Wright, M.; Singer, E.J.; Nath, A. HIV-associated motor neuron disease: HERV-K activation and response to antiretroviral therapy. Neurology 2016, 87, 1756–1762. [Google Scholar]
- Gentile, I.; Bonadies, G.; Buonomo, A.R.; Minei, G.; Borrelli, F.; Foggia, M.; Chiurazzi, F.; Borgia, G. Resolution of autoimmune thrombocytopenia associated with raltegravir use in an HIV-positive patient. Platelets 2013, 2, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Montojo, M.; Fathi, S.; Norato, G.; Smith, B.R.; Rowe, D.B.; Kiernan, M.C.; Vucic, S.; Mathers, S.; van Eijk, R.P.A.; Santamaria, U.; et al. Inhibition of HERV-K (HML-2) in amyotrophic lateral sclerosis patients on antiretroviral therapy. J. Neurol. Sci. 2021, 423, 17358. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, L.; Zhang, Y.; Nakata, Y.; Chan, H.L.; Morey, L. Epigenetic mechanisms in breast cancer therapy and resistance. Nat. Commun. 2021, 12, 786. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer/ Probe | Sequence |
|---|---|---|
| HERV-H pol | Forward | 5′-TGGACTGTGCTGCCGCAA-3′ |
| Reverse | 5′-GAAGSTCATCAATATATTGAATAAGGTGAGA-3′ | |
| Probe | 6FAM-5′-TTCAGGGACAGCCCTCGTTACTTCAGCCAAGCTC-3′-TAMRA | |
| HERV-K pol | Forward | 5′-CCACTGTAGAGCCTCCTAAACCC-3′ |
| Reverse | 5′-TTGGTAGCGGCCACTGATTT-3′ | |
| Probe | 6FAM-5′-CCCACACCGGTTTTTCTGTTTTCCAAGTTAA-3′-TAMRA | |
| HERV-W pol | Forward | 5′-ACMTGGAYKRTYTTRCCCCAA-3′ |
| Reverse | 5′-GTAAATCATCCACMTAYYGAAGGAYMA-3′ | |
| Probe | 6FAM-5′-TYAGGGATAGCCCYCATCTRTTTGGYCAGGCA-3′-TAMRA | |
| Syncytin 1 env | Forward | 5′-ACTTTGTCTCTTCCAGAATCG-3′ |
| Reverse | 5′-GCGGTAGATCTTAGTCTTGG-3′ | |
| Probe | 6FAM-5′-TGCATCTTGGGCTCCAT-3′-TAMRA | |
| Syncytin 2 env | Forward | 5′-GCCTGCAAATAGTCTTCTTT-3′ |
| Reverse | 5′-ATAGGGGCTATTCCCATTAG-3′ | |
| Probe | 6FAM- 5′-TGATATCCGCCAGAAACCTCCC-3′-TAMRA | |
| HERV-W env | Forward | 5′-CTTCCAGAATTGAAGCTGTAAAGC-3′ |
| Reverse | 5′-GGGTTGTGCAGTTGAGATTTCC-3′ | |
| Probe | 6FAM-5′-TTCTTCAAATGGAGCCCCAGATGCAG-3′-TAMRA | |
| TRIM28 | Forward | 5′-GCCTCTGTGTGAGACCTGTGTAGA-3′ |
| Reverse | 5′-CCAGTAGAGCGCACAGTATGGT-3′ | |
| Probe | 6FAM-5′-CGCACCAGCGGGTGAAGTACACC-3′-TAMRA | |
| SETDB1 | Forward | 5′-GCCGTGACTTCATAGAGGAGTATGT-3′ |
| Reverse | 5′-GCTGGCCACTCTTGAGCAGTA-3′ | |
| Probe | 6FAM-5′-TGCCTACCCCAACCGCCCCAT-3′-TAMRA | |
| GAPDH | Forward | 5′-CGAGATCCCTCCAAAATCAA-3′ |
| Reverse | 5′-TTCACACCCATGACGAACAT-3′ | |
| Probe | 6FAM-5′-TCCAACGCAAAGCAATACATGAAC-3′-TAMRA |
| CITP (n = 34) | B1 (n = 64) | B2 (n = 47) | |
|---|---|---|---|
| Mean age (±SD) | 12.1 (±4.1) years | 10.9 (±3.5) years | 10.6 (±3.2) years |
| Males (%) | 17 (50) | 35 (55.7) | 29 (61.7) |
| Mean duration of CITP (±SD) at time of sampling | 4.4 (±3.1) years | ||
| Ongoing therapy n (%) | Eltrombopag: 11 (31.4) MMF: 2 (5.7) Eltrombopag + MMF: 2 (5.7) Sirolimus: 1 (2.9) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tovo, P.-A.; Galliano, I.; Parodi, E.; Calvi, C.; Gambarino, S.; Licciardi, F.; Dini, M.; Montanari, P.; Branca, M.; Ramenghi, U.; et al. Children with Chronic Immune Thrombocytopenia Exhibit High Expression of Human Endogenous Retroviruses TRIM28 and SETDB1. Genes 2023, 14, 1569. https://doi.org/10.3390/genes14081569
Tovo P-A, Galliano I, Parodi E, Calvi C, Gambarino S, Licciardi F, Dini M, Montanari P, Branca M, Ramenghi U, et al. Children with Chronic Immune Thrombocytopenia Exhibit High Expression of Human Endogenous Retroviruses TRIM28 and SETDB1. Genes. 2023; 14(8):1569. https://doi.org/10.3390/genes14081569
Chicago/Turabian StyleTovo, Pier-Angelo, Ilaria Galliano, Emilia Parodi, Cristina Calvi, Stefano Gambarino, Francesco Licciardi, Maddalena Dini, Paola Montanari, Margherita Branca, Ugo Ramenghi, and et al. 2023. "Children with Chronic Immune Thrombocytopenia Exhibit High Expression of Human Endogenous Retroviruses TRIM28 and SETDB1" Genes 14, no. 8: 1569. https://doi.org/10.3390/genes14081569