
Characteristics of microRNAs in Skeletal Muscle of Intrauterine Growth-Restricted Pigs
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Sample Collection
2.3. Small RNA Sequencing
2.4. Prediction and Functional Annotation of Target Genes
2.5. Cell Culture and Transfection
2.6. Reverse Transcription Quantitative PCR (RT-qPCR)
2.7. Statistical Analysis
3. Results
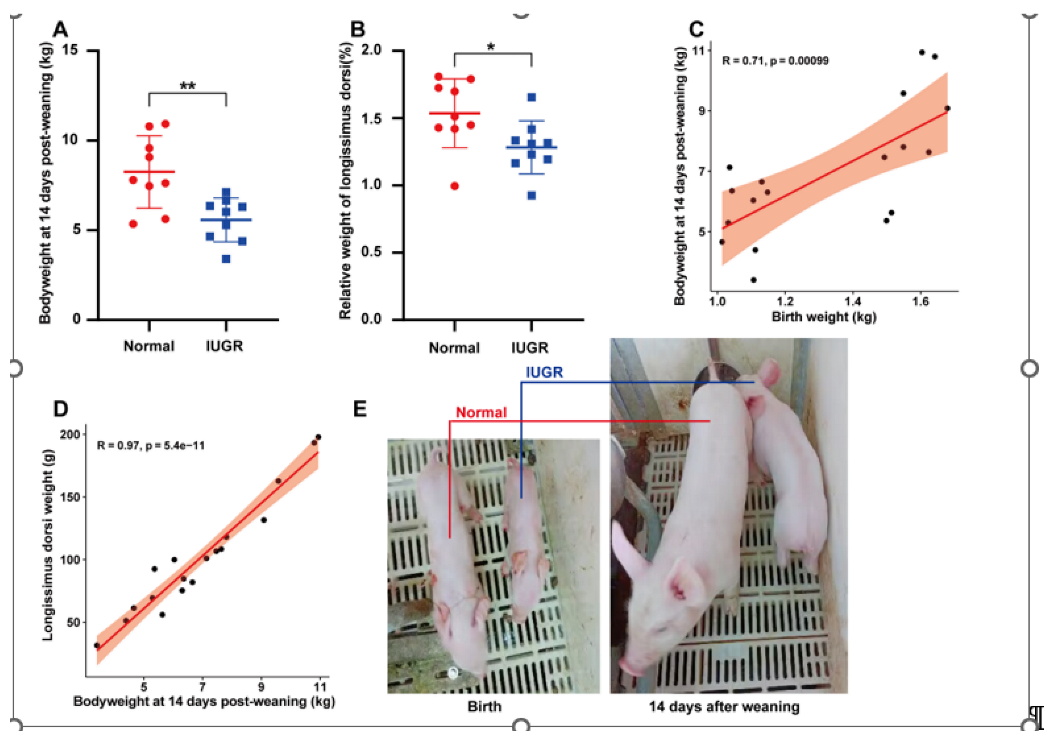
3.1. Phenotypic Characterization of Pigs with Intrauterine Growth Retardation
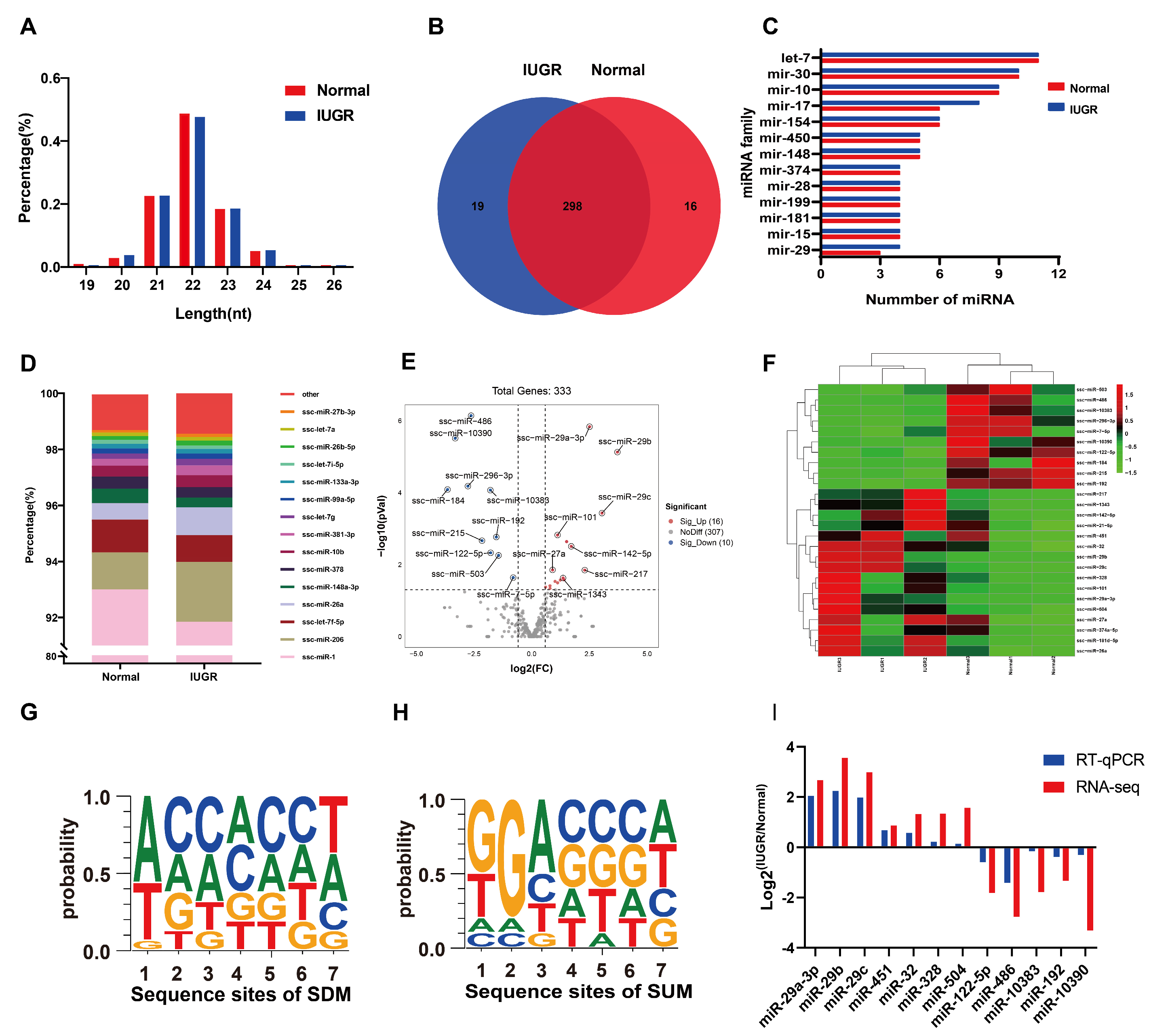
3.2. The Expression Characteristics of miRNA in IUGR Pig Skeletal Muscle
3.3. Differentially Expressed miRNAs between IUGR and Normal Pig Skeletal Muscle
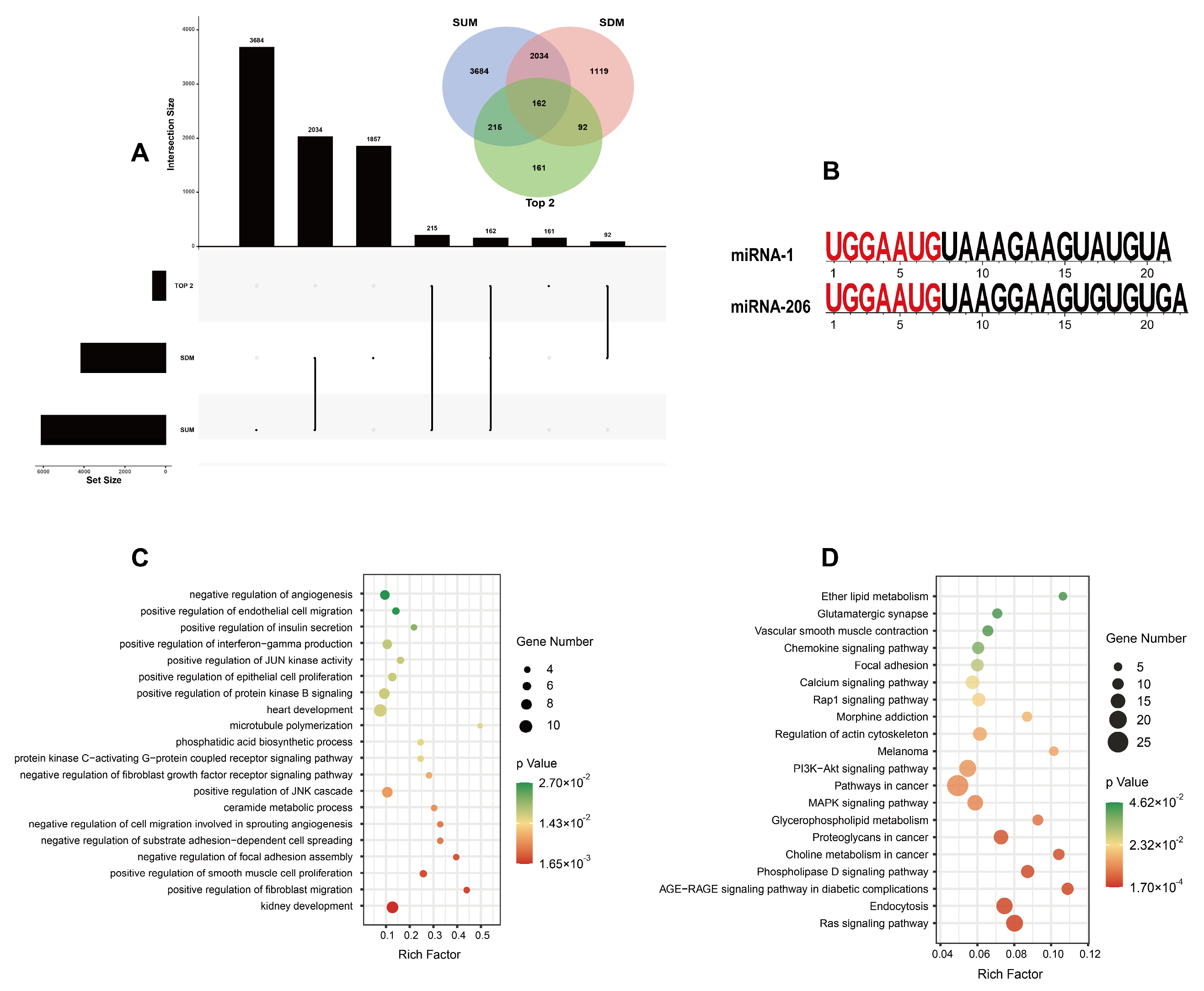
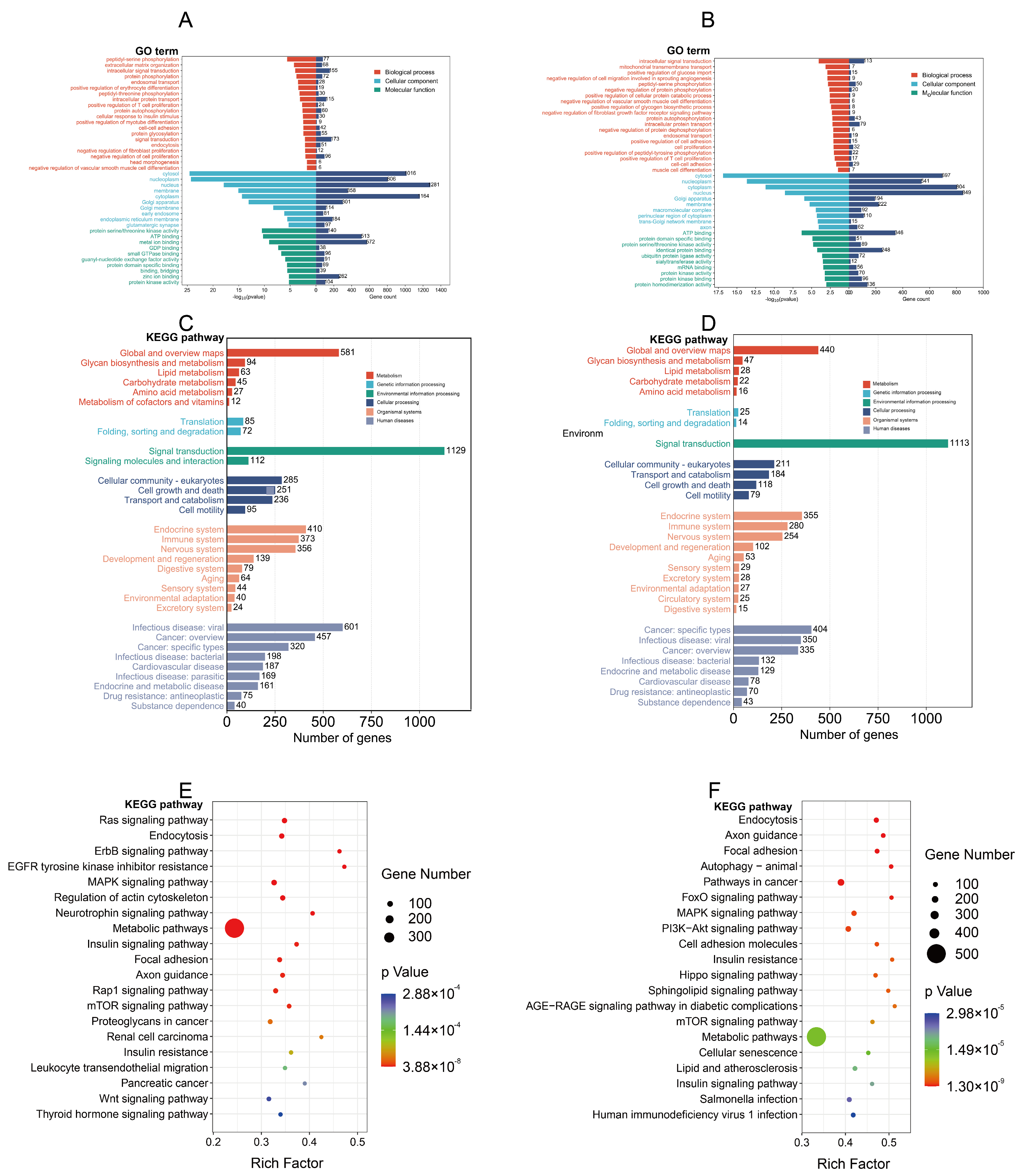
3.4. Target Gene Prediction and Functional Analysis of Differentially Expressed miRNAs
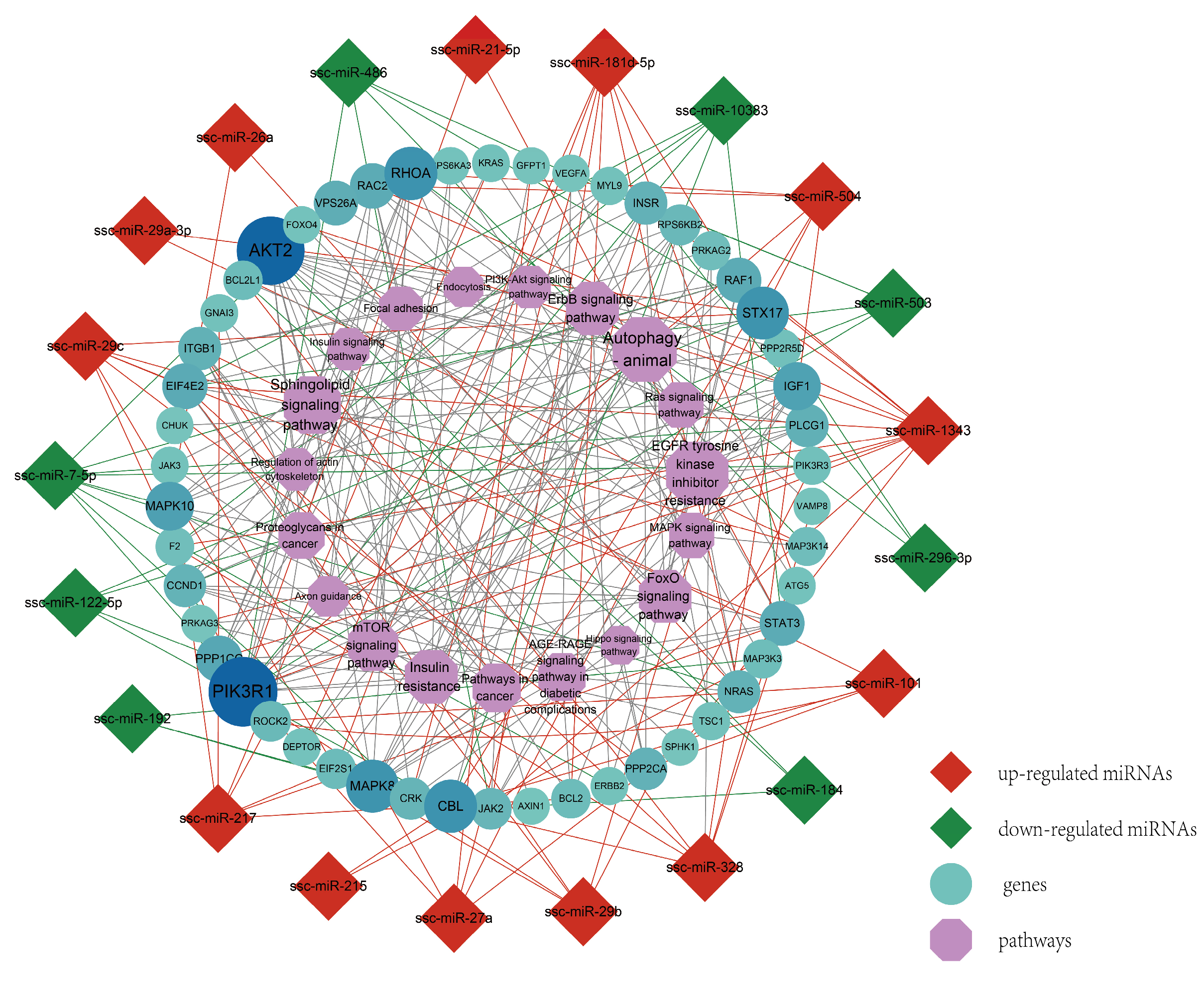
3.5. Protein–Protein Interaction Network Construction of Target Genes
3.6. Regulation Network and Correlation Analysis of Muscle Development-Related Pathways
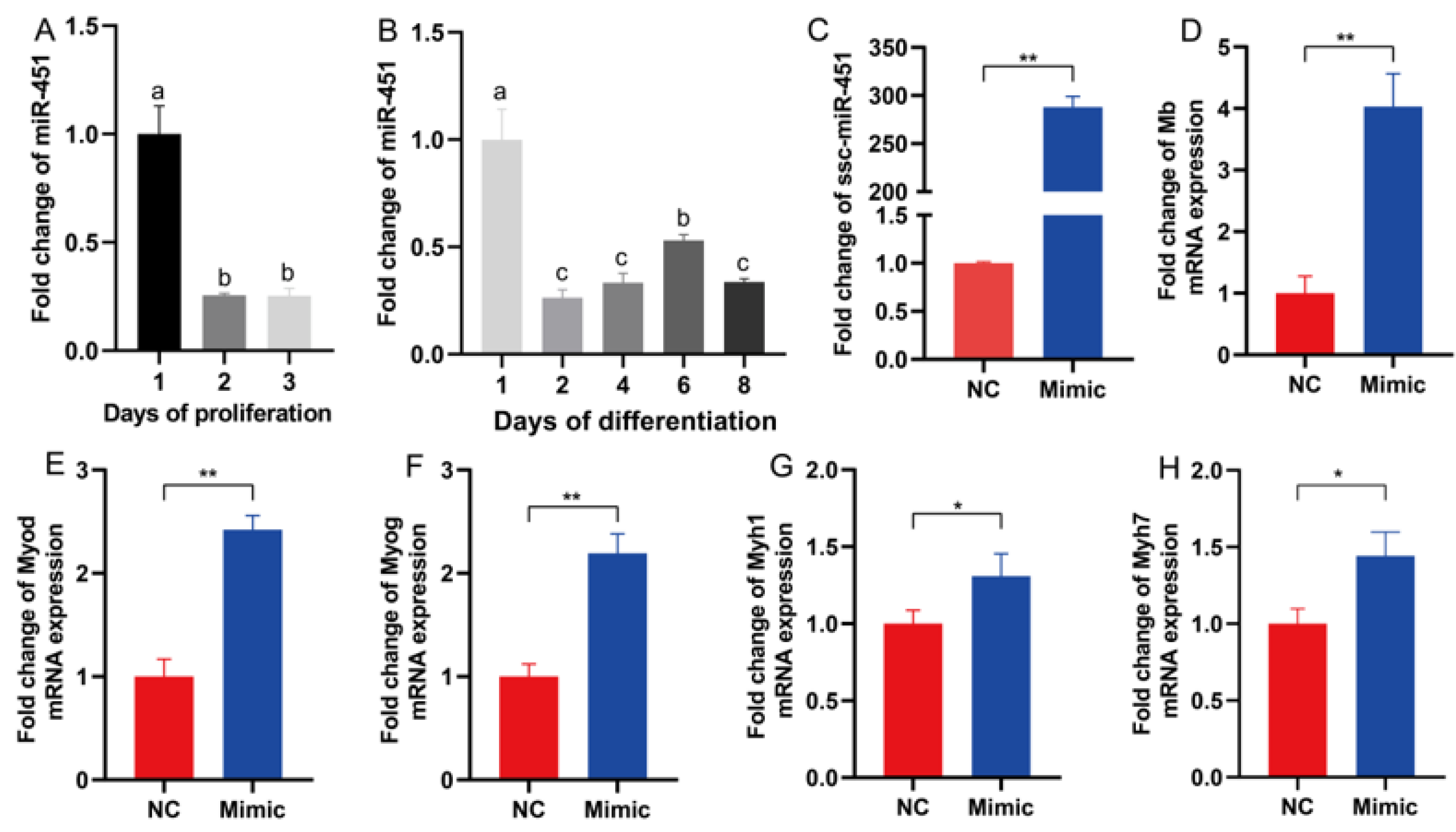
3.7. miR-451 Is Involved in the Regulation of Skeletal Muscle Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharma, D.; Shastri, S.; Sharma, P. Intrauterine Growth Restriction: Antenatal and Postnatal Aspects. Clin. Med. Insights Pediatr. 2016, 10, 67–83. [Google Scholar] [CrossRef]
- Unterscheider, J.; Daly, S.; Geary, M.P.; Kennelly, M.M.; McAuliffe, F.M.; O’Donoghue, K.; Hunter, A.; Morrison, J.J.; Burke, G.; Dicker, P.; et al. Optimizing the definition of intrauterine growth restriction: The multicenter prospective PORTO Study. Am. J. Obstet. Gynecol. 2013, 208, 290.e291–296. [Google Scholar] [CrossRef]
- Majewska, M.; Lipka, A.; Paukszto, L.; Jastrzebski, J.P.; Szeszko, K.; Gowkielewicz, M.; Lepiarczyk, E.; Jozwik, M.; Majewski, M.K. Placenta Transcriptome Profiling in Intrauterine Growth Restriction (IUGR). Int. J. Mol. Sci. 2019, 20, 1510. [Google Scholar] [CrossRef] [PubMed]
- Longo, S.; Borghesi, A.; Tzialla, C.; Stronati, M. IUGR and infections. Early Hum. Dev. 2014, 90 (Suppl. S1), S42–S44. [Google Scholar] [CrossRef] [PubMed]
- Chatmethakul, T.; Roghair, R.D. Risk of hypertension following perinatal adversity: IUGR and prematurity. J. Endocrinol. 2019, 242, T21–T32. [Google Scholar] [CrossRef]
- Nowakowska, B.A.; Pankiewicz, K.; Nowacka, U.; Niemiec, M.; Kozłowski, S.; Issat, T. Genetic Background of Fetal Growth Restriction. Int. J. Mol. Sci. 2021, 23, 36. [Google Scholar] [CrossRef]
- Zur, R.L.; Kingdom, J.C.; Parks, W.T.; Hobson, S.R. The Placental Basis of Fetal Growth Restriction. Obstet. Gynecol. Clin. N. Am. 2020, 47, 81–98. [Google Scholar] [CrossRef]
- De Onis, M.; Blössner, M.; Villar, J. Levels and patterns of intrauterine growth retardation in developing countries. Eur. J. Clin. Nutr. 1998, 52 (Suppl. S1), S5–S15. [Google Scholar]
- Mierzynski, R.; Dluski, D.; Darmochwal-Kolarz, D.; Poniedziałek-Czajkowska, E.; Leszczynska-Gorzelak, B.; Kimber-Trojnar, Z.; Agnieszka, W.; Oleszczuk, J. Intra-uterine Growth Retardation as a Risk Factor of Postnatal Metabolic Disorders. Curr. Pharm. Biotechnol. 2016, 17, 587–596. [Google Scholar] [CrossRef]
- Darendeliler, F. IUGR: Genetic influences, metabolic problems, environmental associations/triggers, current and future management. Best Pract. Res. Clin. Endocrinol. Metab. 2019, 33, 101260. [Google Scholar] [CrossRef]
- Limesand, S.W.; Rozance, P.J.; Smith, D.; Hay, W.W., Jr. Increased insulin sensitivity and maintenance of glucose utilization rates in fetal sheep with placental insufficiency and intrauterine growth restriction. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1716–E1725. [Google Scholar] [CrossRef]
- Cohen, E.; Wong, F.Y.; Horne, R.S.; Yiallourou, S.R. Intrauterine growth restriction: Impact on cardiovascular development and function throughout infancy. Pediatr. Res. 2016, 79, 821–830. [Google Scholar] [CrossRef]
- Sandoval, C.; Wu, G.; Smith, S.B.; Dunlap, K.A.; Satterfield, M.C. Maternal Nutrient Restriction and Skeletal Muscle Development: Consequences for Postnatal Health. Adv. Exp. Med. Biol. 2020, 1265, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Yates, D.T.; Macko, A.R.; Nearing, M.; Chen, X.; Rhoads, R.P.; Limesand, S.W. Developmental programming in response to intrauterine growth restriction impairs myoblast function and skeletal muscle metabolism. J. Pregnancy 2012, 2012, 631038. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Peng, X.; Han, F.; Wu, F.; Chen, D.; Wu, D.; Feyera, T.; Zhang, K.; Che, L. Effects of Birth Weight and Postnatal Nutritional Restriction on Skeletal Muscle Development, Myofiber Maturation, and Metabolic Status of Early-Weaned Piglets. Animals 2020, 10, 156. [Google Scholar] [CrossRef]
- Longo, S.; Bollani, L.; Decembrino, L.; Di Comite, A.; Angelini, M.; Stronati, M. Short-term and long-term sequelae in intrauterine growth retardation (IUGR). J. Matern. Fetal Neonatal Med. 2013, 26, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Gatford, K.L.; Simmons, R.A. Prenatal programming of insulin secretion in intrauterine growth restriction. Clin. Obstet. Gynecol. 2013, 56, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Febbraio, M.A. Muscles, exercise and obesity: Skeletal muscle as a secretory organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef]
- Mukund, K.; Subramaniam, S. Skeletal muscle: A review of molecular structure and function, in health and disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1462. [Google Scholar] [CrossRef]
- Năstase, L.; Cretoiu, D.; Stoicescu, S.M. Skeletal Muscle Damage in Intrauterine Growth Restriction. Adv. Exp. Med. Biol. 2018, 1088, 93–106. [Google Scholar] [CrossRef]
- Chal, J.; Pourquié, O. Making muscle: Skeletal myogenesis in vivo and in vitro. Development 2017, 144, 2104–2122. [Google Scholar] [CrossRef]
- Brown, L.D.; Hay, W.W., Jr. Impact of placental insufficiency on fetal skeletal muscle growth. Mol. Cell. Endocrinol. 2016, 435, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Felicioni, F.; Pereira, A.D.; Caldeira-Brant, A.L.; Santos, T.G.; Paula, T.M.D.; Magnabosco, D.; Bortolozzo, F.P.; Tsoi, S.; Dyck, M.K.; Dixon, W.; et al. Postnatal development of skeletal muscle in pigs with intrauterine growth restriction: Morphofunctional phenotype and molecular mechanisms. J. Anat. 2020, 236, 840–853. [Google Scholar] [CrossRef] [PubMed]
- Rashid, C.S.; Bansal, A.; Simmons, R.A. Oxidative Stress, Intrauterine Growth Restriction, and Developmental Programming of Type 2 Diabetes. Physiology 2018, 33, 348–359. [Google Scholar] [CrossRef]
- De Blasio, M.J.; Gatford, K.L.; Harland, M.L.; Robinson, J.S.; Owens, J.A. Placental restriction reduces insulin sensitivity and expression of insulin signaling and glucose transporter genes in skeletal muscle, but not liver, in young sheep. Endocrinology 2012, 153, 2142–2151. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, H.; Yang, L.; Wang, X.; Yang, L.; Xing, Y.; Lv, X.; Ma, H.; Song, G. The role of CD36-Fabp4-PPARγ in skeletal muscle involves insulin resistance in intrauterine growth retardation mice with catch-up growth. BMC Endocr. Disord. 2022, 22, 10. [Google Scholar] [CrossRef]
- Morrison, J.L.; Duffield, J.A.; Muhlhausler, B.S.; Gentili, S.; McMillen, I.C. Fetal growth restriction, catch-up growth and the early origins of insulin resistance and visceral obesity. Pediatr. Nephrol. 2010, 25, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Thamotharan, M.; Shin, B.C.; Suddirikku, D.T.; Thamotharan, S.; Garg, M.; Devaskar, S.U. GLUT4 expression and subcellular localization in the intrauterine growth-restricted adult rat female offspring. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E935–E947. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Bushati, N.; Cohen, S.M. microRNA functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 175–205. [Google Scholar] [CrossRef]
- Ho, P.T.B.; Clark, I.M.; Le, L.T.T. MicroRNA-Based Diagnosis and Therapy. Int. J. Mol. Sci. 2022, 23, 7167. [Google Scholar] [CrossRef]
- Zhu, Y.; Ma, J.; Pan, H.; Gan, M.; Shen, L. MiR-29a Family as a Key Regulator of Skeletal Muscle Dysplasia in a Porcine Model of Intrauterine Growth Retardation. Biomolecules 2022, 12, 1193. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Kang, X.; Shan, Y.; Zhang, M.; Gao, Y.; Wu, W.; Chen, L. miR-1227-3p participates in the development of fetal growth restriction via regulating trophoblast cell proliferation and apoptosis. Sci. Rep. 2022, 12, 6374. [Google Scholar] [CrossRef] [PubMed]
- Saget, S.; Cong, R.; Decourtye, L.; Endale, M.L.; Martinerie, L.; Girardet, C.; Perret, C.; Clemessy, M.; Leneuve, P.; Dinard, L.; et al. Changes in circulating miRNA19a-3p precede insulin resistance programmed by intra-uterine growth retardation in mice. Mol. Metab. 2020, 42, 101083. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Van Goor, A.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the pig as a human biomedical model. Sci. Transl. Med. 2021, 13, eabd5758. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, L.; Wang, L.; Liu, X.; Hou, X.; Zhao, F.; Yan, H.; Wang, L. Liver transcriptome profiling and functional analysis of intrauterine growth restriction (IUGR) piglets reveals a genetic correction and sexual-dimorphic gene expression during postnatal development. BMC Genom. 2020, 21, 701. [Google Scholar] [CrossRef]
- Gan, M.; Ma, J.; Chen, L.; Zhang, S.; Niu, L.; Zhao, Y.; Li, X.; Pan, H.; Zhu, L.; Shen, L. Identification of tRNA-derived small RNAs and their potential roles in porcine skeletal muscle with intrauterine growth restriction. Front. Physiol. 2022, 13, 962278. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Pathophysiology of placental-derived fetal growth restriction. Am. J. Obstet. Gynecol. 2018, 218, S745–S761. [Google Scholar] [CrossRef]
- Dunlop, K.; Cedrone, M.; Staples, J.F.; Regnault, T.R. Altered fetal skeletal muscle nutrient metabolism following an adverse in utero environment and the modulation of later life insulin sensitivity. Nutrients 2015, 7, 1202–1216. [Google Scholar] [CrossRef]
- Luo, J.; Fan, Y.; Shen, L.; Niu, L.; Zhao, Y.; Jiang, D.; Zhu, L.; Jiang, A.; Tang, Q.; Ma, J.; et al. The Pro-angiogenesis Of Exosomes Derived From Umbilical Cord Blood Of Intrauterine Growth Restriction Pigs Was Repressed Associated with MiRNAs. Int. J. Biol. Sci. 2018, 14, 1426–1436. [Google Scholar] [CrossRef]
- Ma, J.; Gan, M.; Chen, J.; Chen, L.; Zhao, Y.; Zhu, Y.; Niu, L.; Zhang, S.; Jiang, Y.; Guo, Z.; et al. Characteristics of tRNA-Derived Small RNAs and microRNAs Associated with Immunocompromise in an Intrauterine Growth-Restricted Pig Model. Animals 2022, 12, 2102. [Google Scholar] [CrossRef]
- Tan, Y.; Shen, L.; Gan, M.; Fan, Y.; Cheng, X.; Zheng, T.; Niu, L.; Chen, L.; Jiang, D.; Li, X.; et al. Downregulated miR-204 Promotes Skeletal Muscle Regeneration. Biomed. Res. Int. 2020, 2020, 3183296. [Google Scholar] [CrossRef] [PubMed]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA translation and stability by microRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef]
- Khordadmehr, M.; Jigari-Asl, F.; Ezzati, H.; Shahbazi, R.; Sadreddini, S.; Safaei, S.; Baradaran, B. A comprehensive review on miR-451: A promising cancer biomarker with therapeutic potential. J. Cell. Physiol. 2019, 234, 21716–21731. [Google Scholar] [CrossRef]
- Pan, X.; Wang, R.; Wang, Z.X. The potential role of miR-451 in cancer diagnosis, prognosis, and therapy. Mol. Cancer Ther. 2013, 12, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef]
- Sayed, D.; Abdellatif, M. MicroRNAs in development and disease. Physiol. Rev. 2011, 91, 827–887. [Google Scholar] [CrossRef] [PubMed]
- Devaskar, S.U.; Chu, A. Intrauterine Growth Restriction: Hungry for an Answer. Physiology 2016, 31, 131–146. [Google Scholar] [CrossRef]
- Garite, T.J.; Clark, R.; Thorp, J.A. Intrauterine growth restriction increases morbidity and mortality among premature neonates. Am. J. Obstet. Gynecol. 2004, 191, 481–487. [Google Scholar] [CrossRef]
- Wu, Z.; Nie, J.; Wu, D.; Huang, S.; Chen, J.; Liang, H.; Hao, X.; Feng, L.; Luo, H.; Tan, C. Dietary adenosine supplementation improves placental angiogenesis in IUGR piglets by up-regulating adenosine A2a receptor. Anim. Nutr. 2023, 13, 282–288. [Google Scholar] [CrossRef]
- Sempere, L.F.; Freemantle, S.; Pitha-Rowe, I.; Moss, E.; Dmitrovsky, E.; Ambros, V. Expression profiling of mammalian microRNAs uncovers a subset of brain-expressed microRNAs with possible roles in murine and human neuronal differentiation. Genome Biol. 2004, 5, R13. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Lee, Y.S.; Sivaprasad, U.; Malhotra, A.; Dutta, A. Muscle-specific microRNA miR-206 promotes muscle differentiation. J. Cell. Biol. 2006, 174, 677–687. [Google Scholar] [CrossRef]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef]
- Nicolai, S.; Pieraccioli, M.; Smirnov, A.; Pitolli, C.; Anemona, L.; Mauriello, A.; Candi, E.; Annicchiarico-Petruzzelli, M.; Shi, Y.; Wang, Y.; et al. ZNF281/Zfp281 is a target of miR-1 and counteracts muscle differentiation. Mol. Oncol. 2020, 14, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Galimov, A.; Merry, T.L.; Luca, E.; Rushing, E.J.; Mizbani, A.; Turcekova, K.; Hartung, A.; Croce, C.M.; Ristow, M.; Krützfeldt, J. MicroRNA-29a in Adult Muscle Stem Cells Controls Skeletal Muscle Regeneration During Injury and Exercise Downstream of Fibroblast Growth Factor-2. Stem Cells 2016, 34, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Du, H.; Wei, S.; Feng, L.; Li, J.; Yao, F.; Zhang, M.; Hatch, G.M.; Chen, L. Adipocyte-Derived Exosomal MiR-27a Induces Insulin Resistance in Skeletal Muscle Through Repression of PPARγ. Theranostics 2018, 8, 2171–2188. [Google Scholar] [CrossRef]
- Chen, X.; Huang, Z.; Chen, D.; Yang, T.; Liu, G. MicroRNA-27a is induced by leucine and contributes to leucine-induced proliferation promotion in C2C12 cells. Int. J. Mol. Sci. 2013, 14, 14076–14084. [Google Scholar] [CrossRef]
- Samani, A.; Hightower, R.M.; Reid, A.L.; English, K.G.; Lopez, M.A.; Doyle, J.S.; Conklin, M.J.; Schneider, D.A.; Bamman, M.M.; Widrick, J.J.; et al. miR-486 is essential for muscle function and suppresses a dystrophic transcriptome. Life Sci. Alliance 2022, 5, e202101215. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, S.; Kang, Z.; Ru, W.; Shen, X.; Li, M.; Lan, X.; Chen, H. circMEF2D Negatively Regulated by HNRNPA1 Inhibits Proliferation and Differentiation of Myoblasts via miR-486-PI3K/AKT Axis. J. Agric. Food Chem. 2022, 70, 8145–8163. [Google Scholar] [CrossRef]
- Dey, B.K.; Gagan, J.; Yan, Z.; Dutta, A. miR-26a is required for skeletal muscle differentiation and regeneration in mice. Genes Dev. 2012, 26, 2180–2191. [Google Scholar] [CrossRef]
- Wu, J.; He, D.; Yue, B.; Zhang, C.; Fang, X.; Chen, H. miR-101-1 expression pattern in Qinchuan cattle and its role in the regulation of cell differentiation. Gene 2017, 636, 64–69. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, H.; Li, Y.; Liu, S.; Chen, J.; Ying, H. miR-24 and miR-122 Negatively Regulate the Transforming Growth Factor-β/Smad Signaling Pathway in Skeletal Muscle Fibrosis. Mol. Ther. Nucleic Acids 2018, 11, 528–537. [Google Scholar] [CrossRef]
- Wang, F.; Liang, R.; Tandon, N.; Matthews, E.R.; Shrestha, S.; Yang, J.; Soibam, B.; Yang, J.; Liu, Y. H19X-encoded miR-424(322)/-503 cluster: Emerging roles in cell differentiation, proliferation, plasticity and metabolism. Cell. Mol. Life Sci. 2019, 76, 903–920. [Google Scholar] [CrossRef]
- Sarkar, S.; Dey, B.K.; Dutta, A. MiR-322/424 and -503 are induced during muscle differentiation and promote cell cycle quiescence and differentiation by down-regulation of Cdc25A. Mol. Biol. Cell 2010, 21, 2138–2149. [Google Scholar] [CrossRef]
- Li, Z.; Abdalla, B.A.; Zheng, M.; He, X.; Cai, B.; Han, P.; Ouyang, H.; Chen, B.; Nie, Q.; Zhang, X. Systematic transcriptome-wide analysis of mRNA-miRNA interactions reveals the involvement of miR-142-5p and its target (FOXO3) in skeletal muscle growth in chickens. Mol. Genet. Genom. 2018, 293, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Murani, E.; Hadlich, F.; Liu, X.; Wimmers, K.; Ponsuksili, S. In Utero Fetal Weight in Pigs Is Regulated by microRNAs and Their Target Genes. Genes 2021, 12, 1264. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Huang, X.; Liu, C.; Shi, M.; Li, W.; Niu, J.; Cai, C.; Yang, Y.; Gao, P.; Guo, X.; et al. Genome-wide analysis of circular RNA-mediated ceRNA regulation in porcine skeletal muscle development. BMC Genom. 2023, 24, 196. [Google Scholar] [CrossRef]
- Liu, S.; Xie, S.; Chen, H.; Li, B.; Chen, Z.; Tan, Y.; Yang, J.; Zheng, L.; Xiao, Z.; Zhang, Q.; et al. The functional analysis of transiently upregulated miR-101 suggests a “braking” regulatory mechanism during myogenesis. Sci. China Life Sci. 2021, 64, 1612–1623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ran, J.; Li, J.; Yu, C.; Cui, Z.; Amevor, F.K.; Wang, Y.; Jiang, X.; Qiu, M.; Du, H.; et al. miR-21-5p Regulates the Proliferation and Differentiation of Skeletal Muscle Satellite Cells by Targeting KLF3 in Chicken. Genes 2021, 12, 814. [Google Scholar] [CrossRef]
- Zhang, H.; Chi, M.; Chen, L.; Sun, X.; Wan, L.; Yang, Q.; Guo, C. Daidzein alleviates cisplatin-induced muscle atrophy by regulating Glut4/AMPK/FoxO pathway. Phytother. Res. 2021, 35, 4363–4376. [Google Scholar] [CrossRef] [PubMed]
- Thomson, D.M. The Role of AMPK in the Regulation of Skeletal Muscle Size, Hypertrophy, and Regeneration. Int. J. Mol. Sci. 2018, 19, 3125. [Google Scholar] [CrossRef]
- Lee, S.; Dong, H.H. FoxO integration of insulin signaling with glucose and lipid metabolism. J. Endocrinol. 2017, 233, R67–R79. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Tanaka, H.; Tachibana, R.; Yoshikawa, K.; Kawamura, T.; Takakura, S.; Takeuchi, H.; Ikeda, T. Tadalafil Treatment of Mice with Fetal Growth Restriction and Preeclampsia Improves Placental mTOR Signaling. Int. J. Mol. Sci. 2022, 23, 1474. [Google Scholar] [CrossRef]
- Luo, Z.; Luo, W.; Li, S.; Zhao, S.; Sho, T.; Xu, X.; Zhang, J.; Xu, W.; Xu, J. Reactive oxygen species mediated placental oxidative stress, mitochondrial content, and cell cycle progression through mitogen-activated protein kinases in intrauterine growth restricted pigs. Reprod. Biol. 2018, 18, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Thorn, S.R.; Brown, L.D.; Rozance, P.J.; Hay, W.W., Jr.; Friedman, J.E. Increased hepatic glucose production in fetal sheep with intrauterine growth restriction is not suppressed by insulin. Diabetes 2013, 62, 65–73. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Q.; Wang, B.; Wu, W.; Wei, J.; Li, P.; Huang, R. miR-22 regulates C2C12 myoblast proliferation and differentiation by targeting TGFBR1. Eur. J. Cell Biol. 2018, 97, 257–268. [Google Scholar] [CrossRef]
- Wu, J.; Yue, B.; Lan, X.; Wang, Y.; Fang, X.; Ma, Y.; Bai, Y.; Qi, X.; Zhang, C.; Chen, H. MiR-499 regulates myoblast proliferation and differentiation by targeting transforming growth factor β receptor 1. J. Cell. Physiol. 2019, 234, 2523–2536. [Google Scholar] [CrossRef] [PubMed]
- Gan, M.; Shen, L.; Fan, Y.; Tan, Y.; Liu, L.; Chen, L.; Zhao, Y.; Niu, L.; Tang, G.; Li, Q.; et al. ssc-miR-451 Regulates Porcine Primary Adipocyte Differentiation by Targeting ACACA. Animals 2020, 10, 1891. [Google Scholar] [CrossRef]
- Che, J.; Xu, C.; Wu, Y.; Jia, P.; Han, Q.; Ma, Y.; Wang, X.; Zheng, Y. MiR-1290 promotes myoblast differentiation and protects against myotube atrophy via Akt/p70/FoxO3 pathway regulation. Skelet. Muscle 2021, 11, 6. [Google Scholar] [CrossRef]
- Wei, P.; Xie, Y.; Abel, P.W.; Huang, Y.; Ma, Q.; Li, L.; Hao, J.; Wolff, D.W.; Wei, T.; Tu, Y. Transforming growth factor (TGF)-β1-induced miR-133a inhibits myofibroblast differentiation and pulmonary fibrosis. Cell Death Dis. 2019, 10, 670. [Google Scholar] [CrossRef] [PubMed]
- Bruchova-Votavova, H.; Yoon, D.; Prchal, J.T. miR-451 enhances erythroid differentiation in K562 cells. Leuk. Lymphoma 2010, 51, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Qiang, T.T.; Wang, K.Y.; Wang, X.L. Shexiang Baoxin Pill Regulates Intimal Hyperplasia, Migration, and Apoptosis after Platelet-Derived Growth Factor-BB-Stimulation of Vascular Smooth Muscle Cells via miR-451. Chin. J. Integr. Med. 2022, 28, 785–793. [Google Scholar] [CrossRef]
- Song, L.; Su, M.; Wang, S.; Zou, Y.; Wang, X.; Wang, Y.; Cui, H.; Zhao, P.; Hui, R.; Wang, J. MiR-451 is decreased in hypertrophic cardiomyopathy and regulates autophagy by targeting TSC1. J. Cell. Mol. Med. 2014, 18, 2266–2274. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | miRNA-ID | Mature_Seq | Normal-CMP | IUGR-CMP | Fold Change | log2FC | p-Value |
|---|---|---|---|---|---|---|---|
| Downregulated | ssc-miR-184 | UGGACGGAGAACUGAUAAGGGU | 13.777 | 1.224 | 0.089 | −3.493 | 0.000082 |
| ssc-miR-10390 | AUACUACUGACAGACCGCAACCU | 4.344 | 0.440 | 0.101 | −3.304 | 0.000003 | |
| ssc-miR-296-3p | AGGGUUGGGCGGAGGCUUUCC | 2.181 | 0.326 | 0.150 | −2.742 | 0.000066 | |
| ssc-miR-486 | UCCUGUACUGAGCUGCCCCGAG | 69.034 | 10.174 | 0.147 | −2.762 | 0.000001 | |
| ssc-miR-215 | AUGACCUAUGAAUUGACAGAC | 4.107 | 1.039 | 0.253 | −1.983 | 0.002172 | |
| ssc-miR-10383 | UGGUGCCUGACGUCUUGGCAGU | 12.415 | 3.625 | 0.292 | −1.776 | 0.000086 | |
| ssc-miR-122-5p | UGGAGUGUGACAAUGGUGUUUGU | 16.578 | 4.712 | 0.284 | −1.815 | 0.004718 | |
| ssc-miR-192 | CUGACCUAUGAAUUGACAGCC | 14.586 | 5.810 | 0.398 | −1.328 | 0.001716 | |
| ssc-miR-503 | UAGCAGCGGGAACAGUACUGCAG | 10.981 | 4.610 | 0.420 | −1.252 | 0.005568 | |
| ssc-miR-7-5p | UGGAAGACUAGUGAUUUUGUUGUU | 51.110 | 31.545 | 0.617 | −0.696 | 0.023011 | |
| Upregulated | ssc-miR-29b | UAGCACCAUUUGAAAUCAGUGUU | 0.578 | 6.817 | 11.791 | 3.560 | 0.000007 |
| ssc-miR-29c | UAGCACCAUUUGAAAUCGGUUA | 0.289 | 2.283 | 7.905 | 2.983 | 0.000371 | |
| ssc-miR-29a-3p | CUAGCACCAUCUGAAAUCGGUUA | 11.353 | 72.181 | 6.358 | 2.669 | 0.000001 | |
| ssc-miR-217 | UACUGCAUCAGGAACUGAUUGGAU | 0.143 | 0.784 | 5.495 | 2.458 | 0.014239 | |
| ssc-miR-142-5p | CAUAAAGUAGAAAGCACUACU | 6.530 | 19.209 | 2.942 | 1.557 | 0.003117 | |
| ssc-miR-504 | AGACCCUGGUCUGCACUCUAUCU | 3.937 | 11.668 | 2.964 | 1.567 | 0.002305 | |
| ssc-miR-1343 | CUCCUGGGGCCCGCACUCUCGC | 0.768 | 2.116 | 2.756 | 1.463 | 0.026311 | |
| ssc-miR-32 | UAUUGCACAUUACUAAGUUGC | 0.955 | 2.391 | 2.504 | 1.324 | 0.023649 | |
| ssc-miR-328 | CUGGCCCUCUCUGCCCUUCCGU | 2.017 | 5.117 | 2.538 | 1.344 | 0.026897 | |
| ssc-miR-451 | AAACCGUUACCAUUACUGAGUU | 55.217 | 100.369 | 1.818 | 0.862 | 0.032048 | |
| ssc-miR-101 | UACAGUACUGUGAUAACUGAA | 369.650 | 896.228 | 2.425 | 1.278 | 0.001511 | |
| ssc-miR-181d-5p | AACAUUCAUUGUUGUCGGUGGGUU | 2.722 | 5.932 | 2.179 | 1.124 | 0.029318 | |
| ssc-miR-27a | UUCACAGUGGCUAAGUUCCGC | 49.659 | 91.698 | 1.847 | 0.885 | 0.014055 | |
| ssc-miR-374a-5p | UUAUAAUACAACCUGAUAAGUG | 18.821 | 33.522 | 1.781 | 0.833 | 0.046061 | |
| ssc-miR-21-5p | UAGCUUAUCAGACUGAUGUUGA | 641.470 | 1073.217 | 1.673 | 0.742 | 0.038817 | |
| ssc-miR-26a | UUCAAGUAAUCCAGGAUAGGCU | 5960.309 | 9830.052 | 1.649 | 0.722 | 0.042270 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, Y.; Gan, M.; Xie, Z.; Ma, J.; Chen, L.; Zhang, S.; Zhao, Y.; Niu, L.; Wang, Y.; Zhu, L.; et al. Characteristics of microRNAs in Skeletal Muscle of Intrauterine Growth-Restricted Pigs. Genes 2023, 14, 1372. https://doi.org/10.3390/genes14071372
Jing Y, Gan M, Xie Z, Ma J, Chen L, Zhang S, Zhao Y, Niu L, Wang Y, Zhu L, et al. Characteristics of microRNAs in Skeletal Muscle of Intrauterine Growth-Restricted Pigs. Genes. 2023; 14(7):1372. https://doi.org/10.3390/genes14071372
Chicago/Turabian StyleJing, Yunhong, Mailin Gan, Zhongwei Xie, Jianfeng Ma, Lei Chen, Shunhua Zhang, Ye Zhao, Lili Niu, Yan Wang, Li Zhu, and et al. 2023. "Characteristics of microRNAs in Skeletal Muscle of Intrauterine Growth-Restricted Pigs" Genes 14, no. 7: 1372. https://doi.org/10.3390/genes14071372