
Divergent Transcriptomic Effects of Allopregnanolone in Postpartum Depression
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
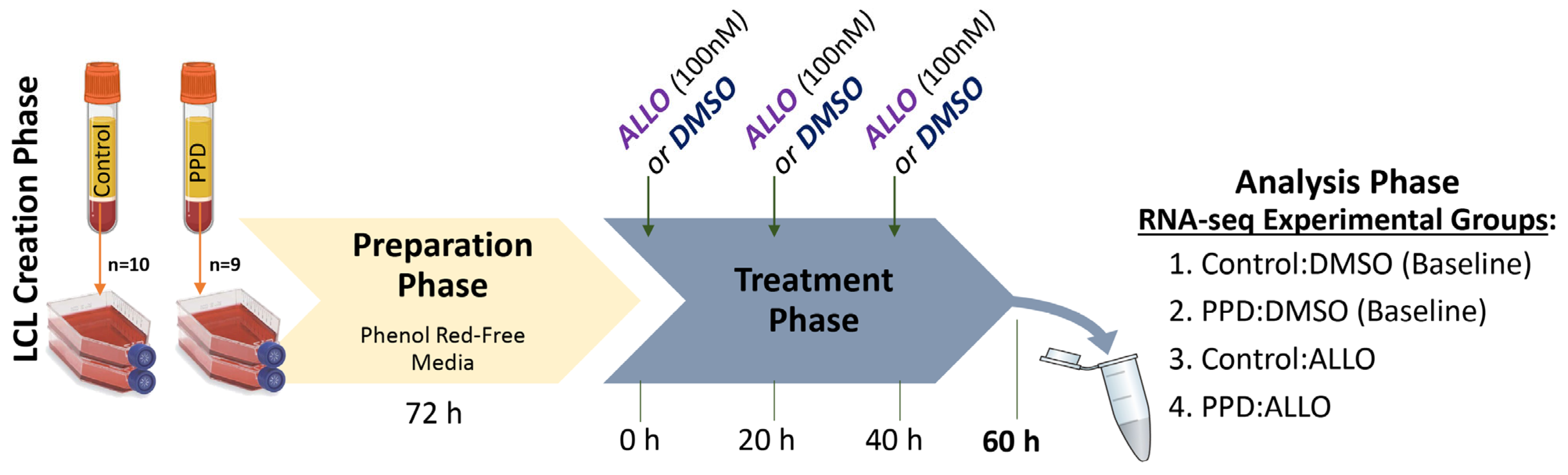
2.1. Lymphoblastoid Cell Lines
2.2. Experimental Treatment Paradigm
2.3. AmpliSeq-RNA Transcriptomics
2.4. RNA-Seq Data Analysis with TAC
2.5. qRT-PCR
2.6. Gene Network and Ontology Analyses
3. Results
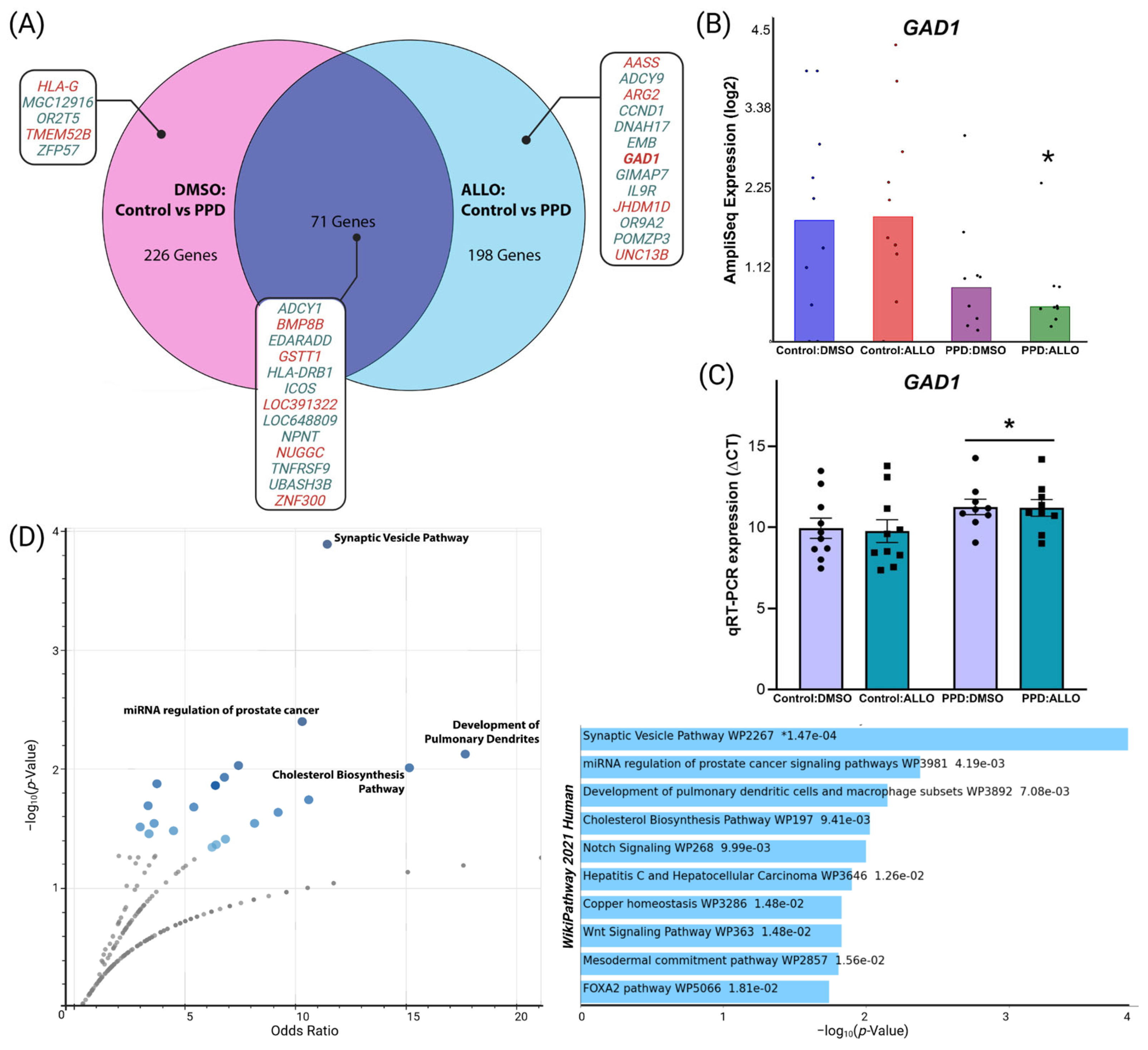
3.1. ALLO Alters Overall Patterns of Differential Gene Expression in PPD LCLs
3.2. Differential GAD1 and Synaptic-Related Gene Network Expression in PPD LCLs
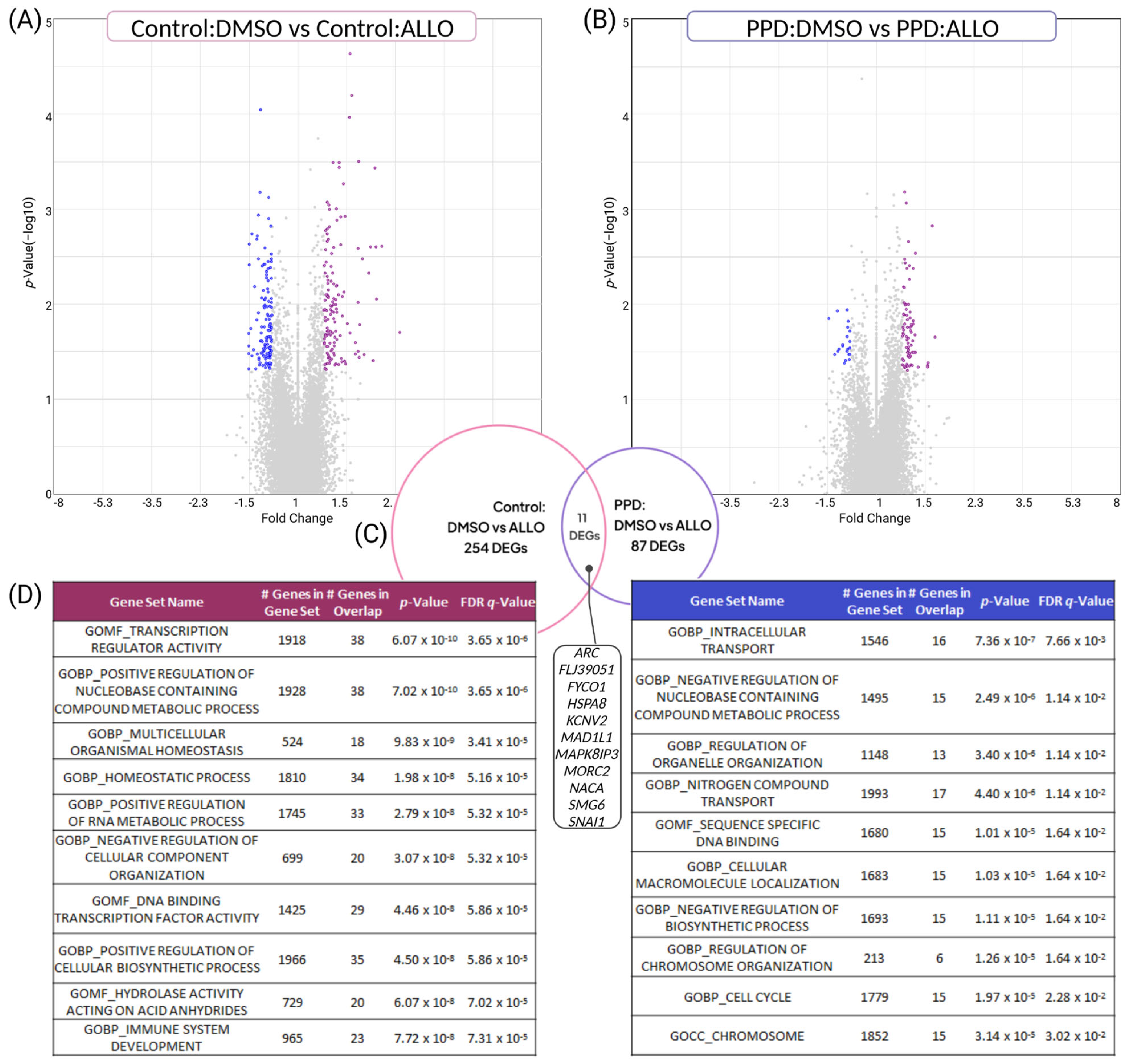
3.3. PPD LCLs Have Diminished, Divergent Transcriptomic Responses to ALLO Compared with Controls
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gavin, N.I.; Gaynes, B.N.; Lohr, K.N.; Meltzer-Brody, S.; Gartlehner, G.; Swinson, T. Perinatal depression: A systematic review of prevalence and incidence. Obstet. Gynecol. 2005, 106, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Segre, L.S.; O’Hara, M.W.; Arndt, S.; Stuart, S. The prevalence of postpartum depression: The relative significance of three social status indices. Soc. Psychiatry Psychiatr. Epidemiol. 2007, 42, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Oates, M. Perinatal psychiatric disorders: A leading cause of maternal morbidity and mortality. Br. Med. Bull. 2003, 67, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, V.; Pearson, J.L.; Colpe, L. Prevalence of suicidality during pregnancy and the postpartum. Arch. Womens Ment. Health 2005, 8, 77–87. [Google Scholar] [CrossRef]
- Munk-Olsen, T.; Ingstrup, K.G.; Johannsen, B.M.; Liu, X. Population-Based Assessment of the Recurrence Risk of Postpartum Mental Disorders: Will It Happen Again? JAMA Psychiatry 2020, 77, 213–214. [Google Scholar] [CrossRef]
- Reddy, D.S. Neurosteroids: Endogenous role in the human brain and therapeutic potentials. Prog. Brain Res. 2010, 186, 113–137. [Google Scholar] [CrossRef]
- Meltzer-Brody, S.; Colquhoun, H.; Riesenberg, R.; Epperson, C.N.; Deligiannidis, K.M.; Rubinow, D.R.; Li, H.; Sankoh, A.J.; Clemson, C.; Schacterle, A.; et al. Brexanolone injection in post-partum depression: Two multicentre, double-blind, randomised, placebo-controlled, phase 3 trials. Lancet 2018, 392, 1058–1070. [Google Scholar] [CrossRef]
- Kanes, S.; Colquhoun, H.; Gunduz-Bruce, H.; Raines, S.; Arnold, R.; Schacterle, A.; Doherty, J.; Epperson, C.N.; Deligiannidis, K.M.; Riesenberg, R.; et al. Brexanolone (SAGE-547 injection) in post-partum depression: A randomised controlled trial. Lancet 2017, 390, 480–489. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Felzenberg, E.; Robbins, A.; Pfaff, D.W.; Schwartz-Giblin, S. Infusions of diazepam and allopregnanolone into the midbrain central gray facilitate open-field behavior and sexual receptivity in female rats. Horm. Behav. 1995, 29, 279–295. [Google Scholar] [CrossRef]
- Pinna, G.; Rasmusson, A.M. Up-regulation of neurosteroid biosynthesis as a pharmacological strategy to improve behavioural deficits in a putative mouse model of post-traumatic stress disorder. J. Neuroendocrinol. 2012, 24, 102–116. [Google Scholar] [CrossRef]
- Antonoudiou, P.; Colmers, P.L.W.; Walton, N.L.; Weiss, G.L.; Smith, A.C.; Nguyen, D.P.; Lewis, M.; Quirk, M.C.; Barros, L.; Melon, L.C.; et al. Allopregnanolone Mediates Affective Switching Through Modulation of Oscillatory States in the Basolateral Amygdala. Biol. Psychiatry 2022, 91, 283–293. [Google Scholar] [CrossRef]
- Maguire, J.; Mody, I. Neurosteroid synthesis-mediated regulation of GABA(A) receptors: Relevance to the ovarian cycle and stress. J. Neurosci. 2007, 27, 2155–2162. [Google Scholar] [CrossRef] [Green Version]
- Melon, L.; Hammond, R.; Lewis, M.; Maguire, J. A Novel, Synthetic, Neuroactive Steroid Is Effective at Decreasing Depression-Like Behaviors and Improving Maternal Care in Preclinical Models of Postpartum Depression. Front. Endocrinol. 2018, 9, 703. [Google Scholar] [CrossRef] [Green Version]
- Walton, N.; Maguire, J. Allopregnanolone-based treatments for postpartum depression: Why/how do they work? Neurobiol. Stress 2019, 11, 100198. [Google Scholar] [CrossRef]
- Schiller, C.E.; Schmidt, P.J.; Rubinow, D.R. Allopregnanolone as a mediator of affective switching in reproductive mood disorders. Psychopharmacology 2014, 231, 3557–3567. [Google Scholar] [CrossRef] [Green Version]
- Gunduz-Bruce, H.; Silber, C.; Kaul, I.; Rothschild, A.J.; Riesenberg, R.; Sankoh, A.J.; Li, H.; Lasser, R.; Zorumski, C.F.; Rubinow, D.R.; et al. Trial of SAGE-217 in Patients with Major Depressive Disorder. N. Engl. J. Med. 2019, 381, 903–911. [Google Scholar] [CrossRef]
- Hernandez, G.D.; Solinsky, C.M.; Mack, W.J.; Kono, N.; Rodgers, K.E.; Wu, C.Y.; Mollo, A.R.; Lopez, C.M.; Pawluczyk, S.; Bauer, G.; et al. Safety, tolerability, and pharmacokinetics of allopregnanolone as a regenerative therapeutic for Alzheimer’s disease: A single and multiple ascending dose phase 1b/2a clinical trial. Alzheimers Dement. 2020, 6, e12107. [Google Scholar] [CrossRef]
- Schiller, C.E.; Meltzer-Brody, S.; Rubinow, D.R. The role of reproductive hormones in postpartum depression. CNS Spectr. 2015, 20, 48–59. [Google Scholar] [CrossRef]
- Rudzinskas, S.A.; Goff, A.C.; Mazzu, M.A.; Schiller, C.E.; Meltzer-Brody, S.; Rubinow, D.R.; Schmidt, P.J.; Goldman, D. Intrinsically dysregulated cellular stress signaling genes and gene networks in postpartum depression. Mol. Psychiatry 2023. Online ahead of print. [Google Scholar] [CrossRef]
- Bloch, M.; Daly, R.C.; Rubinow, D.R. Endocrine factors in the etiology of postpartum depression. Compr. Psychiatry 2003, 44, 234–246. [Google Scholar] [CrossRef]
- Bloch, M.; Schmidt, P.J.; Danaceau, M.; Murphy, J.; Nieman, L.; Rubinow, D.R. Effects of gonadal steroids in women with a history of postpartum depression. Am. J. Psychiatry 2000, 157, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Schiller, C.E.; Walsh, E.; Eisenlohr-Moul, T.A.; Prim, J.; Dichter, G.S.; Schiff, L.; Bizzell, J.; Slightom, S.L.; Richardson, E.C.; Belger, A.; et al. Effects of gonadal steroids on reward circuitry function and anhedonia in women with a history of postpartum depression. J. Affect Disord. 2022, 314, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Nappi, R. Serum allopregnanolone in women with postpartum “blues”. Obstet. Gynecol. 2001, 97, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Hellgren, C.; Åkerud, H.; Skalkidou, A.; Bäckström, T.; Sundström-Poromaa, I. Low Serum Allopregnanolone Is Associated with Symptoms of Depression in Late Pregnancy. Neuropsychobiology 2014, 69, 147–153. [Google Scholar] [CrossRef]
- Osborne, L.M.; Gispen, F.; Sanyal, A.; Yenokyan, G.; Meilman, S.; Payne, J.L. Lower allopregnanolone during pregnancy predicts postpartum depression: An exploratory study. Psychoneuroendocrinology 2017, 79, 116–121. [Google Scholar] [CrossRef] [Green Version]
- Backstrom, T.; Haage, D.; Lofgren, M.; Johansson, I.M.; Stromberg, J.; Nyberg, S.; Andreen, L.; Ossewaarde, L.; van Wingen, G.A.; Turkmen, S.; et al. Paradoxical effects of GABA-A modulators may explain sex steroid induced negative mood symptoms in some persons. Neuroscience 2011, 191, 46–54. [Google Scholar] [CrossRef]
- Timby, E.; Balgard, M.; Nyberg, S.; Spigset, O.; Andersson, A.; Porankiewicz-Asplund, J.; Purdy, R.H.; Zhu, D.; Backstrom, T.; Poromaa, I.S. Pharmacokinetic and behavioral effects of allopregnanolone in healthy women. Psychopharmacology 2006, 186, 414–424. [Google Scholar] [CrossRef] [Green Version]
- Bäckström, T.; Bixo, M.; Johansson, M.; Nyberg, S.; Ossewaarde, L.; Ragagnin, G.; Savic, I.; Strömberg, J.; Timby, E.; van Broekhoven, F.; et al. Allopregnanolone and mood disorders. Prog. Neurobiol. 2014, 113, 88–94. [Google Scholar] [CrossRef]
- Kask, K.; Backstrom, T.; Nilsson, L.G.; Sundstrom-Poromaa, I. Allopregnanolone impairs episodic memory in healthy women. Psychopharmacology 2008, 199, 161–168. [Google Scholar] [CrossRef]
- Freeman, E.W.; Weinstock, L.; Rickels, K.; Sondheimer, S.J.; Coutifaris, C. A placebo-controlled study of effects of oral progesterone on performance and mood. Br. J. Clin. Pharmacol. 1992, 33, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Sripada, R.K.; Marx, C.E.; King, A.P.; Rampton, J.C.; Ho, S.S.; Liberzon, I. Allopregnanolone elevations following pregnenolone administration are associated with enhanced activation of emotion regulation neurocircuits. Biol. Psychiatry 2013, 73, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Martinez, P.E.; Rubinow, D.R.; Nieman, L.K.; Koziol, D.E.; Morrow, A.L.; Schiller, C.E.; Cintron, D.; Thompson, K.D.; Khine, K.K.; Schmidt, P.J. 5alpha-Reductase Inhibition Prevents the Luteal Phase Increase in Plasma Allopregnanolone Levels and Mitigates Symptoms in Women with Premenstrual Dysphoric Disorder. Neuropsychopharmacology 2016, 41, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Freeman, E.W.; Frye, C.A.; Rickels, K.; Martin, P.A.G.; Smith, S.S. Allopregnanolone Levels and Symptom Improvement in Severe Premenstrual Syndrome. J. Clin. Psychopharmacol. 2002, 22, 516–520. [Google Scholar] [CrossRef]
- Andreen, L.; Nyberg, S.; Turkmen, S.; van Wingen, G.; Fernandez, G.; Backstrom, T. Sex steroid induced negative mood may be explained by the paradoxical effect mediated by GABAA modulators. Psychoneuroendocrinology 2009, 34, 1121–1132. [Google Scholar] [CrossRef]
- Dubey, N.; Hoffman, J.F.; Schuebel, K.; Yuan, Q.; Martinez, P.E.; Nieman, L.K.; Rubinow, D.R.; Schmidt, P.J.; Goldman, D. The ESC/E(Z) complex, an effector of response to ovarian steroids, manifests an intrinsic difference in cells from women with premenstrual dysphoric disorder. Mol. Psychiatry 2017, 22, 1172–1184. [Google Scholar] [CrossRef] [Green Version]
- Dionisio, L.; Jose De Rosa, M.; Bouzat, C.; Esandi Mdel, C. An intrinsic GABAergic system in human lymphocytes. Neuropharmacology 2011, 60, 513–519. [Google Scholar] [CrossRef]
- Alam, S.; Laughton, D.L.; Walding, A.; Wolstenholme, A.J. Human peripheral blood mononuclear cells express GABAA receptor subunits. Mol. Immunol. 2006, 43, 1432–1442. [Google Scholar] [CrossRef]
- Mendu, S.K.; Bhandage, A.; Jin, Z.; Birnir, B. Different subtypes of GABA-A receptors are expressed in human, mouse and rat T lymphocytes. PLoS ONE 2012, 7, e42959. [Google Scholar] [CrossRef] [Green Version]
- Kaminsky, Z.; Payne, J. Seeing the future: Epigenetic biomarkers of postpartum depression. Neuropsychopharmacology 2014, 39, 233–234. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.E.; Liu, X.; Byrne, E.M.; Sullivan, P.F.; Wray, N.R.; Agerbo, E.; Nyegaard, M.; Grove, J.; Musliner, K.L.; Ingstrup, K.G.; et al. Genetic risk scores for major psychiatric disorders and the risk of postpartum psychiatric disorders. Transl. Psychiatry 2019, 9, 288. [Google Scholar] [CrossRef] [Green Version]
- Balan, I.; Patterson, R.; Boero, G.; Krohn, H.; O’Buckley, T.K.; Meltzer-Brody, S.; Morrow, A.L. Brexanolone therapeutics in post-partum depression involves inhibition of systemic inflammatory pathways. EBioMedicine 2023, 89, 104473. [Google Scholar] [CrossRef] [PubMed]
- Balan, I.; Beattie, M.C.; O’Buckley, T.K.; Aurelian, L.; Morrow, A.L. Endogenous Neurosteroid (3alpha,5alpha)3-Hydroxypregnan-20-one Inhibits Toll-like-4 Receptor Activation and Pro-inflammatory Signaling in Macrophages and Brain. Sci. Rep. 2019, 9, 1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frye, C.A.; Koonce, C.J.; Walf, A.A. Novel receptor targets for production and action of allopregnanolone in the central nervous system: A focus on pregnane xenobiotic receptor. Front. Cell Neurosci. 2014, 8, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balan, I.; Aurelian, L.; Williams, K.S.; Campbell, B.; Meeker, R.B.; Morrow, A.L. Inhibition of human macrophage activation via pregnane neurosteroid interactions with toll-like receptors: Sex differences and structural requirements. Front. Immunol. 2022, 13, 940095. [Google Scholar] [CrossRef]
- Thomas, P.; Pang, Y. Membrane progesterone receptors: Evidence for neuroprotective, neurosteroid signaling and neuroendocrine functions in neuronal cells. Neuroendocrinology 2012, 96, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Guennoun, R. Progesterone in the Brain: Hormone, Neurosteroid and Neuroprotectant. Int. J. Mol. Sci. 2020, 21, 5271. [Google Scholar] [CrossRef]
- Berthois, Y.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S. Phenol red in tissue culture media is a weak estrogen: Implications concerning the study of estrogen-responsive cells in culture. Proc. Natl. Acad. Sci. USA 1986, 83, 2496–2500. [Google Scholar] [CrossRef] [Green Version]
- Van der Valk, J. Fetal bovine serum-a cell culture dilemma. Science 2022, 375, 143–144. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Liberzon, A.; Birger, C.; Thorvaldsdottir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Deligiannidis, K.M.; Meltzer-Brody, S.; Gunduz-Bruce, H.; Doherty, J.; Jonas, J.; Li, S.; Sankoh, A.J.; Silber, C.; Campbell, A.D.; Werneburg, B.; et al. Effect of Zuranolone vs Placebo in Postpartum Depression: A Randomized Clinical Trial. JAMA Psychiatry 2021, 78, 951–959. [Google Scholar] [CrossRef]
- Pan, D.; Xu, Y.; Zhang, L.; Su, Q.; Chen, M.; Li, B.; Xiao, Q.; Gao, Q.; Peng, X.; Jiang, B.; et al. Gene expression profile in peripheral blood mononuclear cells of postpartum depression patients. Sci. Rep. 2018, 8, 10139. [Google Scholar] [CrossRef] [Green Version]
- Guintivano, J.; Arad, M.; Gould, T.D.; Payne, J.L.; Kaminsky, Z.A. Antenatal prediction of postpartum depression with blood DNA methylation biomarkers. Mol. Psychiatry 2014, 19, 560–567. [Google Scholar] [CrossRef] [Green Version]
- Lee, V.; Maguire, J. The impact of tonic GABAA receptor-mediated inhibition on neuronal excitability varies across brain region and cell type. Front. Neural Circuits 2014, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Carver, C.M.; Reddy, D.S. Neurosteroid interactions with synaptic and extrasynaptic GABAA receptors: Regulation of subunit plasticity, phasic and tonic inhibition, and neuronal network excitability. Psychopharmacology 2013, 230, 151–188. [Google Scholar] [CrossRef]
- Paul, S.M.; Pinna, G.; Guidotti, A. Allopregnanolone: From molecular pathophysiology to therapeutics. A historical perspective. Neurobiol Stress 2020, 12, 100215. [Google Scholar] [CrossRef]
- Carver, C.M.; Wu, X.; Gangisetty, O.; Reddy, D.S. Perimenstrual-like hormonal regulation of extrasynaptic delta-containing GABAA receptors mediating tonic inhibition and neurosteroid sensitivity. J. Neurosci. 2014, 34, 14181–14197. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.S.; Gong, Q.H.; Li, X.; Moran, M.H.; Bitran, D.; Frye, C.A.; Hsu, F.-C. Withdrawal from 3α-OH-5α-Pregnan-20-One Using a Pseudopregnancy Model Alters the Kinetics of Hippocampal GABAA-Gated Current and Increases the GABAA Receptor α4 Subunit in Association with Increased Anxiety. J. Neurosci. 1998, 18, 5275–5284. [Google Scholar] [CrossRef] [Green Version]
- Gulinello, M.; Smith, S.S. Anxiogenic effects of neurosteroid exposure: Sex differences and altered GABAA receptor pharmacology in adult rats. J. Pharmacol. Exp. Ther. 2003, 305, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Maguire, J.; Ferando, I.; Simonsen, C.; Mody, I. Excitability changes related to GABAA receptor plasticity during pregnancy. J. Neurosci. 2009, 29, 9592–9601. [Google Scholar] [CrossRef] [Green Version]
- Kimmel, M.; Kaminsky, Z.; Payne, J.L. Biomarker or pathophysiology? The role of DNA methylation in postpartum depression. Epigenomics 2013, 5, 473–475. [Google Scholar] [CrossRef] [PubMed]
- Katz, E.R.; Stowe, Z.N.; Newport, D.J.; Kelley, M.E.; Pace, T.W.; Cubells, J.F.; Binder, E.B. Regulation of mRNA expression encoding chaperone and co-chaperone proteins of the glucocorticoid receptor in peripheral blood: Association with depressive symptoms during pregnancy. Psychol. Med. 2012, 42, 943–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boero, G.; Porcu, P.; Morrow, A.L. Pleiotropic actions of allopregnanolone underlie therapeutic benefits in stress-related disease. Neurobiol. Stress 2020, 12, 100203. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.L.; Boero, G.; Porcu, P. A Rationale for Allopregnanolone Treatment of Alcohol Use Disorders: Basic and Clinical Studies. Alcohol. Clin. Exp. Res. 2020, 44, 320–339. [Google Scholar] [CrossRef]
- Morrison, K.E.; Cole, A.B.; Kane, P.J.; Meadows, V.E.; Thompson, S.M.; Bale, T.L. Pubertal adversity alters chromatin dynamics and stress circuitry in the pregnant brain. Neuropsychopharmacology 2020, 45, 1263–1271. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudzinskas, S.A.; Mazzu, M.A.; Schiller, C.E.; Meltzer-Brody, S.; Rubinow, D.R.; Schmidt, P.J.; Goldman, D. Divergent Transcriptomic Effects of Allopregnanolone in Postpartum Depression. Genes 2023, 14, 1234. https://doi.org/10.3390/genes14061234
Rudzinskas SA, Mazzu MA, Schiller CE, Meltzer-Brody S, Rubinow DR, Schmidt PJ, Goldman D. Divergent Transcriptomic Effects of Allopregnanolone in Postpartum Depression. Genes. 2023; 14(6):1234. https://doi.org/10.3390/genes14061234
Chicago/Turabian StyleRudzinskas, Sarah A., Maria A. Mazzu, Crystal Edler Schiller, Samantha Meltzer-Brody, David R. Rubinow, Peter J. Schmidt, and David Goldman. 2023. "Divergent Transcriptomic Effects of Allopregnanolone in Postpartum Depression" Genes 14, no. 6: 1234. https://doi.org/10.3390/genes14061234