
Recent Advances in Genetics and Genomics of Snub-Nosed Monkeys (Rhinopithecus) and Their Implications for Phylogeny, Conservation, and Adaptation

 and
and
Abstract
:1. Introduction
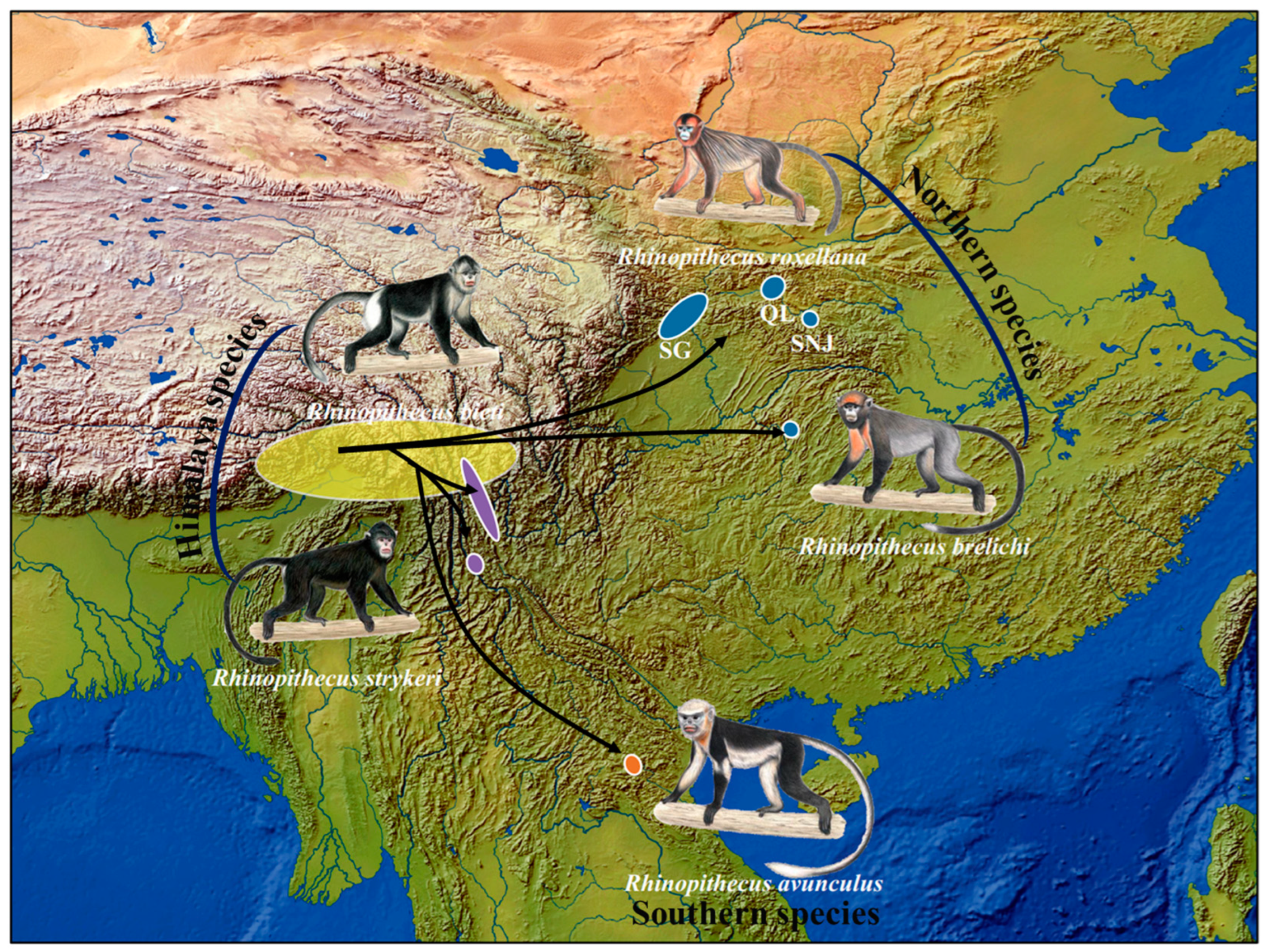
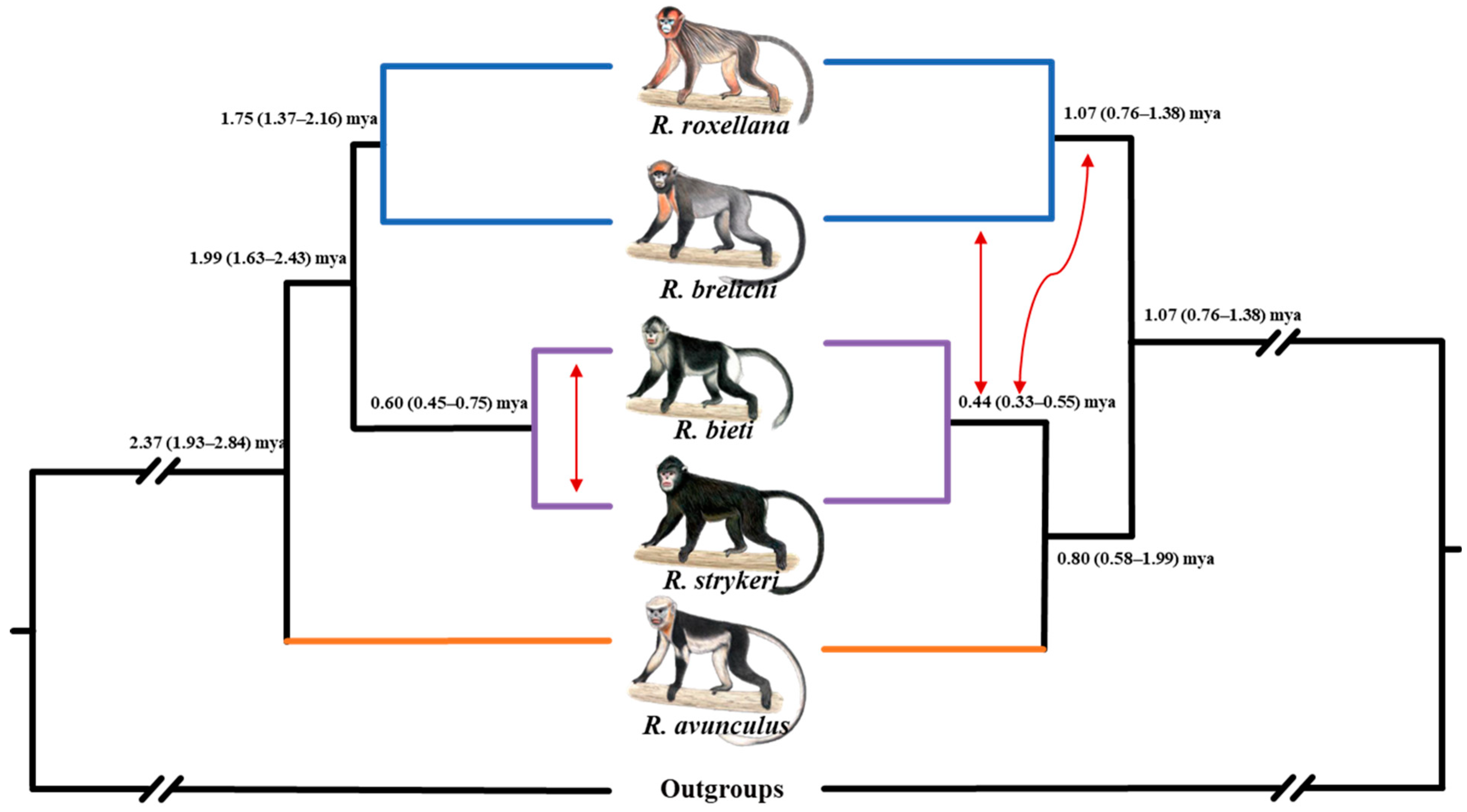
2. Phylogeny and Phylogeography
3. Population Genetics
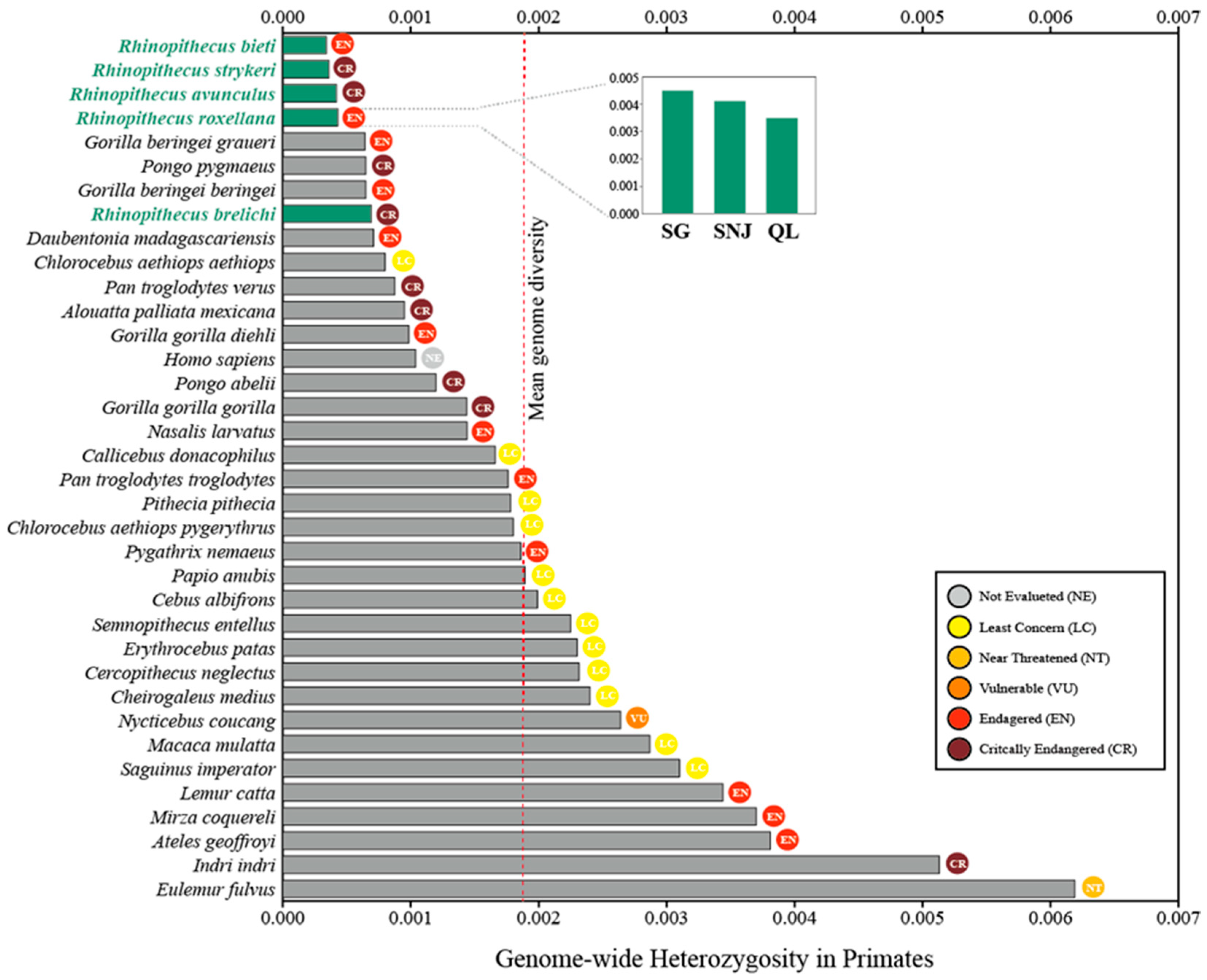
3.1. Genetic Diversity
3.2. Landscape Genetics
3.3. Inbreeding
3.4. Genetic Load
3.5. Demographic History
4. Natural Selection and Adaptation
4.1. Adaptation to Folivory
4.2. Adaptation to High Altitude
5. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yousaf, A.; Liu, J.; Ye, S.; Chen, H. Current progress in evolutionary comparative genomics of great apes. Front. Genet. 2021, 12, 657468. [Google Scholar] [CrossRef]
- Kuderna, L.F.K.; Gao, H.; Janiak, M.C.; Kuhlwilm, M.; Orkin, J.D.; Bataillon, T.; Manu, S.; Valenzuela, A.; Bergman, J.; Rouselle, M.; et al. A global catalog of whole-genome diversity from 233 primate species. Science, 2023; in press. [Google Scholar]
- Delsuc, F.; Brinkmann, H.; Philippe, H. Phylogenomics and the reconstruction of the tree of life. Nat. Rev. Genet. 2005, 6, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, E.F.; Harris, R.A.; Zhang, L.; Raveendran, M.; Kuderna, L.F.K.; Walker, J.A.; Storer, J.M.; Kuhlwilm, M.; Fontsere, C.; Seshadri, L.; et al. Genome-wide coancestry reveals details of ancient and recent male-driven reticulation in baboons. Science, 2023; in press. [Google Scholar]
- Wolfsberger, W.; Battistuzzi, F.; Oleksyk, T. Genomics of adaptation and speciation. Genes 2022, 13, 1187. [Google Scholar] [CrossRef]
- Hendricks, S.; Anderson, E.; Antao, T.; Bernatchez, L.; Forester, B.; Garner, B.; Hand, B.K.; Hohenlohe, P.A.; Kardos, M.; Koop, B.; et al. Recent advances in conservation and population genomics data analysis. Evol. Appl. 2018, 11, 1197–1211. [Google Scholar] [CrossRef]
- Segelbacher, G.; Bosse, M.; Burger, P.; Galbusera, P.; Godoy, J.; Helsen, P.; Hvilsom, C.; Iacolina, L.; Kahrić, A.; Manfrin, C.; et al. New developments in the field of genomic technologies and their relevance to conservation management. Conserv. Genet. 2022, 23, 217–242. [Google Scholar] [CrossRef]
- Theissinger, K.; Fernandes, C.; Formenti, G.; Bista, I.; Berg, P.R.; Bleidorn, C.; Bombarely, A.; Crottini, A.; Gallo, G.; Godoy, J.; et al. How genomics can help biodiversity conservation. Trends Genet. 2023; in press. [Google Scholar] [CrossRef]
- Wang, L.; Wu, J.; Liu, X.; Di, D.; Liang, Y.; Feng, Y.; Zhang, S.; Li, B.G.; Qi, X.G. A high-quality genome assembly for the endangered golden snub-nosed monkey (Rhinopithecus roxellana). Gigascience 2019, 8, giz098. [Google Scholar] [CrossRef]
- Yu, L.; Wang, G.D.; Ruan, J.; Chen, Y.D.; Yang, C.P.; Cao, X.; Wu, H.; Liu, Y.H.; Du, Z.L.; Wang, X.P.; et al. Genomic analysis of snub-nosed monkeys (Rhinopithecus) identifies genes and processes related to high-altitude adaptation. Nat. Genet. 2016, 48, 947–952. [Google Scholar] [CrossRef]
- Shao, Y.; Zhou, L.; Li, F.; Zhao, L.; Zhang, B.; Shao, F.; Chen, J.; Chen, C.; Bi, X.; Zhuang, X.; et al. Phylogenomic analyses provide insights into primate evolution. Science, 2023; in press. [Google Scholar]
- Zhou, X.; Wang, B.; Pan, Q.; Zhang, J.; Kumar, S.; Sun, X.; Liu, Z.; Pan, H.; Lin, Y.; Liu, G.; et al. Whole-genome sequencing of the snub-nosed monkey provides insights into folivory and evolutionary history. Nat. Genet. 2014, 46, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Meng, X.; Liu, Z.; Chang, J.; Wang, B.; Li, M.; Wengel, P.O.T.; Tian, S.; Wen, C.; Wang, Z.; et al. Population genomics reveals low genetic diversity and adaptation to hypoxia in snub-nosed monkeys. Mol. Biol. Evol. 2016, 33, 2670–2781. [Google Scholar] [CrossRef]
- Kuang, W.; Ming, C.; Li, H.; Wu, H.; Frantz, L.; Roos, C.; Zhang, Y.P.; Zhang, C.; Jia, T.; Yang, J.; et al. The origin and population history of the Endangered golden snub-nosed monkey (Rhinopithecus roxellana). Mol. Biol. Evol. 2019, 36, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Kuang, W.; Hu, J.; Wu, H.; Fen, X.; Dai, Q.; Fu, Q.; Xiao, W.; Frantz, L.; Roos, C.; Nadler, T.; et al. Genetic diversity, inbreeding level, and genetic load in endangered snub-nosed monkeys (Rhinopithecus). Front. Genet. 2020, 11, 615926. [Google Scholar] [CrossRef] [PubMed]
- Liedigk, R.; Yang, M.; Jablonski, N.G.; Momberg, F.; Geissmann, T.; Lwin, N.; Hla, T.H.; Liu, Z.; Wong, B.; Li, M.; et al. Evolutionary history of the odd-nosed monkeys and the phylogenetic position of the newly described Myanmar snub-nosed monkey Rhinopithecus strykeri. PLoS ONE 2012, 7, e37418. [Google Scholar] [CrossRef]
- Anandam, M.V.; Bennett, E.L.; Davenport, T.R.B.; Davies, N.J.; Detwiler, K.M.; Engelhardt, A.; Eudey, A.A.; Gadsby, E.L.; Groves, C.P.; Healy, A.; et al. Family Cercopithecidae. In Handbook of the Mammals of the World. Primates; Mittermeier, R.A., Rylands, A.B., Wilson, D.E., Eds.; Lynx Edicions: Barcelona, Spain, 2013; Volume 3, pp. 628–753. [Google Scholar]
- Roos, C.; Boonratana, R.; Supriatna, J.; Fellowes, J.R.; Groves, C.P.; Nash, S.; Rylands, A.; Mittermeier, R. An updated taxonomy and conservation status review of Asian primates. Asian Primates J. 2014, 4, 2–38. [Google Scholar]
- Jablonski, N.G.; Peng, Y.Z. The phylogenetic relationships and classification of the doucs and snub-nosed langurs of China and Vietnam. Folia Primatol. 1993, 60, 36–55. [Google Scholar] [CrossRef]
- Li, M.; Wei, F.; Huang, C.; Pan, R.; de Ruiter, J.R. Phylogeny of snub-nosed monkeys inferred from mitochondrial DNA, cytochrome b, and 12S rRNA sequences. Int. J. Primatol. 2004, 25, 861–873. [Google Scholar] [CrossRef]
- Roos, C.; Thanh, V.N.; Walter, L.; Nadler, T. Molecular systematics of Indochinese primates. Vietn. J. Primatol. 2007, 1, 41–53. [Google Scholar]
- Geissmann, T.; Lwin, N.; Aung, S.S.; Aung, T.N.; Aung, Z.M.; Hla, T.H.; Grindley, M.; Momberg, F. A new species of snub-nosed monkey, genus Rhinopithecus Milne-Edwards, 1872 (Primates, Colobinae), from northern Kachin state, northeastern Myanmar. Am. J. Primatol. 2011, 73, 96–107. [Google Scholar] [CrossRef]
- Jablonski, N.G. The Natural History of the Doucs and Snub-nosed Monkeys. Singap. World Sci. 1998, 110, 13–52. [Google Scholar]
- Roos, C.; Zinner, D.; Kubatko, L.; Schwarz, C.; Yang, M.; Meyer, D.; Nash, S.D.; Xing, J.; Batzer, M.A.; Brameier, M.; et al. Nuclear versus mitochondrial DNA: Evidence for hybridization in colobine monkeys. BMC Evol. Biol. 2011, 11, 77. [Google Scholar] [CrossRef]
- Ji, X.; Youlatos, D.; Jablonski, N.G.; Pan, R.; Zhang, C.; Li, P.; Tang, M.; Yu, T.; Li, W.; Deng, C.; et al. Oldest colobine calcaneus from East Asia (Zhaotong, Yunnan, China). J. Hum. Evol. 2020, 147, 102866. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.R.; Jablonski, N.G. The age and geographical distribution of fossil cercopithecids in China. Hum. Evol. 1987, 2, 59–69. [Google Scholar] [CrossRef]
- Jablonski, N.G.; Pan, Y.R. The evolution and palaeobiogeography of monkeys in China. In The Palaeoenvironment of East Asia from the Mid-Tertiary; Whyte, P., Aigner, J., Jablonski, N.G., Taylor, G., Walker, D., Eds.; Centre of Asian Studies: Hong Kong, 1988; Volume 2, pp. 849–867. [Google Scholar]
- Pan, R.L.; Jablonski, N.G. Scaling of limb proportions and limb bone diameters in three species of Chinese snub-nosed langurs (Genus Rhinopithecus). Folia Primatol. 1993, 60, 56–62. [Google Scholar] [CrossRef]
- Jablonski, N.G.; Ruliang, P. Sexual dimorphism in the snub-nosed langurs (Colobinae: Rhinopithecus). Am. J. Phys. Anthropol. 1995, 96, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Pan, R.; Oxnard, C.E. Extinction of snub-nosed monkeys in China during the past 400 years. Int. J. Primatol. 2002, 23, 1227–1244. [Google Scholar] [CrossRef]
- Nüchel, J.; Bøcher, P.K.; Xiao, W.; Zhu, A.X.; Svenning, J.C. Snub-nosed monkeys (Rhinopithecus): Potential distribution and its implication for conservation. Anim. Biodivers. Conserv. 2018, 27, 1517–1538. [Google Scholar] [CrossRef]
- Li, J.; Li, D.; Xue, Y.; Wu, B.; He, X.; Liu, F. Identifying potential refugia and corridors under climate change: A case study of endangered Sichuan golden monkey (Rhinopithecus roxellana) in Qinling Mountains, China. Am. J. Primatol. 2018, 80, e22929. [Google Scholar] [CrossRef]
- Zhao, X.; Ren, B.; Li, D.; Xiang, Z.; Garber, P.A.; Li, M. Effects of habitat fragmentation and human disturbance on the population dynamics of the Yunnan snub-nosed monkey from 1994 to 2016. PeerJ 2019, 7, e6633. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lin, A.K.; Garber, P.A.; Huang, Z.; Tian, Y.; Behie, A.; Momberg, F.; Grueter, C.C.; Li, W.; Lwin, N.; et al. The 10th anniversary of the scientific description of the black snub-nosed monkey (Rhinopithecus strykeri): It is time to initiate a set of new management strategies to save this critically endangered primate from extinction. Am. J. Primatol. 2022, 84, e23372. [Google Scholar] [CrossRef]
- Guo, Y.; Ren, B.; Dai, Q.; Zhou, J.; Garber, P.A.; Zhou, J. Habitat estimates reveal that there are fewer than 400 Guizhou snub-nosed monkeys, Rhinopithecus brelichi, remaining in the wild. Glob. Ecol. Conserv. 2020, 24, e01181. [Google Scholar] [CrossRef]
- Peng, Y.Z.; Zhang, Y.P.; Ye, Z.Z.; Liu, R.L. Study on the stomaches in three species of snub-nosed monkeys. Zool. Res. 1983, 4, 167–176. [Google Scholar]
- Grueter, C.C.; Li, D.Y.; Feng, S.K.; Ren, B.P. Niche partitioning between sympatric rhesus macaques and Yunnan snub-nosed monkeys at Baimaxueshan Nature Reserve, China. Zool. Res. 2010, 31, 516–522. [Google Scholar] [CrossRef]
- Chiou, K.; Janiak, M.; Schneider-Crease, I.; Sen, S.; Ayele, F.; Chuma, I.; Knauf, F.; Lemma, A.; Signore, A.V.; D’Ippolito, A.M.; et al. Genomic signatures of high-altitude adaptation and chromosomal polymorphism in geladas. Nat. Ecol. Evol. 2022, 6, 630–643. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Kirkpatrick, R.C.; Zhong, T.; Xiao, L. Report on the distribution, population, and ecology of the Yunnan snub-nosed monkey (Rhinopithecus bieti). Primates 1994, 35, 241–250. [Google Scholar] [CrossRef]
- Liu, Z.; Ren, B.; Wei, F.; Long, Y.; Hao, Y.; Li, M. Phylogeography and population structure of the Yunnan snub-nosed monkey (Rhinopithecus bieti) inferred from mitochondrial control region DNA sequence analysis. Mol. Ecol. 2007, 16, 3334–3349. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ren, B.; Wu, R.; Zhao, L.; Hao, Y.; Wang, B.; Wei, F.; Long, Y.; Li, M. The effect of landscape features on population genetic structure in Yunnan snub-nosed monkeys (Rhinopithecus bieti) implies an anthropogenic genetic discontinuity. Mol. Ecol. 2009, 18, 3831–3846. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, G.; Roos, C.; Wang, Z.; Xiang, Z.F.; Zhu, P.; Wang, B.; Ren, B.; Shi, F.; Pan, H.; et al. Implications of genetics and current protected areas for conservation of 5 endangered primates in China. Conserv. Biol. 2015, 29, 1508–1517. [Google Scholar] [CrossRef]
- Wang, X.; Yu, L.; Roos, C.; Ting, N.; Chen, C.; Wang, J.; Zhang, Y. Phylogenetic relationships among the colobine monkeys revisited: New insights from analyses of complete mt genomes and 44 nuclear non-coding markers. PLoS ONE 2012, 7, e36274. [Google Scholar] [CrossRef]
- Hong, Y.; Duo, H.; Hong, J.; Yang, J.; Liu, S.; Yu, L.; Yi, T. Resequencing and comparison of whole mitochondrial genome to gain insight into the evolutionary status of the Shennongjia golden snub-nosed monkey (SNJ R. roxellana). Ecol. Evol. 2017, 7, 4456–4464. [Google Scholar] [CrossRef]
- Rogers, J.; Raveendran, M.; Harris, R.; Mailund, T.; Leppälä, K.; Athanasiadis, G.; Schierup, M.H.; Cheng, J.; Munch, K.; Walker, J.A.; et al. The comparative genomics and complex population history of Papio baboons. Sci. Adv. 2019, 5, eaau6947. [Google Scholar] [CrossRef] [PubMed]
- Haus, T.; Roos, C.; Zinner, D. Discordance between spatial distributions of Y-chromosomal and mitochondrial haplotypes in African green monkeys (Chlorocebus spp.): A result of introgressive hybridization or cryptic diversity? Int. J. Primatol. 2013, 34, 986–999. [Google Scholar] [CrossRef]
- Warren, W.C.; Jasinska, A.J.; García-Pérez, R.; Svardal, H.; Tomlinson, C.; Rocchi, M.; Archidiacono, N.; Capozzi, O.; Minx, P.; Montague, M.J.; et al. The genome of the vervet (Chlorocebus aethiops sabaeus). Genome Res. 2015, 25, 1921–1933. [Google Scholar] [CrossRef]
- Pfeifer, S. The demographic and adaptive history of the African green monkey. Mol. Biol. Evol. 2017, 34, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, Z.; Gou, J.; Ren, B.; Pan, R.; Su, Y.; Funk, S.M.; Wei, F. Phylogeography and population structure of the golden monkeys (Rhinopithecus roxellana): Inferred from mitochondrial DNA sequences. Am. J. Primatol. 2007, 69, 1195–1209. [Google Scholar] [CrossRef]
- Luo, M.; Liu, Z.; Pan, H.; Zhao, L.; Funk, S.M.; Li, M. Historical geographic dispersal of the golden snub-nosed monkey (Rhinopithecus roxellana) and the influence of climatic oscillations. Am. J. Primatol. 2012, 74, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Pan, H.; Liu, Z.; Li, M. Balancing selection and genetic drift at major histocompatibility complex class II genes in isolated populations of golden snub-nosed monkey (Rhinopithecus roxellana). BMC Evol. Biol. 2012, 12, 207. [Google Scholar] [CrossRef]
- Frankham, R.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Allendorf, F.W.; Hohenlohe, P.A.; Luikart, G. Genomics and the future of conservation genetics. Nat. Rev. Genet. 2010, 11, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Kohn, M.H.; Murphy, W.J.; Ostrander, E.A.; Wayne, R.K. Genomics and conservation genetics. Trends Ecol. Evol. 2006, 21, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Hu, H.; Meng, S.; Meng, Z.; Fu, Y.; Zhang, Y.P. A High Polymorphism Level in Rhinopithecus roxellana. Int. J. Primatol. 2009, 30, 337–351. [Google Scholar] [CrossRef]
- Chang, Z.F.; Luo, M.F.; Liu, Z.J.; Yang, J.Y.; Xiang, Z.F.; Li, M.; Vigilant, L. Human influence on the population decline and loss of genetic diversity in a small and isolated population of Sichuan snub-nosed monkeys (Rhinopithecus roxellana). Genetica 2012, 140, 105–114. [Google Scholar] [CrossRef]
- Yang, M.; Yang, Y.; Cui, D.; Fickenscher, G.; Zinner, D.; Roos, C.; Brameier, M. Population genetic structure of Guizhou snub-nosed monkeys (Rhinopithecus brelichi) as inferred from mitochondrial control region sequences, and comparison with R. roxellana and R. bieti. Am. J. Phys. Anthropol. 2012, 147, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kolleck, J.; Yang, M.; Zinner, D.; Roos, C. Genetic diversity in endangered Guizhou snub-nosed monkeys (Rhinopithecus brelichi): Contrasting results from microsatellite and mitochondrial DNA data. PLoS ONE 2013, 8, e73647. [Google Scholar] [CrossRef]
- Huang, K.; Guo, S.; Cushman, S.A.; Dunn, D.W.; Qi, X.; Hou, R.; Zhang, J.; Li, Q.; Zhang, Q.; Shi, Z.; et al. Population structure of the golden snub-nosed monkey Rhinopithecus roxellana in the Qinling Mountains, central China. Integr. Zool. 2016, 11, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Ang, A.; Srivathsan, A.; Meier, R.; Luu, T.B.; Le, Q.K.; Covert, H. No evidence for mitochondrial genetic variability in the largest population of critically endangered Tonkin snub-nosed monkeys in Vietnam. Primates 2016, 57, 449–453. [Google Scholar] [CrossRef]
- Zhang, B.; Hu, H.Y.; Song, C.; Huang, K.; Dunn, D.W.; Yang, X.; Wang, X.W.; Zhao, H.T.; Wang, C.L.; Zhang, P.; et al. MHC-Based Mate Choice in Wild Golden Snub-Nosed Monkeys. Front Genet. 2020, 11, 609414. [Google Scholar] [CrossRef]
- Li, Y.; Huang, K.; Tang, S.; Feng, L.; Yang, J.; Li, Z.; Li, B. Genetic Structure and Evolutionary History of Rhinopithecus roxellana in Qinling Mountains, Central China. Front. Genet. 2020, 11, 611914. [Google Scholar] [CrossRef]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Keller, L.F.; Waller, D.M. Inbreeding effects in wild populations. Trends Ecol. Evol. 2002, 17, 230–241. [Google Scholar] [CrossRef]
- Zoonomia, C. A comparative genomics multitool for scientific discovery and conservation. Nature 2020, 587, 240–245. [Google Scholar] [CrossRef]
- Manel, S.; Schwartz, M.K.; Luikart, G.; Taberlet, P. Landscape genetics: Combining landscape ecology and population genetics. Trends Ecol. Evol. 2003, 18, 189–197. [Google Scholar] [CrossRef]
- McRae, B.H.; Beier, P. Circuit theory predicts gene flow in plant and animal populations. Proc. Natl. Acad. Sci. USA 2007, 104, 19885–19890. [Google Scholar] [CrossRef]
- Bohonak, A.J.; Vandergast, A.G. The value of DNA sequence data for studying landscape genetics. Mol. Ecol. 2011, 20, 2477–2479. [Google Scholar] [CrossRef]
- Roos, C.; Zinner, D. Primate conservation genetics. In The International Encyclopedia of Primatology; Fuentes, A., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017. [Google Scholar]
- Holderegger, R.; Wagner, H.H. Landscape Genetics. BioScience 2008, 58, 199–207. [Google Scholar] [CrossRef]
- Manel, S.; Holderegger, R. Ten years of landscape genetics. Trends Ecol. Evol. 2013, 28, 614–621. [Google Scholar] [CrossRef]
- Zhang, P.; Hu, K.J.; Yang, B.; Yang, D.H. Snub-nosed monkeys (Rhinopithecus spp.): Conservation challenges in the face of environmental uncertainty. Sci. Bull. 2016, 61, 345–348. [Google Scholar] [CrossRef]
- Li, L.; Xue, Y.; Wu, G.; Li, D.; Giraudoux, P. Potential habitat corridors and restoration areas for the black-and-white snub-nosed monkey Rhinopithecus bieti in Yunnan, China. Oryx 2015, 49, 719–726. [Google Scholar] [CrossRef]
- Zhao, X.; Li, X.; Zhang, Z.; Garber, P.A.; Yu, M.; Qiao, H.; Li, M. Differential response to climate change and human activities in three lineages of Sichuan snub-nosed monkeys (Rhinopithecus roxellana). Divers. Distrib. 2022, 28, 2416–2428. [Google Scholar] [CrossRef]
- Barrett, S.C.H.; Charlesworth, D. Effects of a change in the level of inbreeding on the genetic load. Nature 1991, 352, 522–524. [Google Scholar] [CrossRef]
- Charlesworth, D.; Willis, J.H. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef]
- Hedrick, P.W.; Garcia-Dorado, A. Understanding inbreeding depression, purging, and genetic rescue. Trends Ecol. Evol. 2016, 31, 940–952. [Google Scholar] [CrossRef] [PubMed]
- Bertorelle, G.; Raffini, F.; Bosse, M.; Bortoluzzi, C.; Iannucci, A.; Trucchi, E.; Morales, H.E.; van Oosterhout, C. Genetic load: Genomic estimates and applications in non-model animals. Nat. Rev. Genet. 2022, 23, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Plough, L.V. Genetic load in marine animals: A review. Curr. Zool. 2016, 62, 567–579. [Google Scholar] [CrossRef]
- Van Oosterhout, C. Mutation load is the spectre of species conservation. Nat. Ecol. Evol. 2020, 4, 1004–1006. [Google Scholar] [CrossRef]
- Van der Valk, T.; de Manuel, M.; Marquès-Bonet, T.; Guschanski, K. Estimates of genetic load in small populations suggest extensive purging of deleterious alleles. bioRxiv 2019. [Google Scholar] [CrossRef]
- Xue, Y.; Prado-Martinez, J.; Sudmant, P.; Narasimhan, V.; Ayub, Q.; Szpak, M.; Frandsen, P.; Chen, Y.; Yngvadottir, B.; Cooper, D.V.; et al. Mountain gorilla genomes reveal the impact of longterm population decline and inbreeding. Science 2015, 348, 242–245. [Google Scholar] [CrossRef]
- Gomides, L.F.; Duarte, I.D.; Ferreira, R.G.; Perez, A.C.; Francischi, J.N.; Klein, A. Proteinase-activated receptor-4 plays a major role in the recruitment of neutrophils induced by trypsin or carrageenan during pleurisy in mice. Pharmacology 2012, 89, 275–282. [Google Scholar] [CrossRef]
- Hossain, M.B.; Li, H.q.; Hedmer, M.; Tinnerberg, H.; Broberg, K. Exposure to welding fumes is associated with hypomethylation of the F2RL3 gene: A cardiovascular disease marker. Occup. Environ. Med. 2015, 72, 845–851. [Google Scholar] [CrossRef]
- Sheng, C.; Wang, Z.; Yao, C.; Chen, H.M.; Kan, G.; Wang, D.; Chen, H.; Chen, S. CALML6 controls TAK1 ubiquitination and confers protection against acute inflammation. J. Immunol. 2020, 204, 3008–3018. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Sheng, C.; Yao, C.; Chen, H.; Wang, D.; Chen, S. The EF-Hand protein CALML6 suppresses antiviral innate immunity by impairing IRF3 dimerization. Cell Rep. 2019, 26, 1273–1285. [Google Scholar] [CrossRef] [PubMed]
- Steiner, C.C.; Putnam, A.S.; Hoeck, P.E.; Ryder, O.A. Conservation genomics of threatened animal species. Annu. Rev. Anim. Biosci. 2013, 1, 261–281. [Google Scholar] [CrossRef]
- Blischak, P.D.; Barker, M.S.; Gutenkunst, R.N. Inferring the Demographic History of Inbred Species from Genome-Wide SNP Frequency Data. Mol. Biol. Evol. 2020, 37, 2124–2136. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Meng, S.J.; Men, Z.M.; Fu, Y.X.; Zhang, Y.P. Genetic diversity and population history of golden monkeys (Rhinopithecus roxellana). Genetics 2003, 164, 269–275. [Google Scholar] [CrossRef]
- Pan, D.; Fu, Y.; Zhang, Y. Population genetics of Rhinopithecus bieti: A study of the mitochondrial control region. Hereditas 2006, 28, 432–436. [Google Scholar] [CrossRef]
- Clayton, J.B.; Gomez, A.; Amato, K.; Knights, D.; Travis, D.A.; Blekhman, R.; Knight, R.; Leigh, S.; Stumpf, R.; Wolf, T.; et al. The gut microbiome of nonhuman primates: Lessons in ecology and evolution. Am. J. Primatol. 2018, 80, e22867. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Rosenberg, H.F. Adaptive evolution of a duplicated pancreatic ribonuclease gene in a leaf-eating monkey. Nat. Genet. 2002, 30, 411–415. [Google Scholar] [CrossRef]
- Zhang, J. Parallel adaptive origins of digestive RNases in Asian and African leaf monkeys. Nat Genet. 2006, 38, 819–823. [Google Scholar] [CrossRef]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef]
- Ezenwa, V.O.; Gerardo, N.M.; Inouye, D.W.; Medina, M.; Xavier, J.B. Microbiology. Animal behavior and the microbiome. Science 2012, 338, 198–199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, D.; Wang, L.; Hao, J.; Wang, J.; Zhou, X.; Wang, W.; Qiu, Q.; Huang, X.; Zhou, J.; et al. Convergent evolution of rumen microbiomes in high-altitude mammals. Curr. Biol. 2016, 26, 1873–1879. [Google Scholar] [CrossRef] [PubMed]
- Barnard, E.A. Biological function of pancreatic ribonuclease. Nature 1969, 221, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Beintema, J.J. The primary structure of langur (Presbytis entellus) pancreatic ribonuclease: Adaptive features in digestive enzymes in mammals. Mol. Biol. Evol. 1990, 7, 470–477. [Google Scholar] [CrossRef]
- Hale, V.L.; Tan, C.L.; Niu, K.; Yang, Y.; Knight, R.; Zhang, Q.; Cui, D.; Amato, K.R. Diet versus phylogeny: A comparison of gut microbiota in captive colobine monkey species. Microb. Ecol. 2018, 75, 515–527. [Google Scholar] [CrossRef]
- Hale, V.L.; Tan, C.L.; Niu, K.; Yang, Y.; Zhang, Q.; Knight, R.; Amato, K.R. Gut microbiota in wild and captive Guizhou snub-nosed monkeys, Rhinopithecus brelichi. Am. J. Primatol. 2019, 81, e22989. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Z.; Pan, H.; Qi, J.; Li, D.; Zhang, L.; Shen, Y.; Xiang, Z.; Li, M. Captivity influences the gut microbiome of Rhinopithecus roxellana. Front. Microbiol. 2021, 7, 763022. [Google Scholar] [CrossRef]
- Liu, R.; Amato, K.; Hou, R.; Gomez, A.; Dunn, D.W.; Zhang, J.; Garber, P.A.; Chapman, C.A.; Righini, N.; He, G.; et al. Specialized digestive adaptations within the hindgut of a colobine monkey. Innovation 2022, 17, 100207. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Wang, Z.; Zhang, M.; Wang, S.; Xiang, Z.; Pan, H.; Li, M. The relationship between gut microbiome and bile acids in primates with diverse diets. Front. Microbiol. 2022, 11, 899102. [Google Scholar] [CrossRef]
- Li, H.; Xia, W.; Liu, X.; Wang, X.; Liu, G.; Chen, H.; Zhu, L.; Li, D. Food provisioning results in functional, but not compositional, convergence of the gut microbiomes of two wild Rhinopithecus species: Evidence of functional redundancy in the gut microbiome. Sci. Total Environ. 2023, 858, 159957. [Google Scholar] [CrossRef]
- Zhao, G.; Qi, M.; Wang, Q.; Hu, C.; Li, X.; Chen, Y.; Yang, J.; Yu, H.; Chen, H.; Guo, A. Gut microbiome variations in Rhinopithecus roxellanae caused by changes in the environment. BMC Genom. 2023, 24, 62. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, X.; Wang, Z.; Mao, S.; Zhang, J.; Li, M.; Pan, H. Metatranscriptomic analyses Reveal important roles of the gut microbiome in primate dietary adaptation. Genes 2023, 14, 228. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Zhang, J.L.; Pan, H.J.; Chen, Y.X.; Mao, S.X.; Qi, J.W.; Shen, Y.; Zhang, M.Y.; Xiang, Z.F.; Li, M. Unique characteristics of gut microbiota in black snub-nosed monkeys (Rhinopithecus strykeri) reveal an enzymatic mechanism of adaptation to dietary vegetation. Zool. Res. 2023, 44, 357–360. [Google Scholar] [CrossRef]
- Yu, L.; Wang, X.; Ting, N.; Zhang, Y. Mitogenomic analysis of Chinese snub-nosed monkeys: Evidence of positive selection in NADH dehydrogenase genes in high-altitude adaptation. Mitochondrion 2011, 11, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Fan, Z.; Han, E.; Hou, R.; Zhang, L.; Galaverni, M.; Huang, J.; Liu, H.; Silva, P.; Li, P.; et al. Hypoxia adaptations in the grey wolf (Canis lupus chanco) from Qinghai-Tibet Plateau. PLoS Genet. 2014, 10, e1004466. [Google Scholar] [CrossRef]
- Qiu, Q.; Zhang, G.; Ma, T.; Qian, W.; Wang, J.; Ye, Z.; Cao, C.; Hu, Q.; Larkin, D.M.; Auvil, L.; et al. The yak genome and adaptation to life at high altitude. Nat. Genet. 2012, 44, 946–949. [Google Scholar] [CrossRef]
- Cai, Q.; Qian, X.; Lang, Y.; Luo, Y.; Xu, J.; Pan, S.; Hui, Y.; Gou, C.; Cai, Y.; Hao, M.; et al. Genome sequence of ground tit Pseudopodoces humilis and its adaptation to high altitude. Genome Biol. 2013, 14, R29. [Google Scholar] [CrossRef]
- Mable, B.K. Conservation of adaptive potential and functional diversity: Integrating old and new approaches. Conserv. Genet. 2019, 20, 89–100. [Google Scholar] [CrossRef]
- Wold, J.; Koepfli, K.P.; Galla, S.J.; Eccles, D.; Hogg, C.J.; Le Lec, M.F.; Guhlin, J.; Santure, A.W.; Steeves, T.E. Expanding the conservation genomics toolbox: Incorporating structural variants to enhance genomic studies for species of conservation concern. Mol. Ecol. 2021, 30, 5949–5965. [Google Scholar] [CrossRef] [PubMed]
- Leypold, N.A.; Speicher, M.R. Evolutionary conservation in noncoding genomic regions. Trends Genet. 2021, 37, 903–918. [Google Scholar] [CrossRef]
- Snyder-Mackler, N.; Majoros, W.H.; Yuan, M.; Shaver, A.O.; Gordon, J.B.; Kopp, G.H.; Schlebusch, S.A.; Wall, J.D.; Alberts, S.C.; Mukherjee, S.; et al. Efficient Genome-Wide Sequencing and Low-Coverage Pedigree Analysis from Noninvasively Collected Samples. Genetics 2016, 203, 699–714. [Google Scholar] [CrossRef] [PubMed]
- White, L.C.; Fontsere, C.; Lizano, E.; Hughes, D.A.; Angedakin, S.; Arandjelovic, M.; Granjon, A.C.; Hans, J.B.; Lester, J.D.; Rabanus-Wallace, M.T.; et al. A roadmap for high-throughput sequencing studies of wild animal populations using noninvasive samples and hybridization capture. Mol. Ecol. Resour. 2019, 19, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Fontsere, C.; Alvarez-Estape, M.; Lester, J.; Arandjelovic, M.; Kuhlwilm, M.; Dieguez, P.; Agbor, A.; Angedakin, S.; Ayuk Ayimisin, E.; Bessone, M.; et al. Maximizing the acquisition of unique reads in noninvasive capture sequencing experiments. Mol. Ecol. Resour. 2021, 21, 745–761. [Google Scholar] [CrossRef] [PubMed]
- Orkin, J.D.; Montague, M.J.; Tejada-Martinez, D.; Manuel, M.D.; Campo, J.C.; Hernandez, S.C.; Fiore, A.D.; Fontsere, C.; Hodgson, J.A.; Janiak, M.C.; et al. The genomics of ecological flexibility, large brains, and long lives in capuchin monkeys revealed with fecalFACS. Proc. Natl. Acad. Sci. USA 2021, 118, e2010632118. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuang, W.; Zinner, D.; Li, Y.; Yao, X.; Roos, C.; Yu, L. Recent Advances in Genetics and Genomics of Snub-Nosed Monkeys (Rhinopithecus) and Their Implications for Phylogeny, Conservation, and Adaptation. Genes 2023, 14, 985. https://doi.org/10.3390/genes14050985
Kuang W, Zinner D, Li Y, Yao X, Roos C, Yu L. Recent Advances in Genetics and Genomics of Snub-Nosed Monkeys (Rhinopithecus) and Their Implications for Phylogeny, Conservation, and Adaptation. Genes. 2023; 14(5):985. https://doi.org/10.3390/genes14050985
Chicago/Turabian StyleKuang, Weimin, Dietmar Zinner, Yuan Li, Xueqin Yao, Christian Roos, and Li Yu. 2023. "Recent Advances in Genetics and Genomics of Snub-Nosed Monkeys (Rhinopithecus) and Their Implications for Phylogeny, Conservation, and Adaptation" Genes 14, no. 5: 985. https://doi.org/10.3390/genes14050985