
Molecular Survey of Rodent-Borne Infectious Agents in the Ferlo Region, Senegal

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
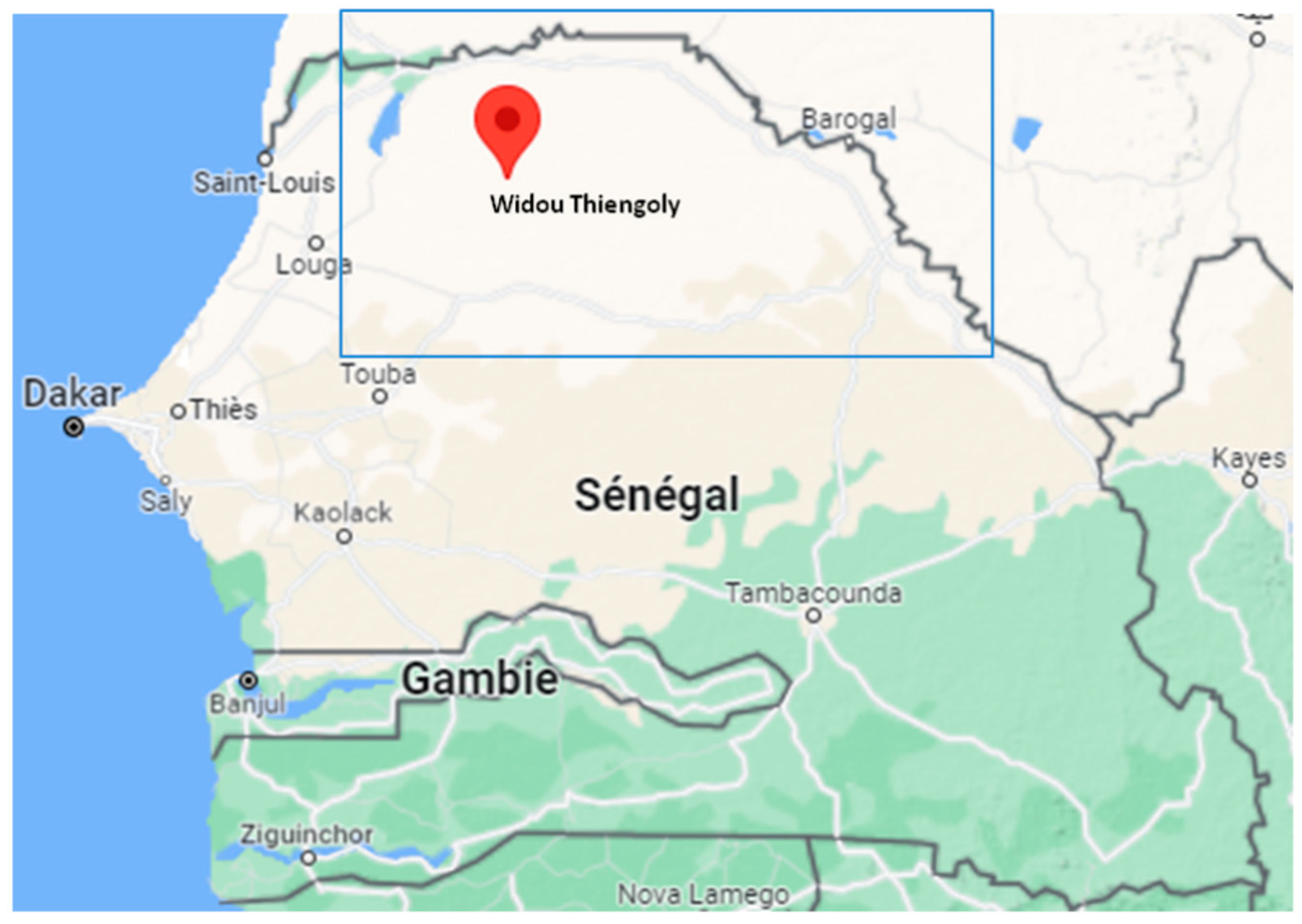
2.1. Study Area and Sample Design
2.2. Ethical Statement Regarding Fieldwork
2.3. DNA Extraction
2.4. PCR Amplification
2.5. Sequencing and Phylogenetic Analysis
2.6. Statistical Analysis
3. Results
3.1. Samples Included in the Study
3.2. Molecular Detection of Microorganisms (Bacteria and Protozoa)
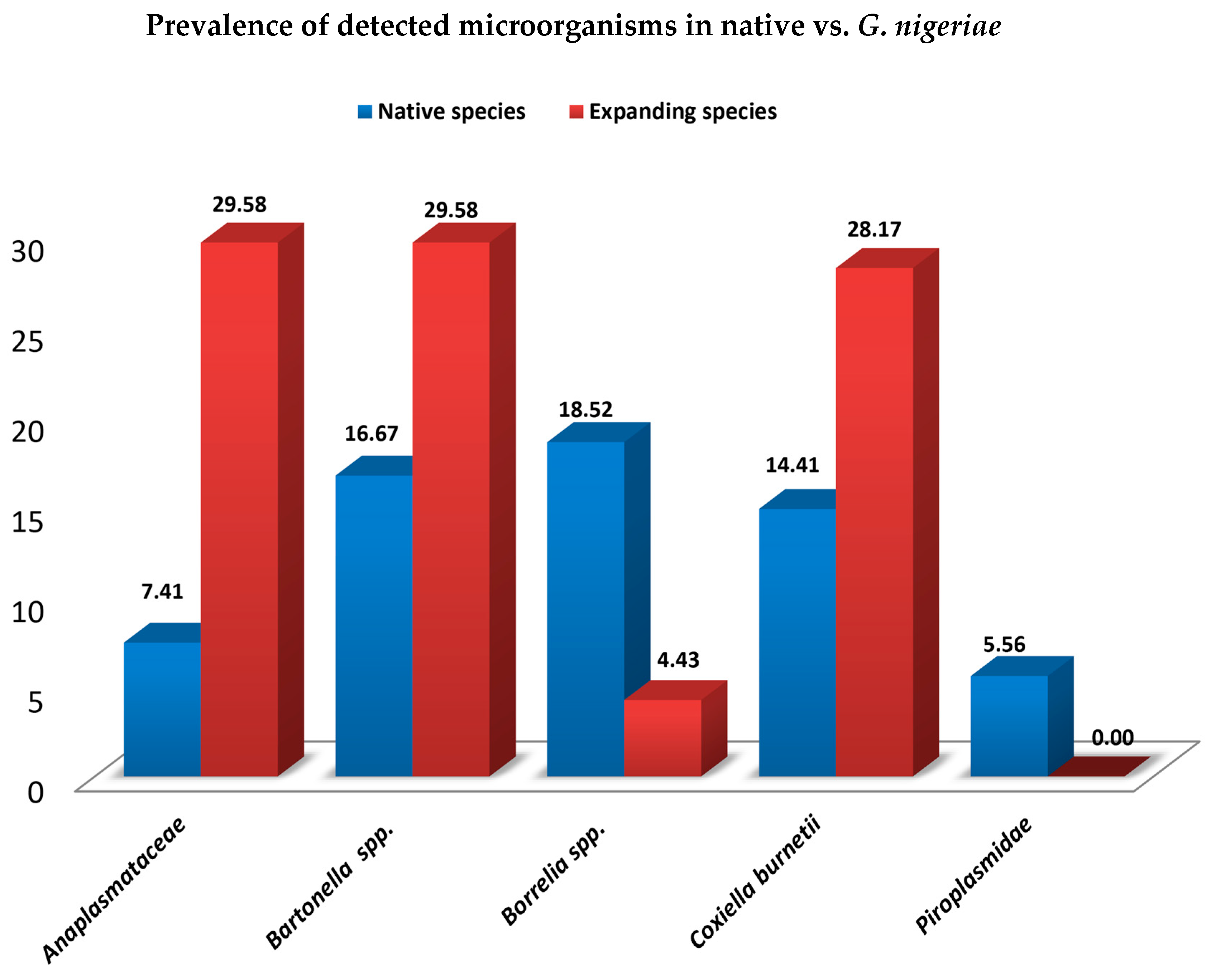
3.3. Prevalence of Microorganisms by Sex, Age, Status and Season
3.4. Coinfections with Multiple Microorganisms
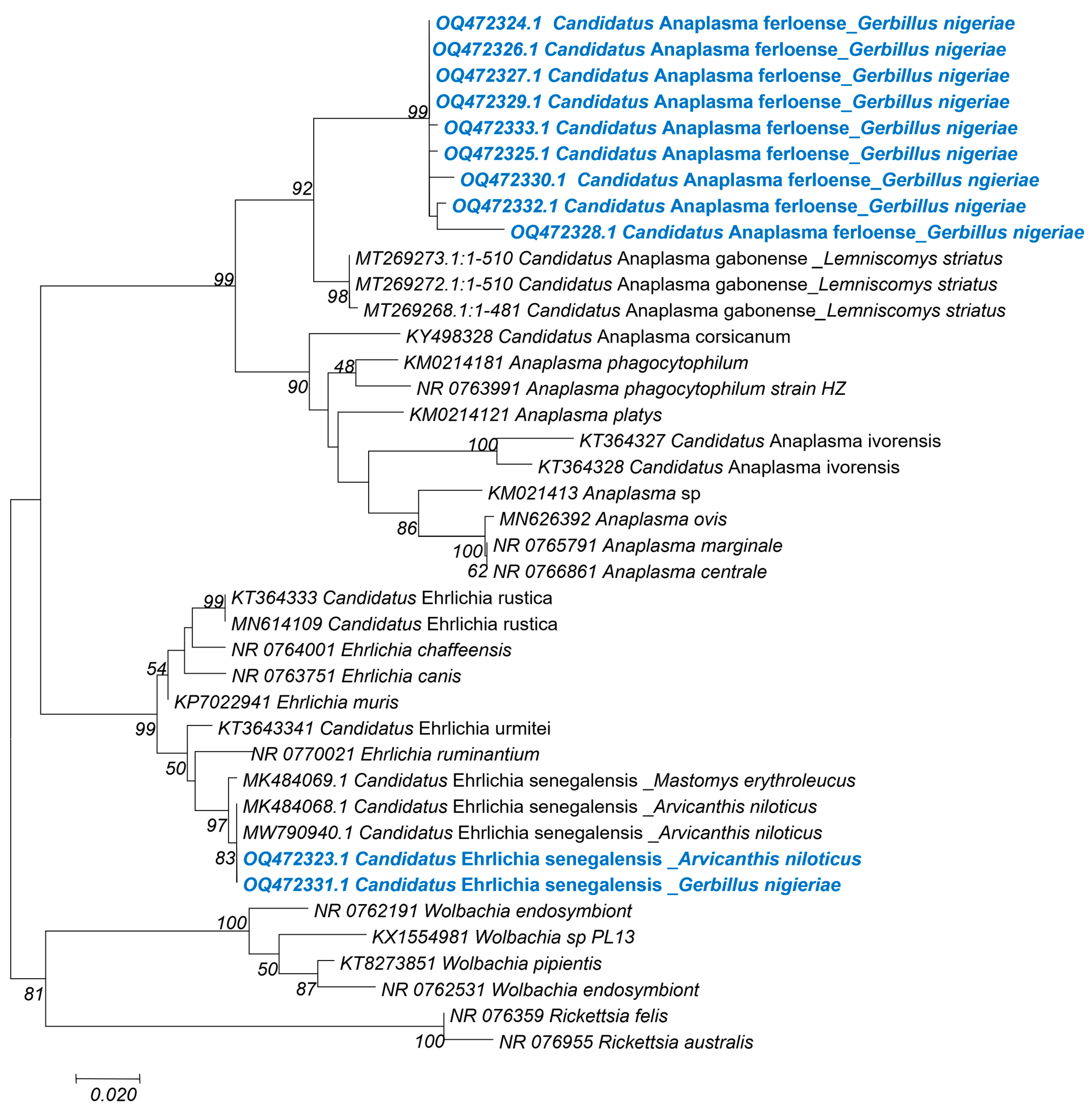
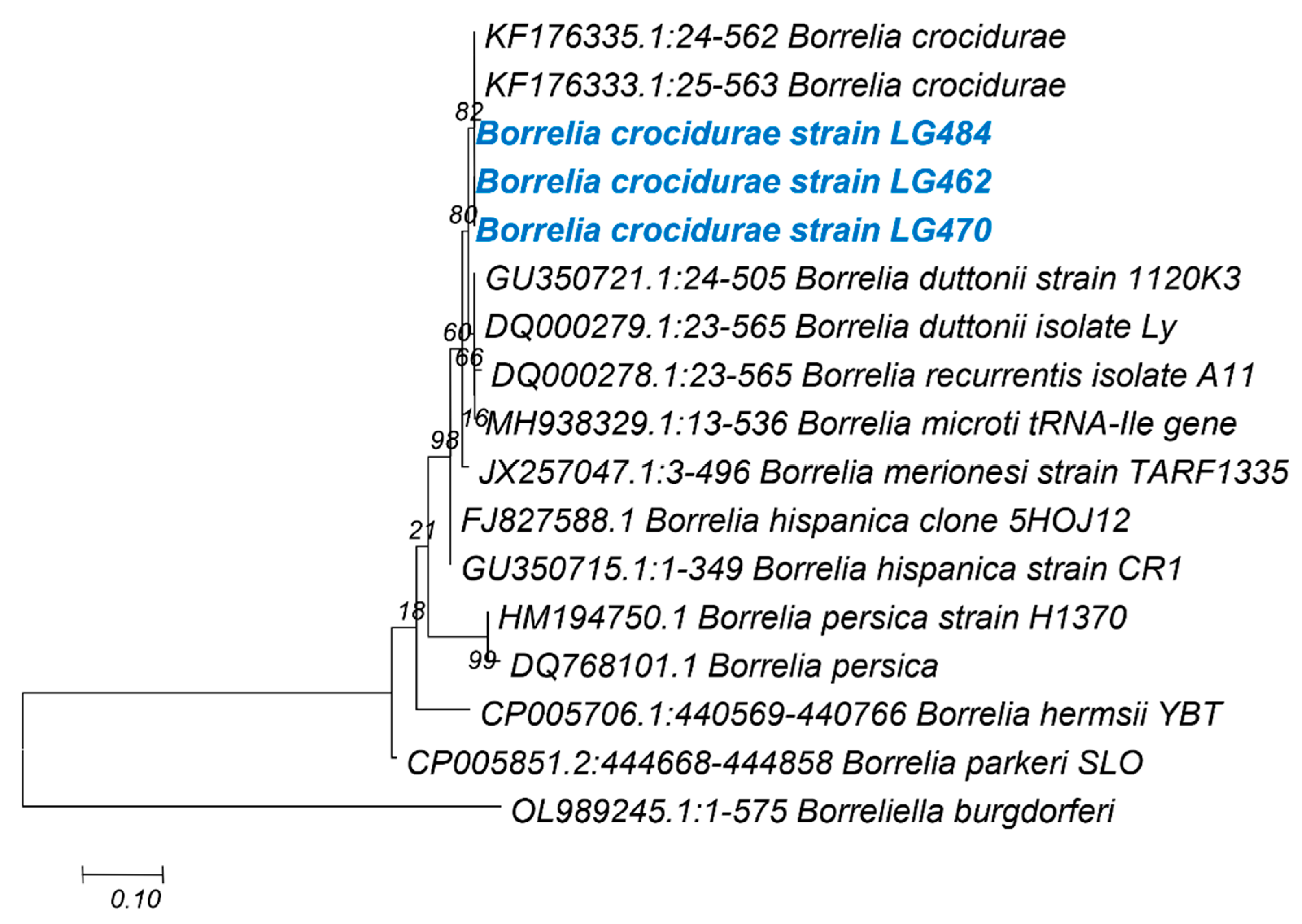
3.5. Phylogenetic Analysis for the Taxonomic Description of Detected Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burgin, C.J.; Colella, J.P.; Kahn, P.L.; Upham, N.S. How many species of mammals are there? J. Mammal. 2018, 99, 1–14. [Google Scholar] [CrossRef]
- Carleton, M.D.; Musser, G.G. Order Rodentia. In Mammal Species of the World, 3rd ed.; Wilson, D.E., Reeder, D.M., Eds.; The Johns Hopkins University Press: Baltimore, MD, USA, 2005; pp. 745–752. [Google Scholar]
- Granjon, L.; Duplantier, J.M. Les Rongeurs de l’Afrique Sahélo-Soudanienne; Publications Scientifiques du Muséum; IRD: Marseille, France, 2009; 215p. [Google Scholar]
- Meerburg, B.G.; Singleton, G.R.; Kijlstra, A. Rodent-borne diseases and their risks for public health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef] [PubMed]
- Han, B.A.; Schmidt, J.P.; Bowden, S.E.; Drake, J.M. Rodent reservoirs of future zoonotic diseases. Proc. Natl. Acad. Sci. USA 2015, 112, 201501598. [Google Scholar] [CrossRef] [PubMed]
- Rabaan, A.A.; Al-Ahmed, S.H.; Alsuliman, S.A.; Aldrazi, F.A.; Alfouzan, W.A.; Haque, S. The rise of pneumonic plague in Madagascar: Current plague outbreak breaks usual seasonal mould. J. Med. Microbiol. Microbiol. Soc. 2019, 68, 292–302. [Google Scholar] [CrossRef]
- Riehm, J.M.; Rahalison, L.; Scholz, H.C.; Thoma, B.; Pfeffer, M.; Razanakoto, L.M.; Al Dahouk, S.; Neubauer, H.; Tomaso, H. Detection of Yersinia pestis using real-time PCR in patients with suspected bubonic plague. Mol. Cell. Probes 2011, 25, 8–12. [Google Scholar] [CrossRef]
- Manning, J.T.; Forrester, N.; Paessler, S. Lassa virus isolates from Mali and the Ivory Coast represent an emerging fifth lineage. Front. Microbiol. 2015, 6, 1037. Available online: http://journal.frontiersin.org/article/10.3389/fmicb.2015.01037 (accessed on 16 January 2023). [CrossRef]
- World Health Organisation. Lassa Fever–Nigeria Disease Outbreak News; World Health Organisation: Geneva, Switzerland, 2019. [Google Scholar]
- Houéménou, G.; Gauthier, P.; Etougbétché, J.; Badou, S.; Dossou, H.J.; Agossou, D.; Picardeau, M.; Dobigny, G. Pathogenic Leptospira in Commensal Small Mammals from the Extensively Urbanized Coastal Benin. Urban Sci. 2019, 3, 99. [Google Scholar] [CrossRef]
- Diagne, C.; Galan, M.; Tamisier, L.; D’Ambrosio, J.; Dalecky, A.; Bâ, K.; Kane, M.; Niang, Y.; Diallo, M.; Sow, A.; et al. Ecological and sanitary impacts of bacterial communities associated to biological invasions in African commensal rodent communities. Sci. Rep. 2017, 7, 14995. [Google Scholar] [CrossRef]
- Dahmana, H.; Granjon, L.; Diagne, C.; Davoust, B.; Fenollar, F.; Mediannikov, O. Rodents as Hosts of Pathogens and Related Zoonotic Disease Risk. Pathogens 2020, 9, 202. Available online: http://www.ncbi.nlm.nih.gov/pubmed/32164206 (accessed on 25 March 2020). [CrossRef]
- Mangombi, J.B.; N’dilimabaka, N.; Lekana-Douki, J.-B.; Banga, O.; Maghendji-Nzondo, S.; Bourgarel, M.; Leroy, E.; Fenollar, F.; Mediannikov, O. First investigation of pathogenic bacteria, protozoa and viruses in rodents and shrews in context of forest-savannah-urban areas interface in the city of Franceville (Gabon). Markotter, W., Ed. PLoS ONE 2021, 16, e0248244. [Google Scholar] [CrossRef]
- Mawanda, P.; Rwego, I.; Kisakye, J.J.; Sheil, D. Rodents as potential hosts and reservoirs of parasites along the edge of a Central African forest: Bwindi impenetrable national park, South Western Uganda. Afr. Health Sci. 2020, 20, 1168. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7751523/ (accessed on 2 February 2020). [CrossRef] [PubMed]
- Suu-Ire, R.; Obodai, E.; Bel-Nono, S.O.; Ampofo, W.K.; Mazet, J.A.K.; Goldstein, T.; Johnson, C.K.; Smith, B.; Boaatema, L.; Asigbee, T.W.; et al. Surveillance for potentially zoonotic viruses in rodent and bat populations and behavioral risk in an agricultural settlement in Ghana. One Health Outlook 2022, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Duplantier, J.M.; Granjon, L.; Ba, K. Répartition biogéographique des petits rongeurs au Sénégal. J. Afr. Zool. 1997, 111, 17–26. [Google Scholar]
- Dalecky, A.; Bâ, K.; Piry, S.; Lippens, C.; Diagne, C.A.; Kane, M.; Sow, A.; Diallo, M.; Niang, Y.; Konečný, A.; et al. Range expansion of the invasive house mouse Mus musculus domesticus in Senegal, West Africa: A synthesis of trapping data over three decades, 1983–2014. Mamm. Rev. 2015, 45, 176–190. Available online: http://doi.wiley.com/10.1111/mam.12043 (accessed on 3 September 2015). [CrossRef]
- Diagne, C.; Granjon, L.; Tatard, C.; Ribas, A.; Ndiaye, A.; Kane, M.; Niang, Y.; Brouat, C. Same Invasion, Different Routes: Helminth Assemblages May Favor the Invasion Success of the House Mouse in Senegal. Front. Vet. Sci. 2021, 8, 1234. [Google Scholar] [CrossRef] [PubMed]
- Granjon, L.; Ba, K.; Diagne, C.; Ndiaye, A.; Piry, S.; Thiam, M. La communauté des petits rongeurs du Ferlo: Tendances historiques et caractéristiques du peuplement actuel. In La Grande Muraille Verte, une Réponse Africaine au Changement Climatique; Boëtsch, G., Duboz, P., Guissé, A., Sarr, P., Eds.; CNRS: Paris, France, 2019; pp. 1–19. [Google Scholar]
- Thiam, M.; Ba, K.; Duplantier, J.M. Impacts of climatic changes on small mammal communities in the Sahel (West Africa) as evidenced by owl pellet analysis|African Zoology. Afr. Zool. 2008, 43, 135–143. Available online: https://journals.co.za/doi/epdf/10.10520/EJC18091 (accessed on 4 March 2023). [CrossRef]
- Mangombi-Pambou, J.; Granjon, L.; Labarrere, C.; Kane, M.; Niang, Y.; Fournier, P.E.; Delerce, J.; Fenollar, F.; Mediannikov, O. New Genotype of Coxiella burnetii Causing Epizootic Q Fever Outbreak in Rodents, Northern Senegal-Volume 29, Number 5—May 2023-Emerging Infectious Diseases journal-CDC. Emerg. Infect. Dis. 2023, 29, 1078. Available online: https://www.nc.cdc.gov/eid/article/29/5/22-1034_article (accessed on 20 April 2023).
- Heyman, P.; Saegerman, C. Les rongeurs en tant que sentinelles dans le cadre des infections à hantavirus. Epidémiol. St. Anim. 2009, 56, 47–52. [Google Scholar]
- Sikes, R.S. Mammalogists the AC and UC of the AS of. 2016 Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. J. Mammal. 2016, 97, 663. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5909806/ (accessed on 17 January 2023). [CrossRef]
- Dahmani, M.; Davoust, B.; Tahir, D.; Raoult, D.; Fenollar, F.; Mediannikov, O. Molecular investigation and phylogeny of Anaplasmataceae species infecting domestic animals and ticks in Corsica, France. Parasites Vectors 2017, 10, 302. [Google Scholar] [CrossRef]
- Mediannikov, O.; Fenollar, F. Looking in ticks for human bacterial pathogens. Microb. Pathog. 2014, 77, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Jensen, W.A.; Fall, M.Z.; Rooney, J.; Kordick, D.L.; Breitschwerdt, E.B. Rapid identification and differentiation of Bartonella species using a single-step PCR assay. J. Clin. Microbiol. 2000, 38, 1717–1722. [Google Scholar] [CrossRef] [PubMed]
- Mediannikov, O.; Abdissa, A.; Socolovschi, C.; Diatta, G.; Trape, J.F.; Raoult, D. Detection of a New Borrelia Species in Ticks Taken from Cattle in Southwest Ethiopia. Vector-Borne Zoonotic Dis. 2013, 13, 266–269. Available online: https://www.liebertpub.com/doi/10.1089/vbz.2011.0874 (accessed on 10 February 2023). [CrossRef] [PubMed]
- Bittar, F.; Keita, M.B.; Lagier, J.C.; Peeters, M.; Delaporte, E.; Raoult, D. Gorilla gorilla gorilla gut: A potential reservoir of pathogenic bacteria as revealed using culturomics and molecular tools. Sci. Rep. 2014, 4, 7174. [Google Scholar] [CrossRef]
- Sokhna, C.; Mediannikov, O.; Fenollar, F.; Bassene, H.; Diatta, G.; Tall, A.; Trape, J.F.; Drancourt, M.; Raoult, D. Point-of-Care Laboratory of Pathogen Diagnosis in Rural Senegal. PLoS Negl. Trop. Dis. 2013, 7, e1999. [Google Scholar] [CrossRef]
- Laidoudi, Y.; Davoust, B.; Varloud, M.; Niang, E.H.A.; Fenollar, F.; Mediannikov, O. Development of a multiplex qPCR-based approach for the diagnosis of Dirofilaria immitis, D. repens and Acanthocheilonema reconditum. Parasites Vectors 2020, 13, 319.AR. Available online: https://parasitesandvectors.biomedcentral.com/articles/10.1186/s13071-020-04185-0 (accessed on 3 February 2023). [CrossRef]
- Dahmana, H.; Amanzougaghene, N.; Davoust, B.; Normand, T.; Carette, O.; Demoncheaux, J.P.; Mulot, B.; Fabrizy, B.; Scandola, P.; Chik, M.; et al. Great diversity of Piroplasmida in Equidae in Africa and Europe, including potential new species. Vet. Parasitol. Reg. Stud. Rep. 2019, 18, 100332. [Google Scholar] [CrossRef]
- Medkour, H.; Varloud, M.; Davoust, B.; Mediannikov, O. New Molecular Approach for the Detection of Kinetoplastida Parasites of Medical and Veterinary Interest. Microorganisms 2020, 8, 356. Available online: https://www.mdpi.com/2076-2607/8/3/356 (accessed on 4 March 2020). [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2007; Available online: http://www.r-project.org/ (accessed on 23 August 2018).
- Ouarti, B.; Sall, M.; Ndiaye, E.H.I.; Diatta, G.; Zan Diarra, A.; Berenger, J.M.; Sokhna, C.; Granjon, L.; Le Fur, J.; Parola, P. Pathogen Detection in Ornithodoros sonrai Ticks and Invasive House Mice Mus musculus domesticus in Senegal. Microorganisms 2022, 10, 2367. [Google Scholar] [CrossRef] [PubMed]
- Brouat, C.; Diagne, C.A.; Ismaïl, K.; Aroussi, A.; Dalecky, A.; Bâ, K.; Kane, M.; Niang, Y.; Diallo, M.; Sow, A.; et al. Seroprevalence of Toxoplasma gondii in commensal rodents sampled across Senegal, West Africa. Parasite 2018, 25, 32. [Google Scholar] [CrossRef] [PubMed]
- Galal, L.; Schares, G.; Stragier, C.; Vignoles, P.; Brouat, C.; Cuny, T.; Dubois, C.; Rohart, T.; Glodas, C.; Dardé, M.-L.; et al. Diversity of Toxoplasma gondii strains shaped by commensal communities of small mammals. Int. J. Parasitol. 2018, 49, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Diagne, C.; Charbonnel, N.; Henttonen, H.; Sironen, T.; Brouat, C. Serological Survey of Zoonotic Viruses in Invasive and Native Commensal Rodents in Senegal, West Africa. Vector-Borne Zoonotic Dis. 2017, 17, 730–733. Available online: http://online.liebertpub.com/doi/10.1089/vbz.2017.2135 (accessed on 10 April 2020). [CrossRef]
- Diagne, M.; Ndione, M.; Di Paola, N.; Fall, G.; Bedekelabou, A.; Sembène, P.; Faye, O.; Zanotto, P.M.d.A.; Sall, A.A. Usutu Virus Isolated from Rodents in Senegal. Viruses 2019, 11, 181. Available online: https://www.mdpi.com/1999-4915/11/2/181 (accessed on 30 April 2020). [CrossRef]
- André, M.R. Diversity of Anaplasma and Ehrlichia/Neoehrlichia Agents in terrestrial wild carnivores worldwide: Implications for human and domestic animal health and wildlife conservation. Front. Vet. Sci. Front. Media S.A. 2018, 5, 293. [Google Scholar] [CrossRef]
- Dumler, J.S.; Walker, D.H. Ehrlichioses and Anaplasmosis. Trop. Infect. Dis. Princ. Pathog. Pract. 2-Vol. Set CD-ROM 2006, 564–573. [Google Scholar]
- Lamine Djiba, M.; Mediannikov, O.; Mbengue, M.; Thiongane, Y.; Molez, J.F.; Talla Seck, M.; Fenollar, F.; Raoult, D.; Ndiaye, M. Survey of Anaplasmataceae bacteria in sheep from Senegal. Trop. Anim. Health Prod. 2013, 45, 1557–1561. Available online: http://blast.ncbi.nlm.nih.gov (accessed on 9 March 2023). [CrossRef]
- Angelakis, E.; Mediannikov, O.; Socolovschi, C.; Mouffok, N.; Bassene, H.; Tall, A.; Niangaly, H.; Doumbo, O.; Znazen, A.; Sarih, M.; et al. Coxiella burnetii-positive PCR in febrile patients in rural and urban Africa. Int. J. Infect. Dis. 2014, 28, 107–110. [Google Scholar] [CrossRef]
- Krügel, M.; Król, N.; Kempf, V.A.J.; Pfeffer, M.; Obiegala, A. Emerging rodent-associated Bartonella: A threat for human health? Parasites Vectors 2022, 15, 113. Available online: https://parasitesandvectors.biomedcentral.com/articles/10.1186/s13071-022-05162-5 (accessed on 27 January 2023). [CrossRef]
- Angelakis, E.; Raoult, D. Q fever. Vet. Microbiol. 2010, 140, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Kosoy, M.; Mandel, É.; Green, D.; Marston, É.; Childs, J. Prospective Studies of Bartonella of Rodents. Part I. Demographic and Temporal Patterns in Population Dynamics. Vector-Borne Infect. Dis. 2004, 4, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Diarra, A.Z.; Kone, A.K.; Niare, S.D.; Laroche, M.; Diatta, G.; Atteynine, S.A.; Coulibaly, M.; Sangare, A.K.; Kouriba, B.; Djimde, A.; et al. Molecular detection of microorganisms associated with small mammals and their ectoparasites in Mali. Am. J. Trop. Med. Hyg. 2020, 103, 2542–2551. [Google Scholar] [CrossRef] [PubMed]
- La Scola, B.; Zeaiter, Z.; Khamis, A.; Raoult, D. Gene-sequence-based criteria for species definition in bacteriology: The Bartonella paradigm. Trends Microbiol. 2003, 11, 318–321. [Google Scholar] [CrossRef]
- Mediannikov, O.; Socolovschi, C.; Bassene, H.; Diatta, G.; Ratmanov, P.; Fenollar, F.; Sokhna, C.; Raoult, D. Borrelia crocidurae infection in acutely febrile patients, Senegal. Emerg. Infect. Dis. 2014, 20, 1335–1338. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Diatta, G.; Socolovschi, C.; Mediannikov, O.; Tall, A.; Bassene, H.; Trape, J.F.; Raoult, D. Tick-Borne Relapsing Fever Borreliosis, Rural Senegal. Emerg. Infect. Dis. 2011, 17, 883. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3321757/ (accessed on 8 March 2023). [CrossRef]
- Trape, J.F.; Diatta, G.; Arnathau, C.; Bitam, I.; Sarih, M.; Belghyti, D.; Bouattour, A.; Elguero, E.; Vial, L.; Mané, Y.; et al. The epidemiology and geographic distribution of relapsing fever borreliosis in West and North Africa, with a review of the Ornithodoros erraticus complex (Acari: Ixodida). PLoS ONE 2013, 8, e78473. [Google Scholar] [CrossRef]
- Godeluck, B.; Duplantier, J.M.; Ba, K.; Trape, J.F. A Longitudinal Survey of Borrelia crocidurae Prevalence in Rodents and Insectivores in Senegal. Am. J. Trop. Med. Hyg. 1994, 50, 165–168. Available online: https://www.ajtmh.org/view/journals/tpmd/50/2/article-p165.xml (accessed on 10 April 2023). [CrossRef]
- Elbir, H.; Raoult, D.; Drancourt, M. Review article: Relapsing fever borreliae in Africa. The American Society of Tropical Medicine and Hygiene. Am. J. Trop. Med. Hyg. 2013, 89, 288–292. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3741250/ (accessed on 23 February 2023). [CrossRef]
- Diouf, F.S.; Ndiaye, E.H.I.; Hammoud, A.; DIamanka, A.; Bassene, H.; Ndiaye, M.; Mediannikov, O.; Parola, P.; Raoult, D.; Sokhna, C.; et al. Detection of Coxiella burnetii and Borrelia spp. DNA in Cutaneous Samples and in Household Dust in Rural Areas, Senegal. Vector-Borne Zoonotic Dis. 2021, 21, 659–666. Available online: https://home.liebertpub.com/vbz (accessed on 20 March 2023). [CrossRef]
- Anh, L.T.L.; Balakirev, A.E.; Van Chau, N. Investigation of multiple infections with zoonotic pathogens of rodents in northern Vietnam. J. Vector Borne Dis. 2021, 58, 47. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.C.; Velásquez, C.V.; Aquib, A.; Al-Nazal, A.; Parveen, N. Transmission Cycle of Tick-Borne Infections and Co-Infections, Animal Models and Diseases. Pathogens 2022, 11, 1309. [Google Scholar] [CrossRef] [PubMed]
- Abbate, J.L.; Galan, M.; Razzauti, M.; Sironen, T.; Voutilainen, L.; Henttonen, H.; Gasqui, P.; Cosson, J.-F.; Charbonnel, N. Pathogen community composition and co-infection patterns in a wild community of rodents. bioRxiv 2021, 2020.02.09.940494. Available online: https://www.biorxiv.org/content/10.1101/2020.02.09.940494v2 (accessed on 10 March 2023).
- Panthawong, A.; Grieco, J.P.; Ngoen-klan, R.; Chao, C.C.; Chareonviriyaphap, T. Detection of Anaplasma spp. and Bartonella spp. from wild-caught rodents and their ectoparasites in Nakhon Ratchasima Province, Thailand. J. Vector Ecol. 2020, 45, 241–253. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/jvec.12395 (accessed on 10 March 2023). [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Organism | Target Gene | Technique | Name | SEQUENCES (5′-3′) | Annealing Temperature | Amplicon | Reference |
|---|---|---|---|---|---|---|---|
| Anaplasmataceae | 23S | Broad-range qPCR | TtAna_F | TGACAGCGTACCTTTTGCAT | 55 °C | 190 bp | [24] |
| TtAna_R | GTAACAGGTTCGGTCCTCCA | ||||||
| TtAna_P | 6FAM- GGATTAGACCCGAAACCAAG | ||||||
| Broad-range conventional PCR | Ana23S-212F | ATAAGCTGCGGGGAATTGTC | 58 °C | 960 bp | [24] | ||
| Ana23S-753R | TGCAAAAGGTACGCTGTCAC (for sequencing only) | ||||||
| Ana23S-908R | GTAACAGGTTCGGTCCTCCA | ||||||
| Bartonella sp. | ITS (Intergenic 16S–23S) | Broad-range qPCR | Barto_ITS3_F | GATGCCGGGGAAGGTTTTC | 60 °C | 104 bp | [25] |
| Barto_ITS3_R | GCCTGGGAGGACTTGAACCT | ||||||
| Barto_ITS3_P | 6FAM- GCGCGCGCTTGATAAGCGTG | ||||||
| Broad-range conventional PCR | Urbarto1 | CTTCGTTTCTCTTTCTTCA | 50 °C | 733 bp | [26] | ||
| Urbarto2 | CTTCTCTTCACAATTTCAAT | ||||||
| Borrelia sp. | 16S | Broad-range qPCR | Bor_16S_3F | AGCCTTTAAAGCTTCGCTTGTAG | 60 °C | 148 bp | [27] |
| Bor_16S_3R | GCCTCCCGTAGGAGTCTGG | ||||||
| Bor_16S_3P | 6FAM- CCGGCCTGAGAGGGTGAACGG | ||||||
| Broad-range conventional PCR nested PCR | * Bor_ITS_F | TATGTTTAGTGAGGGGGGTG | 56 °C | 1034 bp | This study | ||
| * Bor_ITS_R | GATCATAGCTCAGGTGGTTAG | ||||||
| # Bor_ITSi_F | GGGGGGTGAAGTCGTAACAAG | 60 °C | 993 bp | ||||
| # Bor_ITSi_R | TCTGATAAACCTGAGGTCGGA | ||||||
| Mycoplasma sp. | ITS | Broad-range qPCR | Mycop_ITS_F | GGGAGCTGGTAATACCCAAAGT | 60 °C | 114 bp | [28] |
| Mycop_ITS_R | CCATCCCCACGTTCTCGTAG | ||||||
| Mycop_ITS_P | 6FAM-GCCTAAGGTAGGACTGGTGACTGGGG | ||||||
| Rickettsia sp. | gltA (CS) | Broad-range qPCR | RKND03_F | GTGAATGAAAGATTACACTATTTAT | 60 °C | 166 bp | [25,29] |
| RKND03_R | GTATCTTAGCAATCATTCTAATAGC | ||||||
| RKND03 P | 6-FAM-CTATTATGCTTGCGGCTGTCGGTTC | ||||||
| Pan-Filaria | 28S rRNA | Broad-range qPCR | qFil-28S-F | TTG TTT GAG ATT GCA GCC CA | 60 °C | [30] | |
| qFil-28S-R | GTT TCC ATC TCA GCG GTT TC | ||||||
| qFil-28S-P | 6FAM-CAA GTA CCG TGA GGG AAA GT | ||||||
| Piroplasma sp. | 5.8S | Broad-range qPCR | 5,8s-F5 | AYYKTYAGCGRTGGATGTC | 60 °C | 40 bp | [31] |
| 5,8s-R1 | TCGCAGRAGTCTKCAAGTC | ||||||
| 5,8s-S | 6-FAM-TTYGCTGCGTCCTTCATCGTTGT | ||||||
| Pan-Leishmania/Trypanosoma | 28S LSU | Broad-range qPCR | F Leish/Tryp | AGATCTTGGTTGGCGTAG | 60 °C | 135 bp | [32] |
| R Leish/Tryp | ATAACGTTGTGCTCAGTTTCC | ||||||
| P. Leish/Tryp | FAM-GGGAAGGATTTCGTGCCAACG | ||||||
| Pan-Kinetoplastidae | 28S LSU (24 alpha) | Broad-range qPCR | F. 24a; 5198 | AGTATTGAGCCAAAGAAGG | 60 °C | 200 bp | [32] |
| R. 24a; 5412 | TTGTCACGACTTCAGGTTCTAT | ||||||
| P. 24a; 5345 | FAM- TAGGAAGACCGATAGCGAACAAGTAG |
| Rodent Species | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Microorganism detected (qPCR-positive individual number) | A. niloticus N = 29 | D. braueri N = 3 | T. pygargus N = 8 | G. nancillus N = 9 | J. jaculus N = 4 | # G. nigeriae N = 71 | X. erythropus N = 1 | Total N = 125 (prevalence%) | ||
| Prevalence of microorganisms detected | Anaplasmataceae (25) | 2/29 (6.9%) | 0 | 2/8 (25%) | 0 | 0 | 21/71 (29.6%) | 0 | 25/125 (20%) | |
| Bartonella spp. (30) | 3/29 (10.3%) | 1/3 (33.3%) | 4/8 (50%) | 0 | 0 | 21/71 (29.6%) | 1/1 (100%) | 30/125 (24%) | ||
| Borrelia spp. (13) | 5/29 (17.2%) | 1/3 (33.3%) | 3/8 (37.5%) | 1/9 (11.1%) | 0 | 3/71 (4.22%) | 0 | 13/125 (10.4%) | ||
| C. burnetii (28) | 0 | 1/3 (33.3%) | 3/8 (37.5%) | 3/9 (33.3%) | 1 (25%) | 20/71 (28.2%) | 0 | 28/125 (22.4) | ||
| Piroplasmida (3) | 0 | 0 | 3/8 (37.5%) | 0 | 0 | 0 | 0 | 3/125 (2.4%) | ||
| Infection type | One infection (43) | Anaplasma spp. | 1 | 0 | 0 | 0 | 0 | 7 | 0 | 8 |
| Bartonella spp. | 2 | 1 | 0 | 0 | 0 | 11 | 1 | 15 | ||
| Borrelia spp. | 5 | 1 | 1 | 1 | 0 | 1 | 0 | 9 | ||
| C. burnetii | 0 | 0 | 1 | 3 | 1 | 6 | 0 | 11 | ||
| Double infections (16) | Anaplasmataceae/Bartonella | 1 | 0 | 1 | 0 | 0 | 2 | 0 | 4 | |
| Anaplasmataceae/Borrelia | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | ||
| Anaplasmataceae/Coxiella | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 6 | ||
| Borrelia/Coxiella | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | ||
| Bartonella/Coxiella | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 2 | ||
| Bartonella/Borrelia | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | ||
| Bartonella/Piroplasmida | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | ||
| Triple infections (8) | Anaplasmataceae/Bartonella/Coxiella | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 5 | |
| Anaplasmataceae/Coxiella/Piroplasmida | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | ||
| Bartonella/Borrelia/Coxiella | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | ||
| Bartonella/Coxiella/Piroplasmida | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | ||
| Total infected rodents | 9 (31.03%) | 3 (100%) | 7 (87.5%) | 4 (44.4%) | 1 (25%) | 42 (59.1%) | 1 (100%) | 67 | ||
| Label | Variable | Infection | Total | Test | |
|---|---|---|---|---|---|
| 0 | 1 | ||||
| Age | Adults | 37 (39.8%) | 56 (60.2%) | 93 (74.4%) | p-value: 0.01 |
| Juveniles | 21 (65.6%) | 11 (34.4%) | 32 (25.6%) | ||
| Total | 58 (46.4%) | 67 (53.6%) | 125 (100.0%) | ||
| Sex | Females | 32 (51.6%) | 30 (48.4%) | 62 (49.6%) | p-value: 1 |
| Males | 26 (41.3%) | 37 (58.7%) | 63 (50.4%) | ||
| Total | 58 (46.4%) | 67 (53.6%) | 125 (100.0%) | ||
| Status | Expanding species | 29 (40.8%) | 42 (59.2%) | 71 (56.8%) | p-value: 0.75 |
| Native species | 29 (53.7%) | 25 (46.3%) | 54 (43.2%) | ||
| Total | 58 (46.4%) | 67 (53.6%) | 125 (100.0%) | ||
| Season | Dry season | 38 (42.2%) | 52 (57.8%) | 90 (72.0%) | p-value: 0.12 |
| Rainy season | 20 (57.1%) | 15 (42.9%) | 35 (28.0%) | ||
| Total | 58 (46.4%) | 67 (53.6%) | 125 (100.0%) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mangombi-Pambou, J.B.; Granjon, L.; Flirden, F.; Kane, M.; Niang, Y.; Davoust, B.; Fenollar, F.; Mediannikov, O. Molecular Survey of Rodent-Borne Infectious Agents in the Ferlo Region, Senegal. Genes 2023, 14, 1107. https://doi.org/10.3390/genes14051107
Mangombi-Pambou JB, Granjon L, Flirden F, Kane M, Niang Y, Davoust B, Fenollar F, Mediannikov O. Molecular Survey of Rodent-Borne Infectious Agents in the Ferlo Region, Senegal. Genes. 2023; 14(5):1107. https://doi.org/10.3390/genes14051107
Chicago/Turabian StyleMangombi-Pambou, Joa Braïthe, Laurent Granjon, Fabien Flirden, Mamadou Kane, Youssoupha Niang, Bernard Davoust, Florence Fenollar, and Oleg Mediannikov. 2023. "Molecular Survey of Rodent-Borne Infectious Agents in the Ferlo Region, Senegal" Genes 14, no. 5: 1107. https://doi.org/10.3390/genes14051107