

The Operon as a Conundrum of Gene Dynamics and Biochemical Constraints: What We Have Learned from Histidine Biosynthesis

Abstract
:1. The “Operon Model”: Story of an Idea
2. Definition of Operon
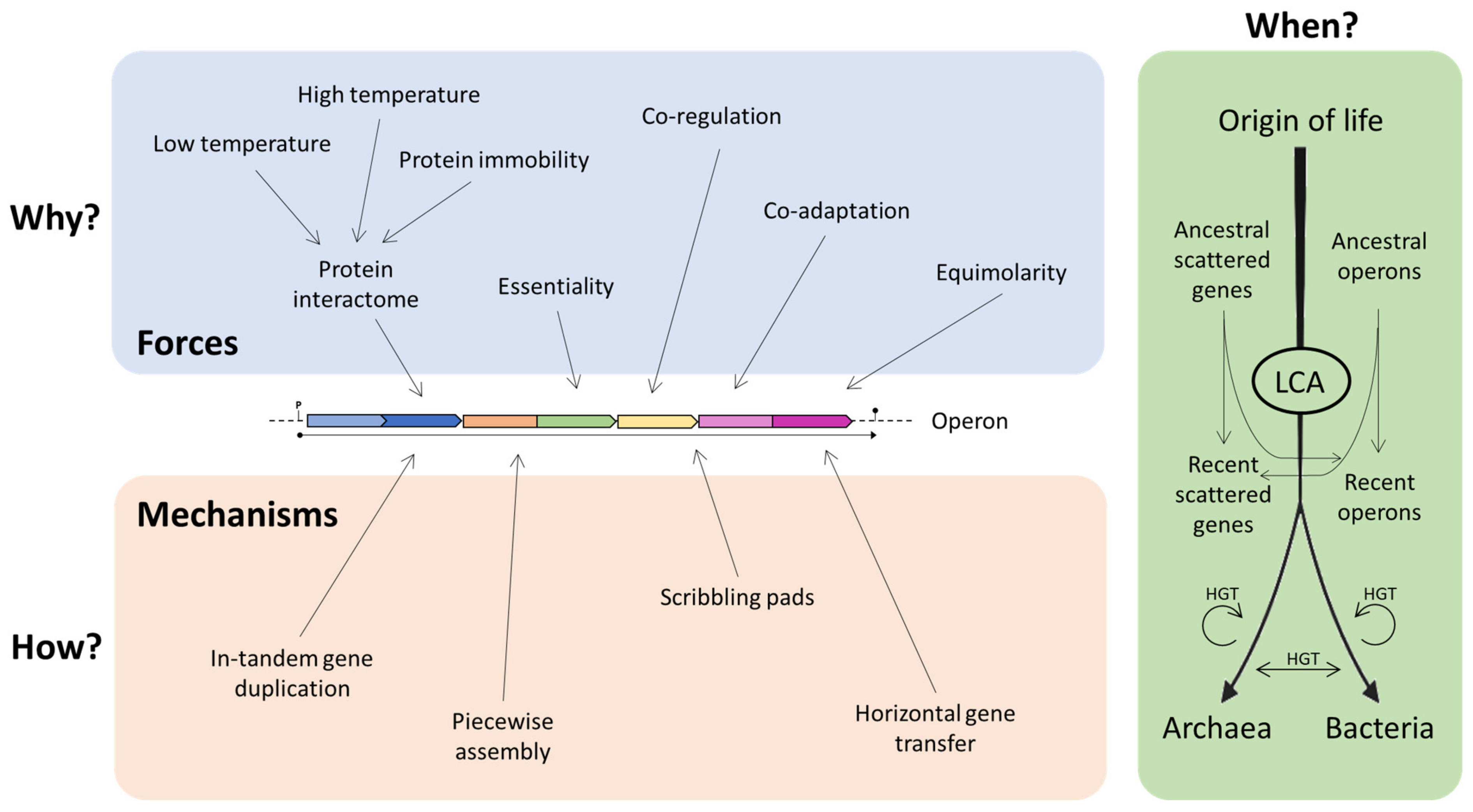
3. Hypotheses on the Origin and Evolution of Operons
3.1. Why?
- i.
- ii.
- The “co-regulation model” predicts that genes should be found in operons when their co-regulation would be the most advantageous; indeed, gene clusters promote coordinated expression and regulation ([43], and references therein).
- iii.
- iv.
- In the “selfish operon model”, proposed by Lawrence and Roth in 1996 [37], horizontal gene transfer (HGT) guarantees the spread of functionally related genes organized in gene clusters. The physical closeness of genes does not provide any selective advantages to the single organism but enhances the fitness of the entire gene cluster, as when genes are physically close the probability of a combined transfer of genes increases.
- v.
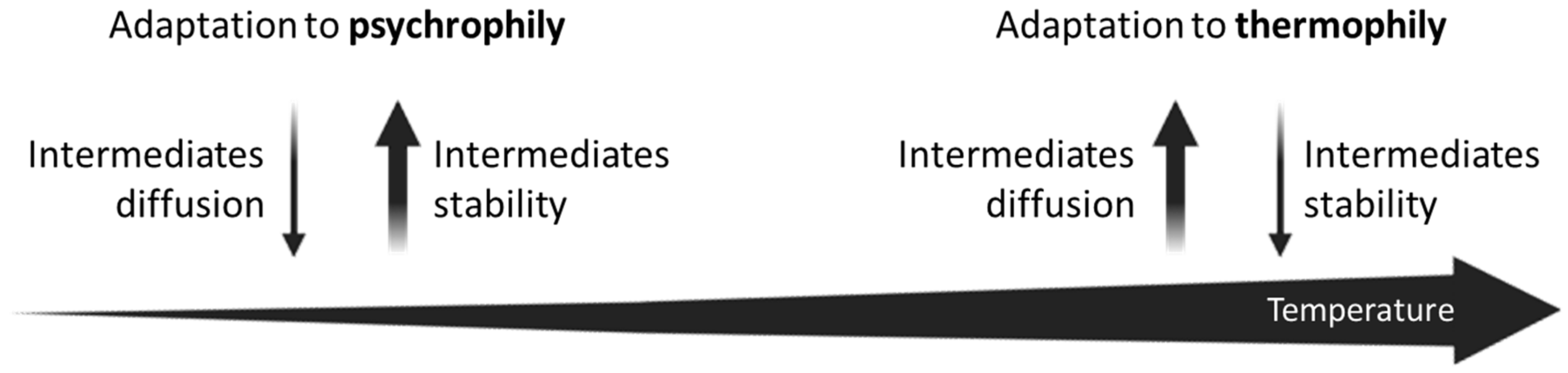
- Glansdorff [38] proposed that the “adaptation to thermophily” performed a crucial role in the formation of operons. Co-translation of proteins that are functionally related would have facilitated the formation of multienzyme complexes able to channel thermolabile substrates and the mutual stabilization of intrinsically thermolabile proteins.
- vi.
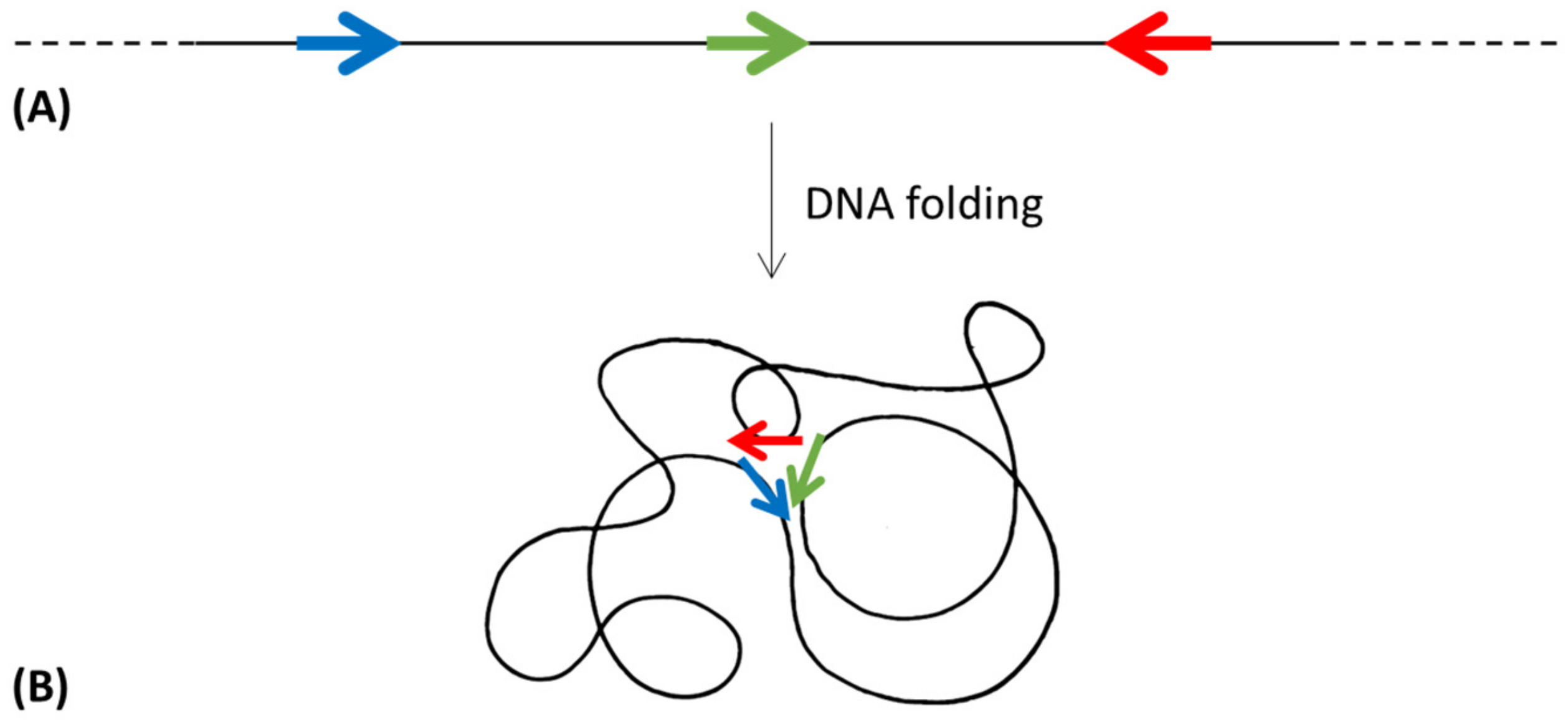
- In accordance with the previous model, the “protein immobility model” (PIM) [39] proposes that gene clustering was pushed by a thermodynamic advantage obtained by the physical closeness of newly translated proteins; in this way the product of one enzyme could easily find its target.
- vii.
- More recently, Nguyen and coworkers [40] developed and used a maximum parsimony algorithm to recreate ancestral operon states. They suggested that two forces, i.e., “the essentiality (the trait of being essential to life) and the formation of a protein complex are two drivers for gene block conservation”. Their idea relies on the analysis of some Bacillus subtilis and E. coli operons. They also suggested that (i) some operons can quickly and independently evolve in various branches in their taxonomic groups, suggesting that selective pressure plays a key role in the evolution of bacterial operons; (ii) other operons are highly conserved, since their evolution predates the LCA of the investigated clades, (iii) some ancestral operons can be described as intermediate functional structures, and (iv) some operon conservation is occasional, suggesting an involvement of horizontal gene transfer.
3.2. How?
- i.
- The “natal model” proposes that operons arose in situ by in-tandem gene divergence and duplication [43], corresponding to the Horowitz “retrograde hypothesis” on the origin and evolution of metabolic routes [41]. According to Horowitz, in the primordial heterotrophic bacteria, various biosynthetic capacities were acquired in a stepwise and sequential enzyme development following a reverse order compared to that of the extant pathways [46]. However, the Horowitz hypothesis has been shown to be relevant in very few cases ([44] and references therein).
- ii.
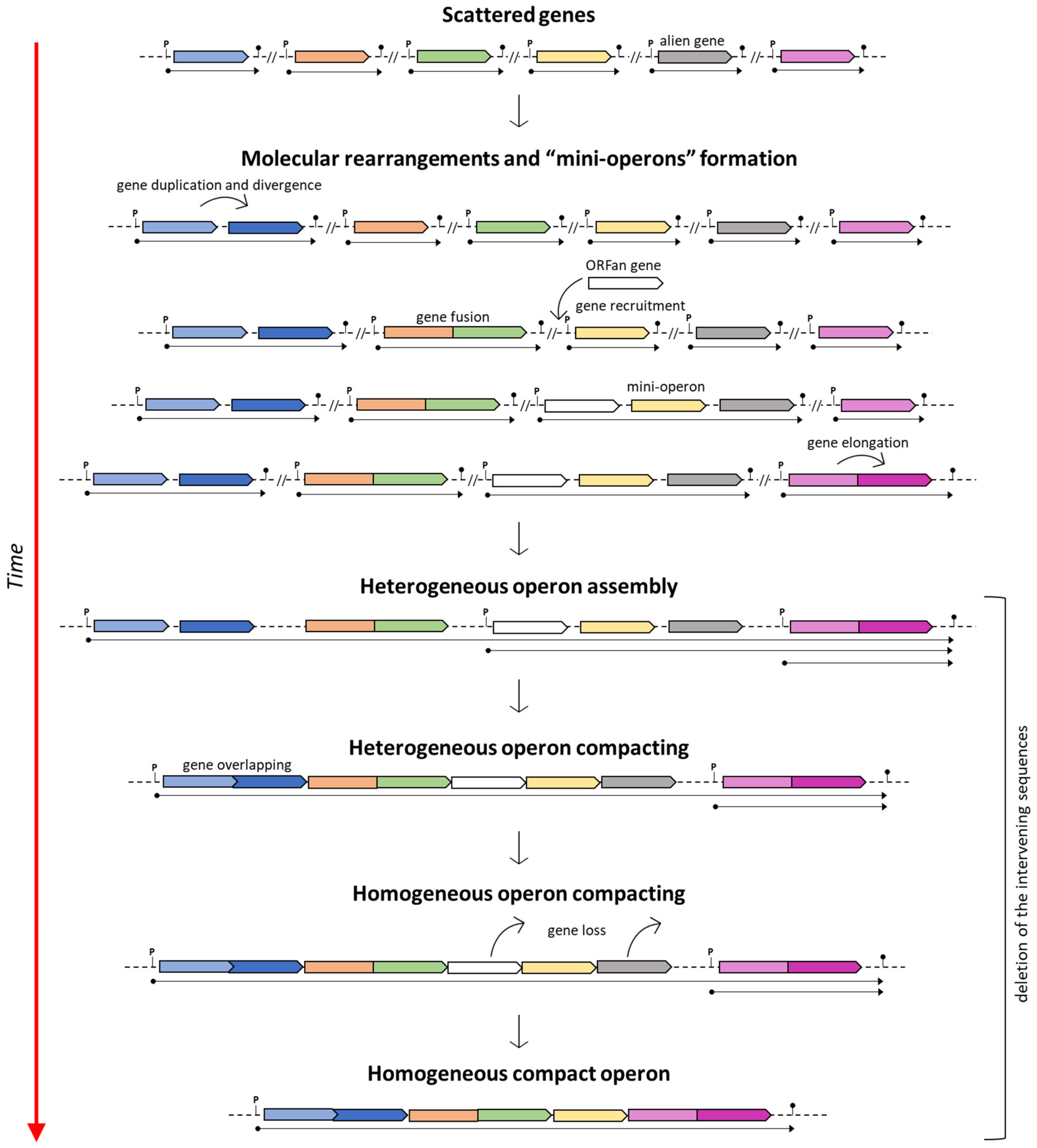
- Fani and coworkers [13] proposed a “piecewise model” for the origin and evolution of the histidine operon in proteobacteria. According to this model, in the ancestor of proteobacteria his genes were initially scattered, coding for monofunctional enzymes; then, they underwent a stepwise compacting process that reached its climax in some γ-proteobacteria.
- iii.
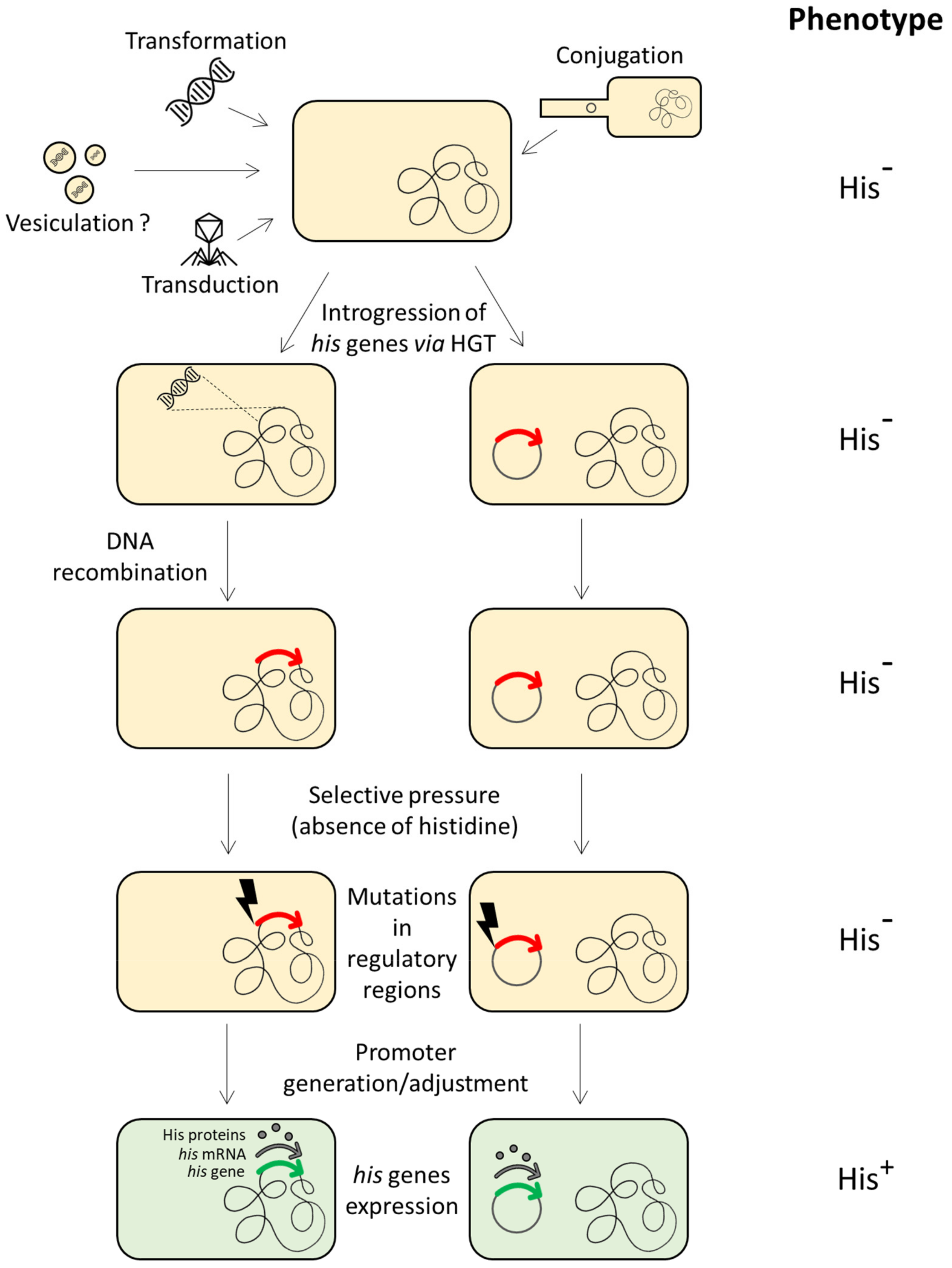
- In the “scribbling pad model”, Norris and Merieau [42] proposed that operon construction could be due to plasmids and integrative conjugative elements. According to this hypothesis, (i) a gene is copied onto a plasmid, (ii) this copy is mutated, (iii) other genes encoding related functions are duplicated and mutated on the plasmid, (iv) these genes are rearranged on the plasmid forming operons, and (v) the resulting operons are transferred back to the chromosome and/or to other bacteria.
3.3. When?
- i.
- in the genome of the LCA, genes were clustered in operons; this arrangement was then entirely or partly torn down during evolution in some descendants’ branches;
- ii.
- LCA genes were (partially) scattered throughout the genome and the construction of clusters and/or operons occurred in some of the descendants.
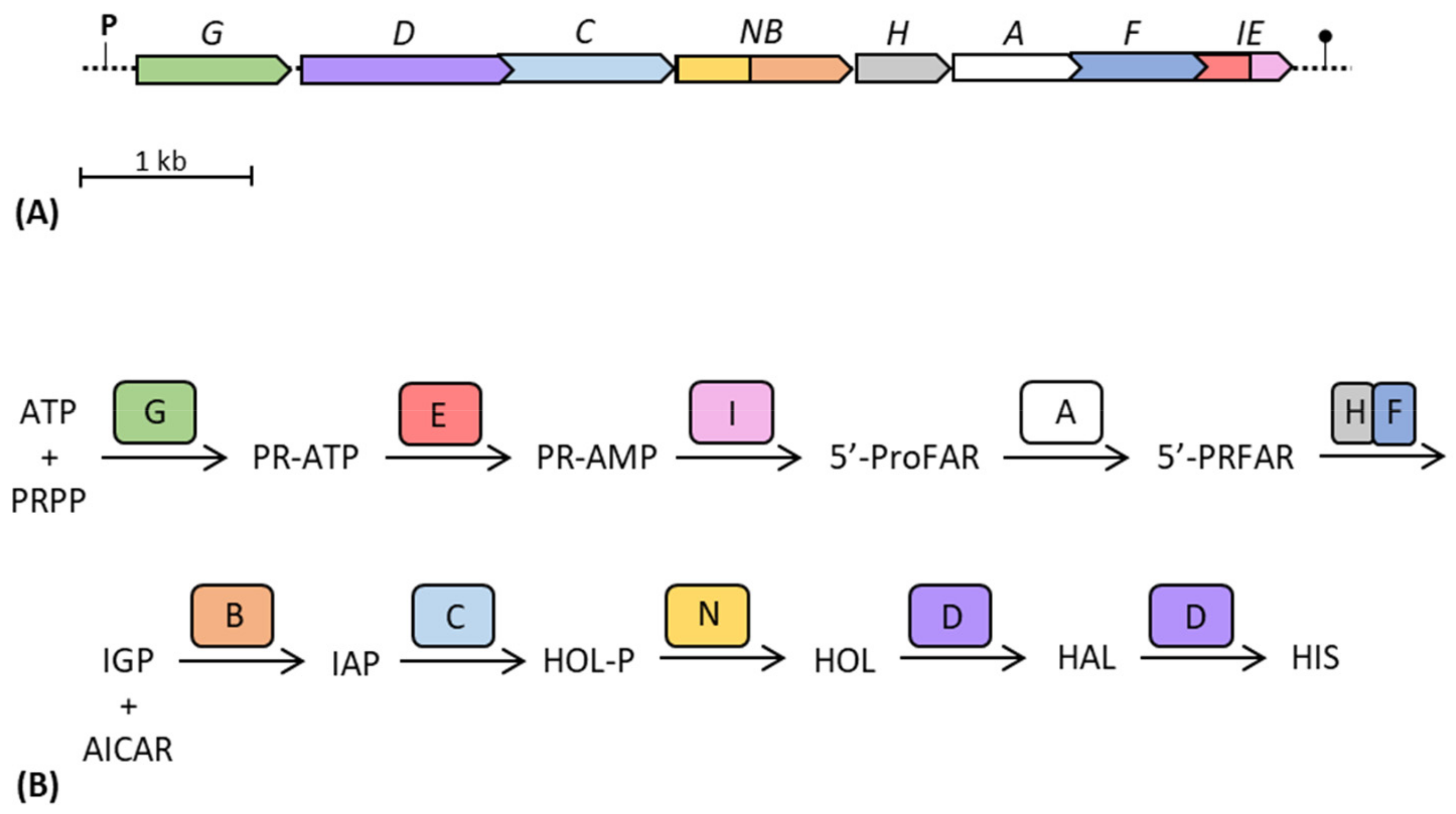
4. The Histidine Biosynthetic Pathway
5. The Histidine Operon as a Model for the Study of Operon Origin and Evolution
5.1. The Piecewise Model
5.2. The Selfish Operon Model
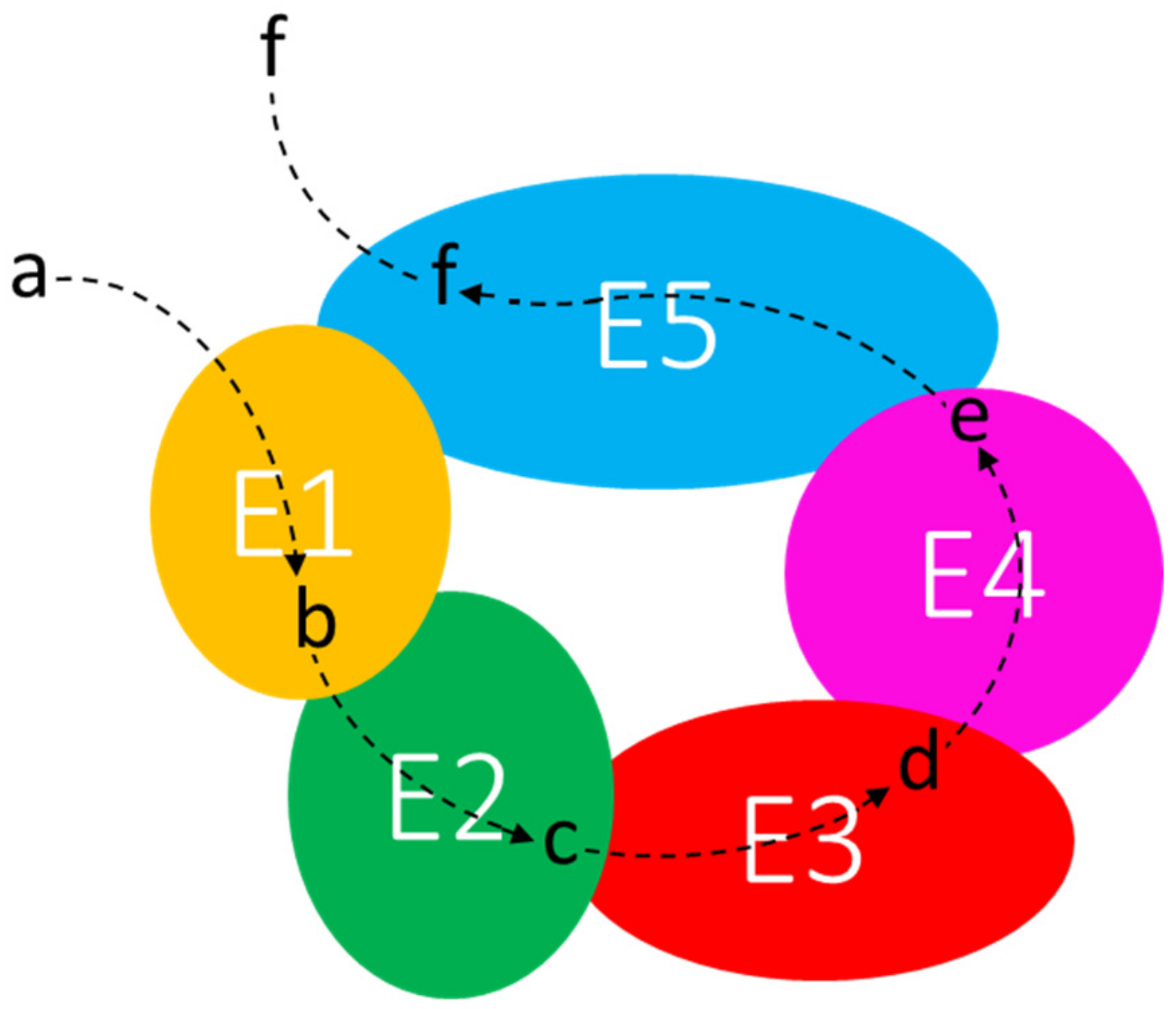
5.3. The Interactome Model
5.4. The Adaptation to Extreme Temperatures
5.5. Other Hypotheses
- i.
- The organization and the origin of the gene pair hisA-hisF supports the natal model [61,113]. Horowitz [114] suggested that the evolution of all genes constituting an operon (and thus, all of the His biosynthetic genes) results from the duplication of a common ancestor gene; however, the analysis of the nucleotide sequence of most of the E. coli and S. enterica his genes did not reveal any consistent sequence homology between different genes [113]. Despite this, hisA and hisF originated from a gene duplication event of a common ancestral gene. They both code for (β/α)8-barrels and the comparative analysis of the amino acid sequences of HisA and HisF suggested that their respective genes are paralogous and originated from a common ancestor gene through duplication and following evolutionary divergence [113]. For these reasons, HisA and HisF can be viewed as models of retrograde evolution of enzymes in a biosynthetic pathway.
- ii.
- The proximity of hisH and hisF in the his operon/core might be in agreement with the molarity model; indeed, the two enzymes must interact in a 1:1 ratio to obtain the functioning imidazole glycerol phosphate (IGP) synthase, the heterodimeric holoenzyme that links His biosynthesis to both nitrogen metabolism and de novo synthesis of purines [112]. The possibility that also other (if not all) histidine biosynthetic enzymes could physically interact forming a metabolon (see Section 5.3) further supports this evolutionary model.
- iii.
- The existence of multiple sophisticated regulatory systems controlling his gene expression supports the co-regulation model. In bacteria and lower eukaryotes, the histidine pathway is controlled by regulatory mechanisms working at the levels of both gene expression and enzyme regulation [65]. One post-translational regulatory system is the histidine-mediated feedback inhibition of HisG [115,116]. Then, at least in E. coli and S. enterica, histidine biosynthesis can be also regulated at the levels of (i) transcription initiation [60,117,118] thanks to the presence of a primary promoter and two internal promoters, (ii) transcription elongation [60,119,120] through attenuation mechanisms, (iii) transcription termination at the level of cryptic intra-cistronic Rho-dependent terminators [60,121,122,123], and (iv) post-transcription mRNA processing and decay [60,124]. The translational coupling due to the substantial overlap existing between his genes and the presence of three genes encoding bifunctional enzymes (hisD, hisNB and hisIE) also reinforce the necessity for coregulation.
6. Histidine Genes Order and Biochemical Constraints for Operon Assembly
7. When Genes Are Not Organized in Operons
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- English, M.A.; Gayet, R.V.; Collins, J.J. Designing Biological Circuits: Synthetic Biology within the Operon Model and beyond. Annu. Rev. Biochem. 2021, 90, 221–244. [Google Scholar] [CrossRef]
- Yaniv, M. The 50th anniversary of the publication of the operon theory in the journal of molecular biology: Past, present and future. J. Mol. Biol. 2011, 409, 1–6. [Google Scholar] [CrossRef]
- Morange, M. The scientific legacy of Jacques Monod. Res. Microbiol. 2010, 161, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F. The birth of the operon. Science 2011, 332, 767. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, A. Jacques Monod, 1910-1976: His life, his work and his commitments. Res. Microbiol. 2010, 161, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Monod, J. Genetic regulatory mechanisms in the synthesis of proteins. J. Mol. Biol. 1961, 3, 318–356. [Google Scholar] [CrossRef]
- Beckwith, J. The operon as paradigm: Normal science and the beginning of biological complexity. J. Mol. Biol. 2011, 409, 7–13. [Google Scholar] [CrossRef]
- Reznikoff, W.S. The operon revisited. Annu. Rev. Genet. 1972, 6, 133–156. [Google Scholar] [CrossRef]
- Gayon, J.; Gros, F.; Morange, M. Jacques Monod: Fifty years after—Foreword. Comptes Rendus Biol. 2015, 338, 369–371. [Google Scholar] [CrossRef]
- Tajbakhsh, S.; Cavalli, G.; Richet, E. Integrated Gene Regulatory Circuits: Celebrating the 50th Anniversary of the Operon Model. Mol. Cell 2011, 43, 505–514. [Google Scholar] [CrossRef]
- Jayaraman, R. Jacques Monod and the advent of the age of operons. Resonance 2010, 15, 1084–1096. [Google Scholar] [CrossRef]
- Kirschner, M.W.; Shapiro, L.; McAdams, H.; Almouzni, G.; Sharp, P.A.; Young, R.A.; Alon, U. Fifty years after Jacob and Monod: What are the unanswered questions in molecular biology? Mol. Cell 2011, 42, 403–404. [Google Scholar] [CrossRef] [PubMed]
- Fani, R.; Brilli, M.; Liò, P. The origin and evolution of operons: The piecewise building of the proteobacterial histidine operon. J. Mol. Evol. 2005, 60, 378–390. [Google Scholar] [CrossRef]
- Hurst, L.D.; Pál, C.; Lercher, M.J. The evolutionary dynamics of eukaryotic gene order. Nat. Rev. Genet. 2004, 5, 299–310. [Google Scholar] [CrossRef]
- Boycheva, S.; Daviet, L.; Wolfender, J.L.; Fitzpatrick, T.B. The rise of operon-like gene clusters in plants. Trends Plant Sci. 2014, 19, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Omelchenko, M.V.; Makarova, K.S.; Wolf, Y.I.; Rogozin, I.B.; Koonin, E.V. Evolution of mosaic operons by horizontal gene transfer and gene displacement in situ. Genome Biol. 2003, 4, R55. [Google Scholar] [CrossRef]
- Rocha, E.P.C. The organization of the bacterial genome. Annu. Rev. Genet. 2008, 42, 211–233. [Google Scholar] [CrossRef]
- Price, M.N.; Arkin, A.P.; Alm, E.J. The life-cycle of operons. PLoS Genet. 2006, 2, e96. [Google Scholar] [CrossRef]
- Okuda, S.; Kawashima, S.; Kobayashi, K.; Ogasawara, N.; Kanehisa, M.; Goto, S. Characterization of relationships between transcriptional units and operon structures in Bacillus subtilis and Escherichia coli. BMC Genom. 2007, 8, 48. [Google Scholar] [CrossRef]
- Zheng, Y.; Szustakowski, J.D.; Fortnow, L.; Roberts, R.J.; Kasif, S. Computational identification of operons in microbial genomes. Genome Res. 2002, 12, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- De Daruvar, A.; Collado-Vides, J.; Valencia, A. Analysis of the cellular functions of Escherichia coli operons and their conservation in Bacillus subtilis. J. Mol. Evol. 2002, 55, 211–221. [Google Scholar] [CrossRef]
- Lupski, J.R.; Nigel Godson, G. The rpsU-dnaG-rpoD macromolecular synthesis operon of E. coli. Cell 1984, 39, 251–252. [Google Scholar] [CrossRef] [PubMed]
- Touchon, M.; Rocha, E.P.C. Coevolution of the organization and structure of prokaryotic genomes. Cold Spring Harb. Perspect. Biol. 2016, 8, a018168. [Google Scholar] [CrossRef]
- Itoh, T.; Takemoto, K.; Mori, H.; Gojobort, T. Evolutionary instability of operon structures disclosed by sequence comparisons of complete microbial genomes. Mol. Biol. Evol. 1999, 16, 332–346. [Google Scholar] [CrossRef] [PubMed]
- Wolf, Y.I.; Rogozin, I.B.; Kondrashov, A.S.; Koonin, E.V. Genome alignment, evolution of prokaryotic genome organization, and prediction of gene function using genomic context. Genome Res. 2001, 11, 356–372. [Google Scholar] [CrossRef]
- Fondi, M.; Emiliani, G.; Fani, R. Origin and evolution of operons and metabolic pathways. Res. Microbiol. 2009, 160, 502–512. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.N.; Lee, Y.; Hussein, R. Fundamental relationship between operon organization and gene expression. Proc. Natl. Acad. Sci. USA 2011, 108, 10626–10631. [Google Scholar] [CrossRef] [PubMed]
- Spieth, J.; Brooke, G.; Kuersten, S.; Lea, K.; Blumenthal, T. Operons in C. elegans: Polycistronic mRNA precursors are processed by trans-splicing of SL2 to downstream coding regions. Cell 1993, 73, 521–532. [Google Scholar] [CrossRef]
- Cutter, A.D.; Agrawal, A.F. The evolutionary dynamics of operon distributions in eukaryote genomes. Genetics 2010, 185, 685–693. [Google Scholar] [CrossRef]
- Kominek, J.; Doering, D.T.; Opulente, D.A.; Shen, X.X.; Zhou, X.; DeVirgilio, J.; Hulfachor, A.B.; Groenewald, M.; Mcgee, M.A.; Karlen, S.D.; et al. Eukaryotic Acquisition of a Bacterial Operon. Cell 2019, 176, 1356–1366. [Google Scholar] [CrossRef]
- Osbourn, A.E.; Field, B. Operons. Cell. Mol. Life Sci. 2009, 66, 3755–3775. [Google Scholar] [CrossRef]
- Brouwer, R.W.W.; Kuipers, O.P.; Van Hijum, S.A.F.T. The relative value of operon predictions. Brief. Bioinform. 2008, 9, 367–375. [Google Scholar] [CrossRef]
- Tjaden, B. A computational system for identifying operons based on RNA-seq data. Methods 2020, 176, 62–70. [Google Scholar] [CrossRef]
- Tamames, J. Evolution of gene order conservation in prokaryotes. Genome Biol. 2001, 2, 1–11. [Google Scholar] [CrossRef]
- Fisher, R.A. The Genetical Theory of Natural Selection: A Complete Variorum Edition; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Jacob, F.; Perrin, D.; Sánchez, C.; Monod, J. The operon: A group of genes whose expression is coordinated by an operator. Compte Rendu L’academie Des Sci. 1960, 250, 1727–1729. [Google Scholar]
- Lawrence, J.G.; Roth, J.R. Selfish operons: Horizontal transfer may drive the evolution of gene clusters. Genetics 1996, 143, 1843–1860. [Google Scholar] [CrossRef] [PubMed]
- Glansdorff, N. On the origin of operons and their possible role in evolution toward thermophily. J. Mol. Evol. 1999, 49, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Svetic, R.E.; MacCluer, C.R.; Buckley, C.O.; Smythe, K.L.; Jackson, J.H. A metabolic force for gene clustering. Bull. Math. Biol. 2004, 66, 559–581. [Google Scholar] [CrossRef]
- Nguyen, H.N.; Jain, A.; Eulenstein, O.; Friedberg, I. Tracing the ancestry of operons in bacteria. Bioinformatics 2019, 35, 2998–3004. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, N.H. On the Evolution of Biochemical Syntheses. Proc. Natl. Acad. Sci. USA 1945, 31, 153–157. [Google Scholar] [CrossRef]
- Norris, V.; Merieau, A. Plasmids as scribbling pads for operon formation and propagation. Res. Microbiol. 2013, 164, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.G. Selfish operons and speciation by gene transfer. Trends Microbiol. 1997, 5, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Fani, R.; Fondi, M. Origin and evolution of metabolic pathways. Phys. Life Rev. 2009, 6, 23–52. [Google Scholar] [CrossRef]
- Swain, P.S. Efficient attenuation of stochasticity in gene expression through post-transcriptional control. J. Mol. Biol. 2004, 344, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Lazcano, A.; Díaz-Villagómez, E.; Mills, T.; Oró, J. On the levels of enzymatic substrate specificity: Implications for the early evolution of metabolic pathways. Adv. Space Res. 1995, 15, 345–356. [Google Scholar] [CrossRef]
- Woese, C. The universal ancestor. Proc. Natl. Acad. Sci. USA 1998, 95, 6854–6859. [Google Scholar] [CrossRef] [PubMed]
- Mushegian, A.R.; Koonin, E.V. A minimal gene set for cellular life derived by comparison of complete bacterial genomes. Proc. Natl. Acad. Sci. USA 1996, 93, 10268–10273. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Mori, H.; Itoh, T.; Gojobori, T. Genome plasticity as a paradigm of eubacteria evolution. J. Mol. Evol. 1997, 44, S57–S64. [Google Scholar] [CrossRef] [PubMed]
- Kolstø, A.B. Dynamic bacterial genome organization. Mol. Microbiol. 1997, 24, 241–248. [Google Scholar] [CrossRef]
- Huynen, M.A.; Bork, P. Measuring genome evolution. Proc. Natl. Acad. Sci. USA 1998, 95, 5849–5856. [Google Scholar] [CrossRef]
- Dandekar, T.; Snel, B.; Huynen, M.; Bork, P. Conservation of gene order: A fingerprint of proteins that physically interact. Trends Biochem. Sci. 1998, 23, 324–328. [Google Scholar] [CrossRef]
- Liao, S.M.; Du, Q.S.; Meng, J.Z.; Pang, Z.W.; Huang, R.B. The multiple roles of histidine in protein interactions. Chem. Cent. J. 2013, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Ingle, R.A. Histidine biosynthesis. Arab. Book/Am. Soc. Plant Biol. 2011, 9, e0141. [Google Scholar] [CrossRef]
- Haas, F.; Mitchell, M.B.; Ames, B.N.; Mitchell, H.K. A series of histidineless mutants of Neurospora crassa. Genetics 1952, 37, 217. [Google Scholar] [CrossRef]
- Fani, R.; Brilli, M.; Fondi, M.; Lió, P. The role of gene fusions in the evolution of metabolic pathways: The histidine biosynthesis case. BMC Evol. Biol. 2007, 7, S4. [Google Scholar] [CrossRef]
- Chioccioli, S.; Del Duca, S.; Vassallo, A.; Castronovo, L.M.; Fani, R. Exploring the role of the histidine biosynthetic hisF gene in cellular metabolism and in the evolution of (ancestral) genes: From LUCA to the extant (micro)organisms. Microbiol. Res. 2020, 240, 126555. [Google Scholar] [CrossRef]
- Del Duca, S.; Chioccioli, S.; Vassallo, A.; Castronovo, L.M.; Fani, R. The role of gene elongation in the evolution of histidine biosynthetic genes. Microorganisms 2020, 8, 732. [Google Scholar] [CrossRef] [PubMed]
- Kulis-Horn, R.K.; Persicke, M.; Kalinowski, J. Corynebacterium glutamicum ATP-phosphoribosyl transferases suitable for l-histidine production—Strategies for the elimination of feedback inhibition. J. Biotechnol. 2015, 206, 26–37. [Google Scholar] [CrossRef]
- Alifano, P.; Fani, R.; Liò, P.; Lazcano, A.; Bazzicalupo, M.; Carlomagno, M.S.; Bruni, C.B. Histidine biosynthetic pathway and genes: Structure, regulation, and evolution. Microbiol. Rev. 1996, 60, 44–69. [Google Scholar] [CrossRef] [PubMed]
- Fani, R.; Liò, P.; Lazcano, A. Molecular evolution of the histidine biosynthetic pathway. J. Mol. Evol. 1995, 41, 760–774. [Google Scholar] [CrossRef]
- Shen, C.; Mills, T.; Oró, J. Prebiotic synthesis of histidyl-histidine. J. Mol. Evol. 1990, 31, 175–179. [Google Scholar] [CrossRef]
- Fani, R.; Mori, E.; Tamburini, E.; Lazcano, A. Evolution of the structure and chromosomal distribution of histidine biosynthetic genes. Orig. Life Evol. Biosph. 1998, 28, 555–570. [Google Scholar] [CrossRef] [PubMed]
- Del Duca, S.; Riccardi, C.; Vassallo, A.; Fontana, G.; Castronovo, L.M.; Chioccioli, S.; Fani, R. The histidine biosynthetic genes in the superphylum Bacteroidota-Rhodothermota-Balneolota-Chlorobiota: Insights into the evolution of gene structure and organization. Microorganisms 2021, 9, 1439. [Google Scholar] [CrossRef] [PubMed]
- Stepansky, A.; Leustek, T. Histidine biosynthesis in plants. Amino Acids 2006, 30, 127–142. [Google Scholar] [CrossRef]
- Martin, R.G. The One Operon-One Messenger Theory of Transcription. Cold Spring Harb. Symp. Quant. Biol. 1963, 28, 357–361. [Google Scholar] [CrossRef]
- Hartman, P.E. Linked loci in the control of consecutive steps in the primary pathway of histidine synthesis in Salmonella typhimurium. Genet. Stud. Bact. 1956, 612, 35–61. [Google Scholar]
- Ames, B.N.; Garry, B.; Herzenberg, L.A. The genetic control of the enzymes of histidine biosynthesis in Salmonella typhimurium. J. Gen. Microbiol. 1960, 22, 369–378. [Google Scholar] [CrossRef]
- Ames, B.N.; Hartman, P.E. The Histidine Operon. Cold Spring Harb. Symp. Quant. Biol. 1963, 28, 349–356. [Google Scholar] [CrossRef]
- Jacob, F.; Monod, J. On the Regulation of Gene Activity. Cold Spring Harb. Symp. Quant. Biol. 1961, 26, 193–211. [Google Scholar] [CrossRef]
- Roth, J.R.; Ames, B.N. Histidine regulatory mutants in Salmonella typhimurium: II. Histidine regulatory mutants having altered histidyl-tRNA synthetase. J. Mol. Biol. 1966, 22, 325–334. [Google Scholar] [CrossRef]
- Silbert, D.F.; Fink, G.R.; Ames, B.N. Histidine regulatory mutants in Salmonella typhimurium: III. A class of regulatory mutants deficient in tRNA for histidine. J. Mol. Biol. 1966, 22, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Fink, G.R.; Kłopotowski, T.; Ames, B.N. Histidine regulatory mutants in Salmonella typhimurium. IV. A positive selection for polar histidine-requiring mutants from histidine operator constitutive mutants. J. Mol. Biol. 1967, 30, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Antón, D.N. Histidine regulatory mutants in Salmonella typhimurium. V. Two new classes of histidine regulatory mutants. J. Mol. Biol. 1968, 33, 533–546. [Google Scholar] [CrossRef]
- Kasai, T. Regulation of the expression of the histidine operon in Salmonella typhimurium. Nature 1974, 249, 523–527. [Google Scholar] [CrossRef]
- Yanofsky, C. Attenuation in the control of expression of bacterial operons. Nature 1981, 289, 751–758. [Google Scholar] [CrossRef]
- Jackson, E.N.; Yaetofsky, C. The region between the operator and first structural gene of the tryptophan operon of Escherichia coli may have a regulatory function. J. Mol. Biol. 1973, 76, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Brilli, M.; Fani, R. The origin and evolution of eucaryal HIS7 genes: From metabolon to bifunctional proteins? Gene 2004, 339, 149–160. [Google Scholar] [CrossRef]
- Limauro, D.; Avitabile, A.; Cappellano, C.; Puglia, A.M.; Bruni, C.B. Cloning and characterization of the histidine biosynthetic gene cluster of Streptomyces coelicolor A3(2). Gene 1990, 90, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Henner, D.J.; Band, L.; Flaggs, G.; Chen, E. The organization and nucleotide sequence of the Bacillus subtilis hisH, tyrA and aroE genes. Gene 1986, 49, 147–152. [Google Scholar] [CrossRef]
- Fani, R.; Alifano, P.; Allotta, G.; Bazzicalupo, M.; Carlomagno, M.S.; Gallori, E.; Rivellini, F.; Polsinelli, M. The histidine operon of Azospirillum brasilense: Organization, nucleotide sequence and functional analysis. Res. Microbiol. 1993, 144, 187–200. [Google Scholar] [CrossRef]
- Beckler, G.S.; Reeve, J.N. Conservation of primary structure in the hisI gene of the archaebacterium, Methanococcus vannielii, the eubacterium Escherichia coli, and the eucaryote Saccharomyces cerevisiae. MGG Mol. Gen. Genet. 1986, 204, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Chun, J.Y.; Yim, S.H.; Cheon, C.I.; Song, E.; Lee, S.S.; Lee, M.S. Organization and analysis of the histidine biosynthetic genes from Corynebacterium glutamicum. Genes Genom. 2009, 31, 315–323. [Google Scholar] [CrossRef]
- Fondi, M.; Emiliani, G.; Liò, P.; Gribaldo, S.; Fani, R. The evolution of histidine biosynthesis in archaea: Insights into the his genes structure and organization in LUCA. J. Mol. Evol. 2009, 69, 512–526. [Google Scholar] [CrossRef]
- Price, M.N.; Alm, E.J.; Arkin, A.P. The histidine operon is ancient. J. Mol. Evol. 2006, 62, 807–808. [Google Scholar] [CrossRef]
- Fani, R.; Brilli, M.; Liò, P. Inference from proteobacterial operons shows piecewise organization: A reply to Price et al. J. Mol. Evol. 2006, 63, 577–580. [Google Scholar] [CrossRef]
- Arnold, B.J.; Huang, I.T.; Hanage, W.P. Horizontal gene transfer and adaptive evolution in bacteria. Nat. Rev. Microbiol. 2021, 20, 206–218. [Google Scholar] [CrossRef]
- Jain, R.; Rivera, M.C.; Moore, J.E.; Lake, J.A. Horizontal Gene Transfer in Microbial Genome Evolution. Theor. Popul. Biol. 2002, 61, 489–495m. [Google Scholar] [CrossRef] [PubMed]
- Ochman, H.; Lawrence, J.G.; Grolsman, E.A. Lateral gene transfer and the nature of bacterial innovation. Nature 2000, 405, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Soucy, S.M.; Huang, J.; Gogarten, J.P. Horizontal gene transfer: Building the web of life. Nat. Rev. Genet. 2015, 16, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Mira, A.; Ochman, H.; Moran, N.A. Deletional bias and the evolution of bacterial genomes. Trends Genet. 2001, 17, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Copley, S.D. Evolution of new enzymes by gene duplication and divergence. FEBS J. 2020, 287, 1262–1283. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R. Ancient horizontal gene transfer. Nat. Rev. Genet. 2003, 4, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Emamalipour, M.; Seidi, K.; Zununi Vahed, S.; Jahanban-Esfahlan, A.; Jaymand, M.; Majdi, H.; Amoozgar, Z.; Chitkushev, L.T.; Javaheri, T.; Jahanban-Esfahlan, R.; et al. Horizontal Gene Transfer: From Evolutionary Flexibility to Disease Progression. Front. Cell Dev. Biol. 2020, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Faddetta, T.; Vassallo, A.; Del Duca, S.; Gallo, G.; Fani, R.; Puglia, A.M. Unravelling the DNA sequences carried by Streptomyces coelicolor membrane vesicles. Sci. Rep. 2022, 12, 16651. [Google Scholar] [CrossRef]
- Dabizzi, S.; Ammannato, S.; Fani, R. Expression of horizontally transferred gene clusters: Activation by promoter-generating mutations. Res. Microbiol. 2001, 152, 539–549. [Google Scholar] [CrossRef]
- Shieh, Y.W.; Minguez, P.; Bork, P.; Auburger, J.J.; Guilbride, D.L.; Kramer, G.; Bukau, B. Operon structure and cotranslational subunit association direct protein assembly in bacteria. Science 2015, 350, 678–680. [Google Scholar] [CrossRef] [PubMed]
- Fulton, A.B. How crowded is the cytoplasm? Cell 1982, 30, 345–347. [Google Scholar] [CrossRef] [PubMed]
- Mathews, C.K. The cell—Bag of enzymes or network of channels? J. Bacteriol. 1993, 175, 6377–6381. [Google Scholar] [CrossRef]
- Ovádi, J.; Saks, V. On the origin of intracellular compartmentation and organized metabolic systems. Mol. Cell. Biochem. 2004, 256, 5–12. [Google Scholar] [CrossRef]
- Hoppert, M.; Mayer, F. Principles of macromolecular organization and cell function in bacteria and archaea. Cell Biochem. Biophys. 1999, 31, 247–284. [Google Scholar] [CrossRef]
- Meyer, F.M.; Gerwig, J.; Hammer, E.; Herzberg, C.; Commichau, F.M.; Völker, U.; Stülke, J. Physical interactions between tricarboxylic acid cycle enzymes in Bacillus subtilis: Evidence for a metabolon. Metab. Eng. 2011, 13, 18–27. [Google Scholar] [CrossRef]
- Weng, X.; Xiao, J. Spatial organization of transcription in bacterial cells. Trends Genet. 2014, 30, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.B.; Trach, S.O. Estimation of macromolecule concentrations and excluded volume effects for the cytoplasm of Escherichia coli. J. Mol. Biol. 1991, 222, 599–620. [Google Scholar] [CrossRef] [PubMed]
- Srere, P.A. Complexes of sequential metabolic enzymes. Annu. Rev. Biochem. 1987, 56, 89–124. [Google Scholar] [CrossRef]
- McConkey, E.H. Molecular evolution, intracellular organization, and the quinary structure of proteins. Proc. Natl. Acad. Sci. USA 1982, 79, 3236–3240. [Google Scholar] [CrossRef] [PubMed]
- Pedley, A.M.; Benkovic, S.J. A New View into the Regulation of Purine Metabolism: The Purinosome. Trends Biochem. Sci. 2017, 42, 141–154. [Google Scholar] [CrossRef]
- Sweetlove, L.J.; Fernie, A.R. The role of dynamic enzyme assemblies and substrate channelling in metabolic regulation. Nat. Commun. 2018, 9, 2136. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.A.; Ahmad, S. Nested gene fusions as markers of phylogenetic branchpoints in prokaryotes. Trends Ecol. Evol. 1990, 5, 219–224. [Google Scholar] [CrossRef]
- Yanai, I.; Derti, A.; DeLisi, C. Genes linked by fusion events are generally of the same functional category: A systematic analysis of 30 microbial genomes. Proc. Natl. Acad. Sci. USA 2001, 98, 7940–7945. [Google Scholar] [CrossRef] [PubMed]
- Papaleo, M.C.; Russo, E.; Fondi, M.; Emiliani, G.; Frandi, A.; Brilli, M.; Pastorelli, R.; Fani, R. Structural, evolutionary and genetic analysis of the histidine biosynthetic “core” in the genus Burkholderia. Gene 2009, 448, 16–28. [Google Scholar] [CrossRef]
- Chioccioli, S.; Bogani, P.; Del Duca, S.; Castronovo, L.M.; Vassallo, A.; Puglia, A.M.; Fani, R. In vivo evaluation of the interaction between the Escherichia coli IGP synthase subunits using the bacterial two-hybrid system. FEMS Microbiol. Lett. 2020, 367, fnaa112. [Google Scholar] [CrossRef]
- Fani, R.; Liò, P.; Chiarelli, I.; Bazzicalupo, M. The evolution of the histidine biosynthetic genes in prokaryotes: A common ancestor for the hisA and hisF genes. J. Mol. Evol. 1994, 367, fnaa112. [Google Scholar] [CrossRef]
- Horowitz, N.H. The Evolution of Biochemical Syntheses—Retrospect and Prospect. In Evolving Genes and Proteins; Academic Press: Cambridge, MA, USA, 1965; pp. 15–23. [Google Scholar]
- Martin, R.G. The First Enzyme in Histidine Biosynthesis: The Nature of Feedback Inhibition by Histidine. J. Biol. Chem. 1963, 238, 257–268. [Google Scholar] [CrossRef]
- Goldberger, R.F.; Kovach, J.S. Regulation of Histidine Biosynthesis in Salmonella typhimurium. Curr. Top. Cell. Regul. 1972, 5, 285–308. [Google Scholar] [PubMed]
- Riggs, D.L.; Mueller, R.D.; Kwan, H.S.; Artz, S.W. Promoter domain mediates guanosine tetraphosphate activation of the histidine operon. Proc. Natl. Acad. Sci. USA 1986, 83, 9333–9337. [Google Scholar] [CrossRef] [PubMed]
- Ely, B.; Ciesla, Z. Internal promoter P2 of the histidine operon of Salmonella typhimurium. J. Bacteriol. 1974, 120, 984–986. [Google Scholar] [CrossRef]
- Ramos-Montañez, S.; Winkler, M.E. Biosynthesis of Histidine. EcoSal Plus 2009, 3, 1–9. [Google Scholar] [CrossRef]
- Barnes, W.M. DNA sequence from the histidine operon control region: Seven histidine codons in a row. Proc. Natl. Acad. Sci. USA 1978, 75, 4281–4285. [Google Scholar] [CrossRef] [PubMed]
- Alifano, P.; Ciampi, M.S.; Nappo, A.G.; Bruni, C.B.; Carlomagno, M.S. In vivo analysis of the mechanisms responsible for strong transcriptional polarity in a “sense” mutant within an intercistronic region. Cell 1988, 55, 351–360. [Google Scholar] [CrossRef]
- Ciampi, M.S.; Alifano, P.; Nappo, A.G.; Bruni, C.B.; Carlomagno, M.S. Features of the rho-dependent transcription termination polar element within the hisG cistron of Salmonella typhimurium. J. Bacteriol. 1989, 171, 4472–4478. [Google Scholar] [CrossRef]
- Ciampi, M.S. Rho-dependent terminators and transcription termination. Microbiology 2006, 152, 2515–2528. [Google Scholar] [CrossRef]
- Alifano, P.; Rivellini, F.; Nappo, A.G.; Bruni, C.B.; Carlomagno, M.S. Alternative patterns of his operon transcription and mRNA processing generated by metabolic perturbation. Gene 1994, 146, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.N.; Bergendahl, L.T.; Marsh, J.A. Operon Gene Order Is Optimized for Ordered Protein Complex Assembly. Cell Rep. 2016, 14, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Kolesov, G.; Wunderlich, Z.; Laikova, O.N.; Gelfand, M.S.; Mirny, L.A. How gene order is influenced by the biophysics of transcription regulation. Proc. Natl. Acad. Sci. USA 2007, 104, 13948–13953. [Google Scholar] [CrossRef] [PubMed]
- Dame, R.T.; Rashid, F.Z.M.; Grainger, D.C. Chromosome organization in bacteria: Mechanistic insights into genome structure and function. Nat. Rev. Genet. 2020, 21, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Bryant, J.A.; Sellars, L.E.; Busby, S.J.W.; Lee, D.J. Chromosome position effects on gene expression in Escherichia coli K-12. Nucleic Acids Res. 2014, 42, 11383–11392. [Google Scholar] [CrossRef]
- Keene, J.D.; Tenenbaum, S.A. Eukaryotic mRNPs may represent posttranscriptional operons. Mol. Cell 2002, 9, 1161–1167. [Google Scholar] [CrossRef]
- Nair, R.R.; Pataki, E.; Gerst, J.E. Transperons: RNA operons as effectors of coordinated gene expression in eukaryotes. Trends Genet. 2022, 38, 1217–1227. [Google Scholar] [CrossRef]
- Morales-Polanco, F.; Lee, J.H.; Barbosa, N.M.; Frydman, J. Cotranslational Mechanisms of Protein Biogenesis and Complex Assembly in Eukaryotes. Annu. Rev. Biomed. Data Sci. 2022, 5, 67–94. [Google Scholar] [CrossRef]
- Lazcano, A.; Miller, S.L. On the origin of metabolic pathways. J. Mol. Evol. 1999, 49, 424–431. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Name | Reference | Year | N. of Citations | Degree of Interest |
|---|---|---|---|---|
| Fisher | [35] | 1958 | 26,143 | 402 |
| Co-regulation | [36] | 1960 | 616 | 9.8 |
| Molarity | not available | |||
| Selfish operon | [37] | 1996 | 755 | 28.0 |
| Adaptation to thermophily | [38] | 1999 | 49 | 2.0 |
| Protein immobility | [39] | 2004 | 17 | 0.9 |
| Nguyen | [40] | 2019 | 5 | 1.3 |
| Natal | [41] | 1945 | 800 | 10.3 |
| Piecewise | [13] | 2005 | 61 | 3.4 |
| Scribbling pad | [42] | 2013 | 13 | 1.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Duca, S.; Semenzato, G.; Esposito, A.; Liò, P.; Fani, R. The Operon as a Conundrum of Gene Dynamics and Biochemical Constraints: What We Have Learned from Histidine Biosynthesis. Genes 2023, 14, 949. https://doi.org/10.3390/genes14040949
Del Duca S, Semenzato G, Esposito A, Liò P, Fani R. The Operon as a Conundrum of Gene Dynamics and Biochemical Constraints: What We Have Learned from Histidine Biosynthesis. Genes. 2023; 14(4):949. https://doi.org/10.3390/genes14040949
Chicago/Turabian StyleDel Duca, Sara, Giulia Semenzato, Antonia Esposito, Pietro Liò, and Renato Fani. 2023. "The Operon as a Conundrum of Gene Dynamics and Biochemical Constraints: What We Have Learned from Histidine Biosynthesis" Genes 14, no. 4: 949. https://doi.org/10.3390/genes14040949