
Highly Efficient A-to-G Editing in PFFs via Multiple ABEs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Expression Vectors
2.2. Cell Culture and Transfection
2.3. Off-Target Analysis
2.4. Isolation of Single-Cell Colonies Targeting the APOE Gene
2.5. Detection of APOE Expression via qRT-PCR and Western Blotting
2.6. RNA Editing Analysis in APOE Gene-Edited PFF Cells
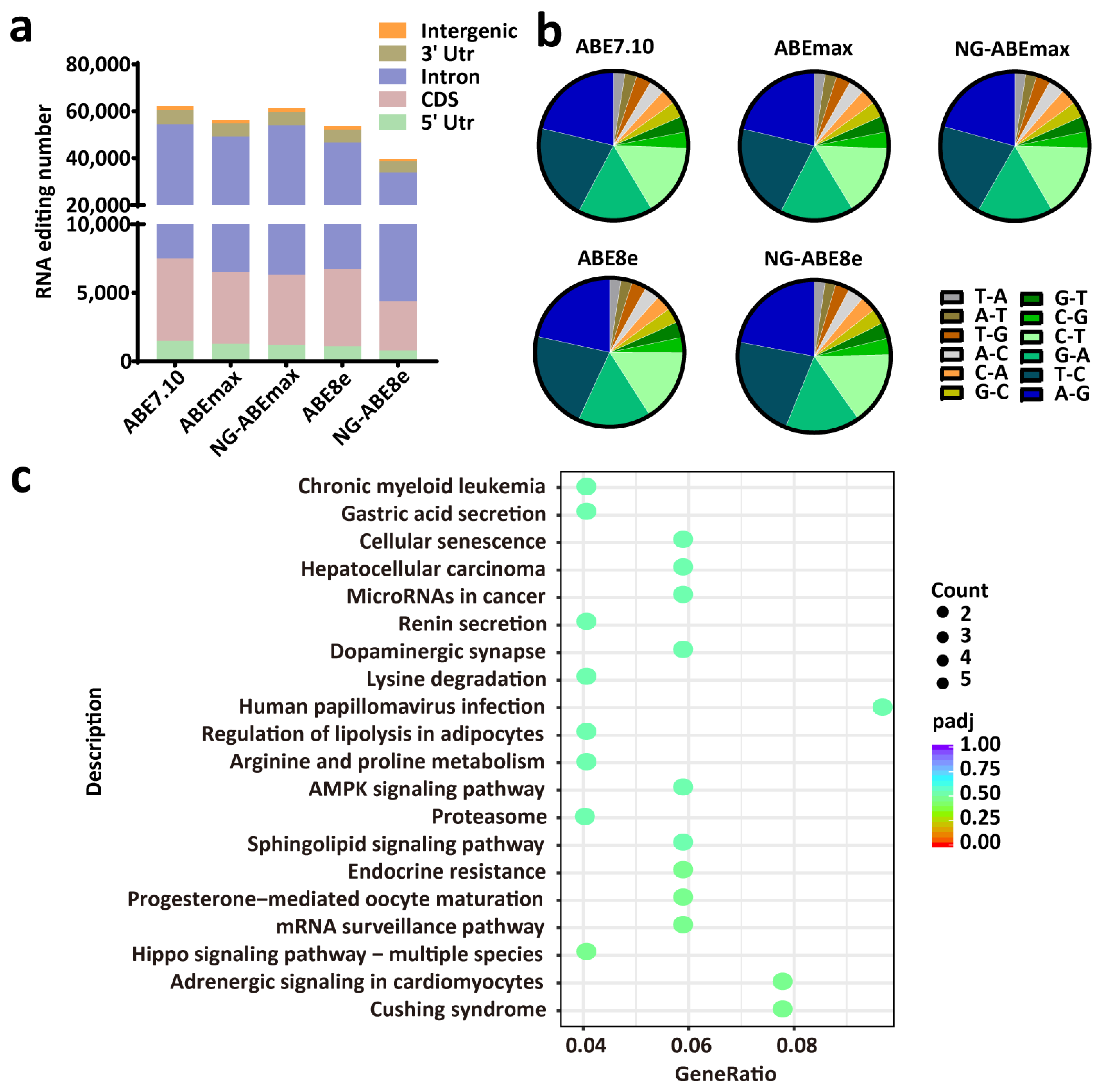
2.7. KEGG Enrichment Analysis of APOE Gene-Edited Cells
2.8. Statistical Analysis
3. Results
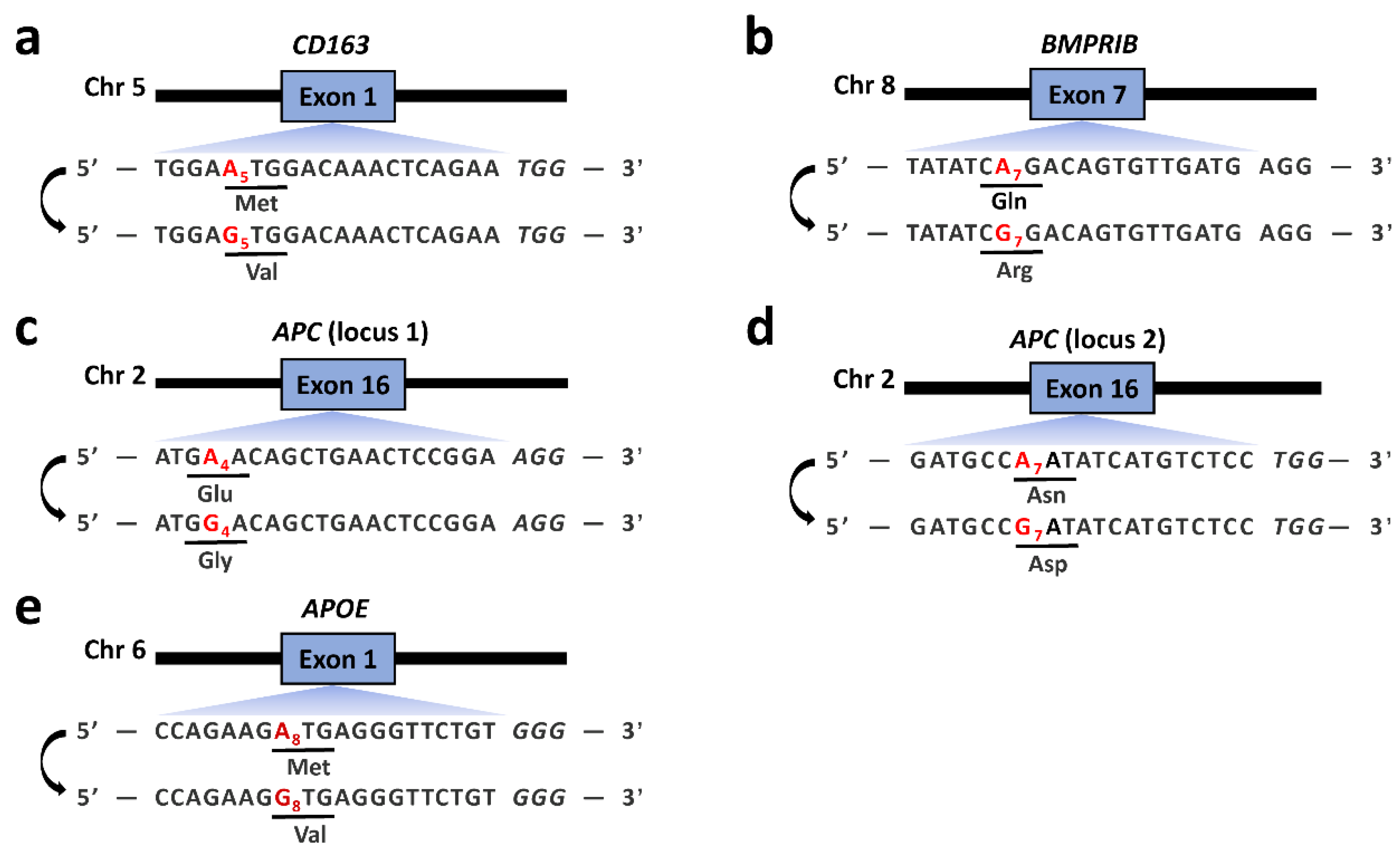
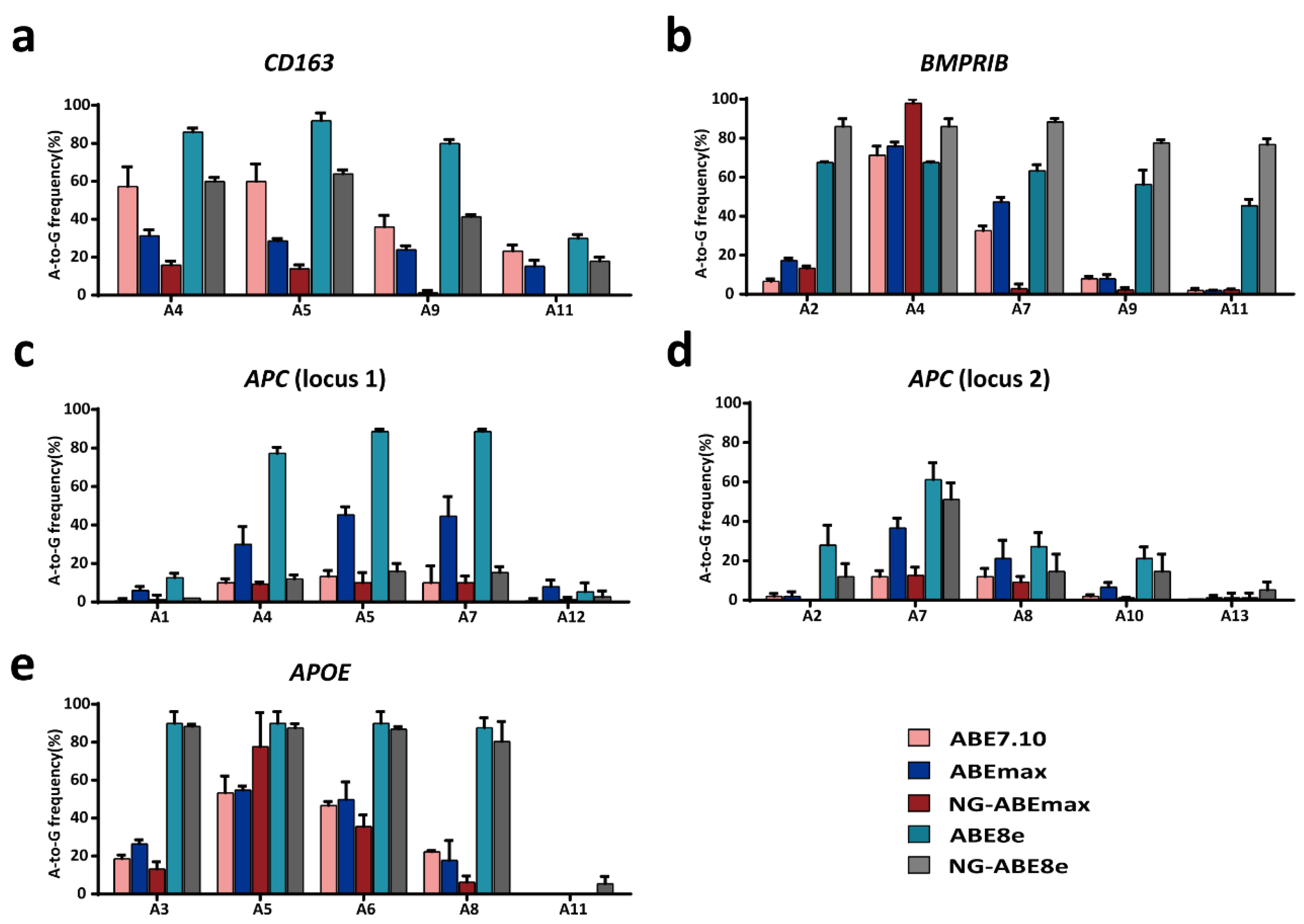
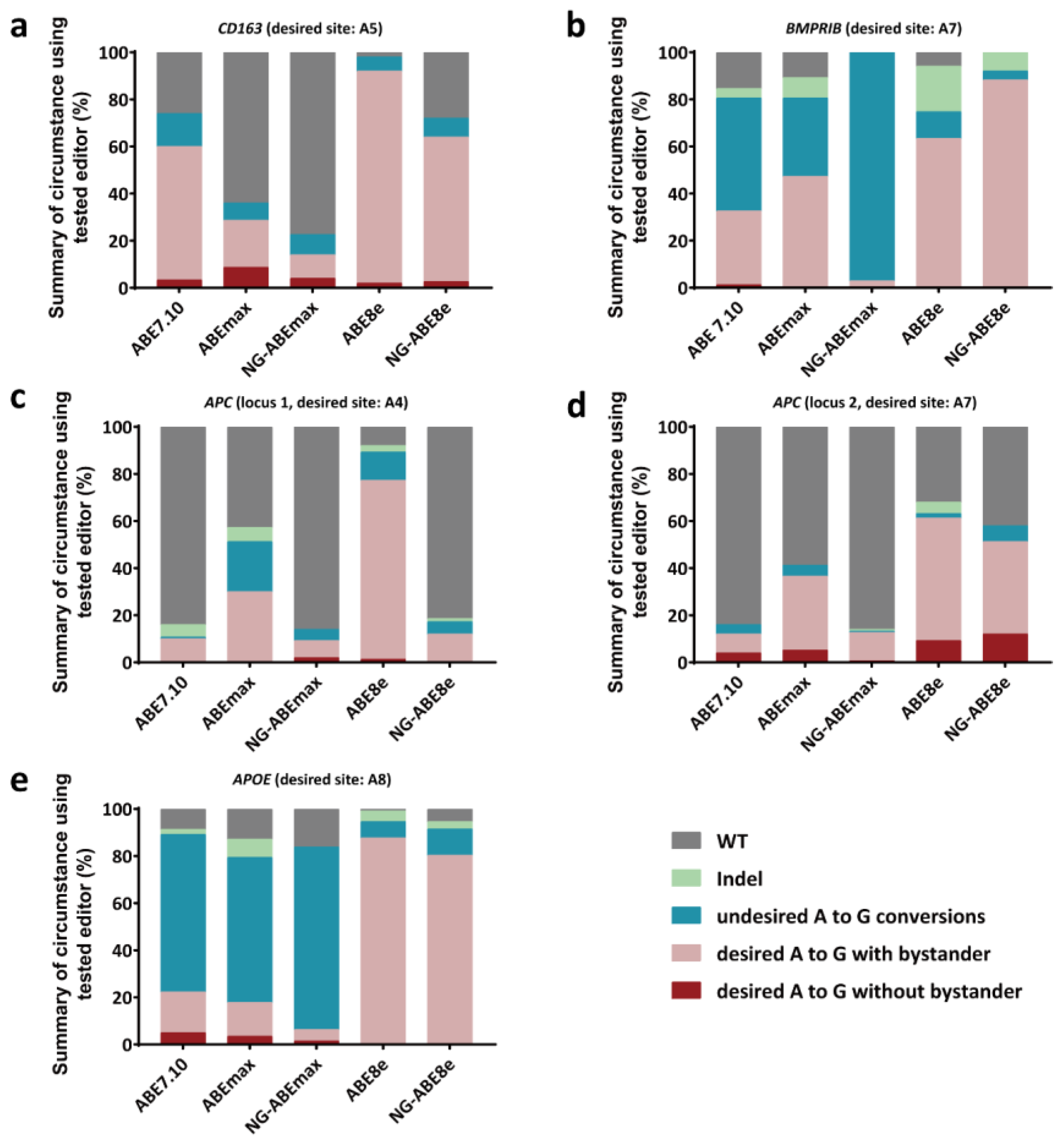
3.1. Variable A-to-G Editing Efficiencies of Different ABEs in the PFFs
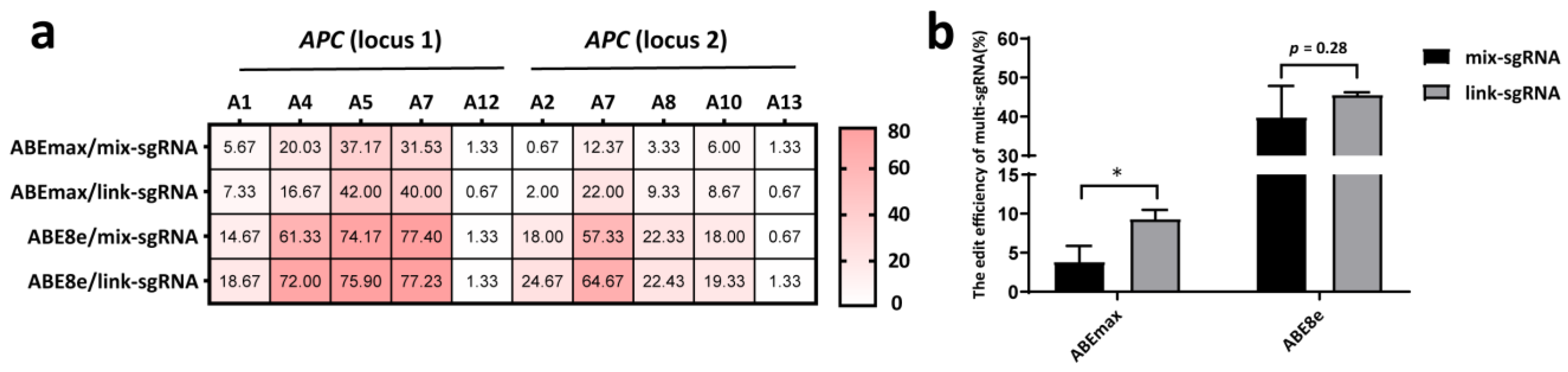
3.2. The Link-sgRNA Strategy Had Superior Editing Efficiencies to the Mix-sgRNA Strategy for Two-site Simultaneous Editing
3.3. Analysis of the DNA Off-Target Effects for Each sgRNA
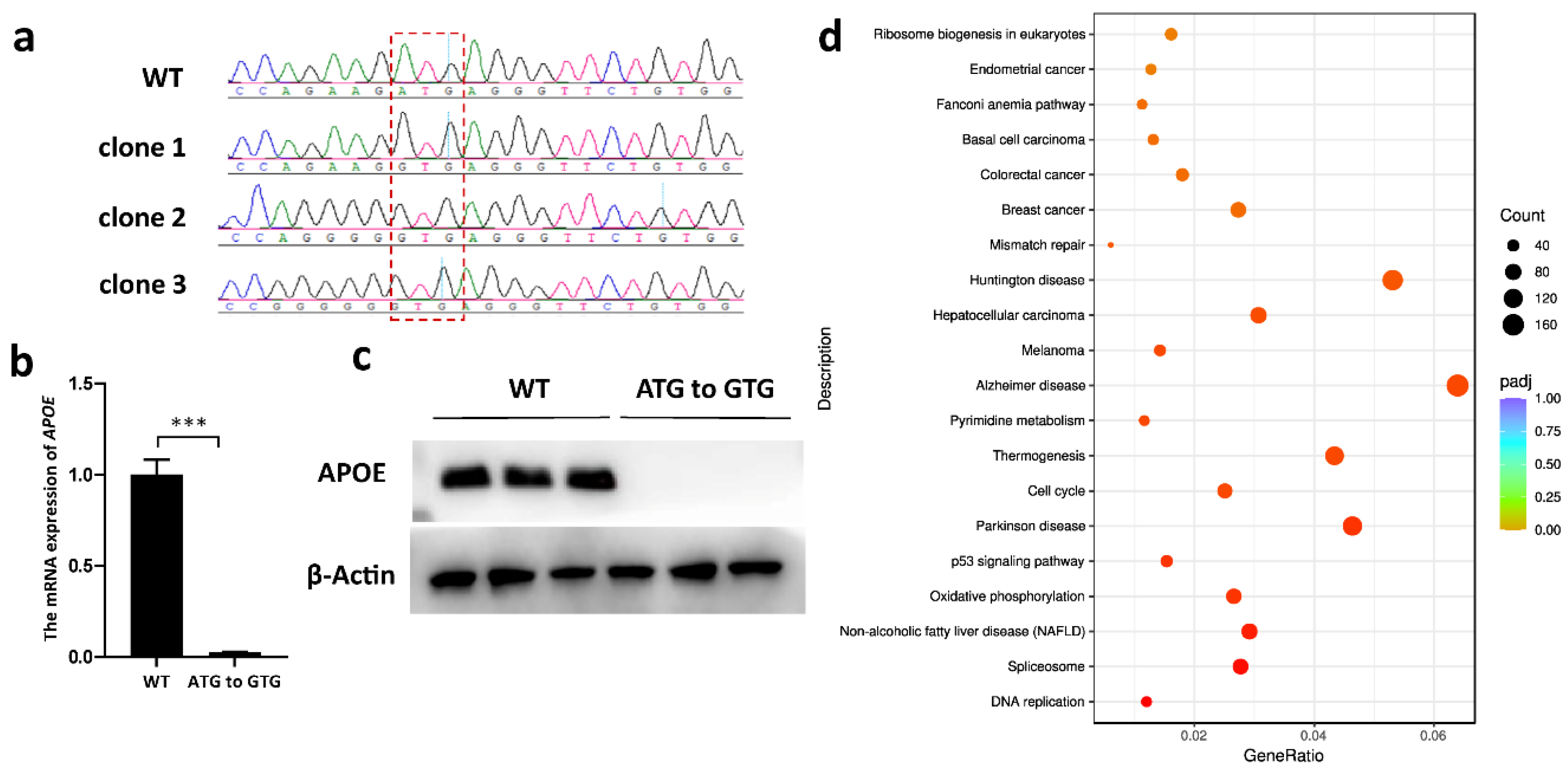
3.4. APOE Expression Was Silenced via ABE-Mediated Start-Codon Mutation
3.5. All Editors Showed Similar Numbers of RNA Editing Sites and Similar Editing-Type Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Janik, E.; Niemcewicz, M.; Ceremuga, M.; Krzowski, L.; Saluk-Bijak, J.; Bijak, M. Various aspects of a gene editing system-crispr-cas9. Int. J. Mol. Sci. 2020, 21, 9604. [Google Scholar] [CrossRef]
- Barman, A.; Deb, B.; Chakraborty, S. A glance at genome editing with CRISPR-Cas9 technology. Curr. Genet. 2020, 66, 447–462. [Google Scholar] [CrossRef]
- Shalem, O.; Sanjana, N.E.; Zhang, F. High-throughput functional genomics using CRISPR-Cas9. Nat. Rev. Genet. 2015, 16, 299–311. [Google Scholar] [CrossRef] [Green Version]
- Kosicki, M.; Tomberg, K.; Bradley, A. Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements. Nat. Biotechnol. 2018, 36, 765–771. [Google Scholar] [CrossRef]
- Adikusuma, F.; Piltz, S.; Corbett, M.A.; Turvey, M.; McColl, S.R.; Helbig, K.J.; Beard, M.R.; Hughes, J.; Pomerantz, R.T.; Thomas, P.Q. Large deletions induced by Cas9 cleavage. Nature 2018, 560, E8–E9. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Qiao, C.; Liu, W.; Jiang, H.; Jing, Q.; Liao, Y.; Ren, J.; Xing, Y. BMPR-IB gene disruption causes severe limb deformities in pigs. Zool. Res. 2022, 43, 391–403. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A• T to G• C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Forner, J.; Karcher, D.; Bock, R. DNA base editing in nuclear and organellar genomes. Trends Genet. 2022, 38, 1147–1169. [Google Scholar] [CrossRef]
- Sasaguri, H.; Nagata, K.; Sekiguchi, M. Introduction of pathogenic mutations into the mouse Psen1 gene by Base Editor and Target-AID. Nat. Commun. 2018, 9, 2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Liu, Z.; Li, G.; Dang, L.; Ma, X. Efficient Gene Silencing by Adenine Base Editor Mediated Start Codon Mutation. Mol. Ther. 2019, 28, 431–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratner, L.D.; La Motta, G.E.; Briski, O.; Salamone, D.F.; Fernandez-Martin, R. Practical approaches for knock-out gene editing in pigs. Front. Genet. 2021, 11, 617850. [Google Scholar] [CrossRef] [PubMed]
- Vajta, G.; Zhang, Y.; Macháty, Z. Somatic cell nuclear transfer in pigs: Recent achievements and future possibilities. Reprod. Fertil. Dev. 2007, 19, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Rakha, A. Cloning efficiency and a comparison between donor cell types. Clon. Transgen. 2015, 2, 141. [Google Scholar] [CrossRef]
- Jiang, H.; Jing, Q.; Yang, Q.; Qiao, C.; Liao, Y.; Liu, W.; Xing, Y. Efficient Simultaneous Introduction of Premature Stop Codons in Three Tumor Suppressor Genes in PFFs via a Cytosine Base Editor. Genes 2022, 13, 835. [Google Scholar] [CrossRef]
- Xie, J.; Huang, X.; Wang, X.; Gou, S.; Lai, L. ACBE, a new base editor for simultaneous C-to-T and A-to-G substitutions in mammalian systems. BMC Biol. 2020, 18, 131. [Google Scholar] [CrossRef]
- Liang, Y.; Xie, J.; Zhang, Q.; Wang, X.; Gou, S.; Lin, L.; Chen, T.; Ge, W.; Zhuang, Z.; Lian, M. AGBE: A dual deaminase-mediated base editor by fusing CGBE with ABE for creating a saturated mutant population with multiple editing patterns. Nucleic Acids Res. 2022, 50, 5384–5399. [Google Scholar] [CrossRef]
- Koblan, L.W.; Doman, J.L.; Wilson, C.; Levy, J.M.; Tay, T.; Newby, G.A.; Liu, D.R. Improving cytidine and adenine base editors by expression optimization and ancestral reconstruction. Nat. Biotechnol. 2018, 36, 843–846. [Google Scholar] [CrossRef]
- Huang, T.P.; Zhao, K.T.; Miller, S.M.; Gaudelli, N.M.; Liu, D.R. Circularly permuted and PAM-modified Cas9 variants broaden the targeting scope of base editors. Nat. Biotechnol. 2019, 37, 626–631. [Google Scholar] [CrossRef]
- Richter, M.F.; Zhao, K.T.; Eton, E.; Lapinaite, A.; Liu, D.R. Phage-assisted evolution of an adenine base editor with improved Cas domain compatibility and activity. Nat. Biotechnol. 2020, 38, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, L.F.S.; Dos Santos Silva, D.B.; Gimenez, D.F.J.; Baldi, F.; Ferro, J.A.; Chardulo, L.A.L.; de Albuquerque, L.G. Gene expression profiling and identification of hub genes in Nellore cattle with different marbling score levels. Genomics 2020, 112, 873–879. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picardi, E.; Pesole, G. REDItools: High-throughput RNA editing detection made easy. Bioinformatics 2013, 29, 1813–1814. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.; Han, Y.; He, Q. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swift, M.L. GraphPad prism, data analysis, and scientific graphing. J. Chem. Inf. Comput. Sci. 1997, 37, 411–412. [Google Scholar] [CrossRef]
- Porto, E.M.; Komor, A.C.; Slaymaker, I.M.; Yeo, G.W. Base editing: Advances and therapeutic opportunities. Nat. Rev. Drug Discov. 2020, 19, 839–859. [Google Scholar] [CrossRef]
- Rees, H.A.; Liu, D.R. Base editing: Precision chemistry on the genome and transcriptome of living cells. Nat. Rev. Genet. 2018, 19, 770–788. [Google Scholar] [CrossRef]
- Newby, G.A.; Liu, D.R. In vivo somatic cell base editing and prime editing. Mol. Ther. 2021, 29, 3107–3124. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, R.; Wu, J.; Xiong, Y.; Wei, J.; Zhang, S.; Yang, B.; Chen, J.; Yang, L. Comparison of cytosine base editors and development of the BEable-GPS database for targeting pathogenic SNVs. Genome Biol. 2019, 20, 218. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Kim, H.K.; Lee, S.; Kim, Y.; Seo, S.Y.; Park, J.; Kim, H. Sequence-specific prediction of the efficiencies of adenine and cytosine base editors. Nat. Biotechnol. 2020, 38, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, Q.; Zhao, K.; Jiang, C.; Ren, D.; Xu, P.; He, X.; Liao, R.; Jiang, K.; Ma, J. Production of transgenic pigs with an introduced missense mutation of the bone morphogenetic protein receptor type IB gene related to prolificacy. Asian-Australas. J. Anim. Sci. 2016, 29, 925–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.P.; Newby, G.A.; Liu, D.R. Precision genome editing using cytosine and adenine base editors in mammalian cells. Nat. Protoc. 2021, 16, 1089–1128. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Sun, H.; Zhang, X.; Xie, X.; Zhang, J.; Bai, Y.; Ouyang, X.; Zhi, S.; Xiong, Y.; Ma, W. Effective and precise adenine base editing in mouse zygotes. Protein Cell 2018, 9, 808–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Aida, T.; Del Rosario, R.C.; Wilde, J.J.; Ding, C.; Zhang, X.; Baloch, Z.; Huang, Y.; Tang, Y.; Li, D. Multiplex precise base editing in cynomolgus monkeys. Nat. Commun. 2020, 11, 2325. [Google Scholar] [CrossRef]
- Zhou, S.; Ding, Y.; Liu, J.; Liu, Y.; Zhao, X.; Li, G.; Zhang, C.; Li, C.; Wang, Y.; Kalds, P. Highly efficient generation of sheep with a defined FecBB mutation via adenine base editing. Genet. Sel. Evol. 2020, 52, 35. [Google Scholar] [CrossRef]
- Chen, F.; Lian, M.; Ma, B.; Gou, S.; Luo, X.; Yang, K.; Shi, H.; Xie, J.; Ge, W.; Ouyang, Z. Multiplexed base editing through Cas12a variant-mediated cytosine and adenine base editors. Commun. Biol. 2022, 5, 1163. [Google Scholar] [CrossRef]
- Grünewald, J.; Zhou, R.; Garcia, S.P.; Iyer, S.; Lareau, C.A.; Aryee, M.J.; Joung, J.K. Transcriptome-wide off-target RNA editing induced by CRISPR-guided DNA base editors. Nature 2019, 569, 433–437. [Google Scholar] [CrossRef]
- Jeong, Y.; Lee, S.; Hwang, G.; Hong, G.H.; Bae, S. Adenine base editor engineering reduces editing of bystander cytosines. Nat. Biotechnol. 2021, 39, 1426–1433. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, S.; Xue, N.; Hong, M.; Li, D. Engineering a precise adenine base editor with minimal bystander editing. Nat. Chem. Biol. 2023, 19, 101–110. [Google Scholar] [CrossRef]
- Sakuma, T.; Nishikawa, A.; Kume, S.; Chayama, K.; Yamamoto, T. Multiplex genome engineering in human cells using all-in-one CRISPR/Cas9 vector system. Sci. Rep. 2014, 4, 5400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Wu, L.; Zhang, S.; Lu, M.; Cheung, W.K.; Cai, W.; Gale, M.; Xu, Q.; Yan, Q. An easy and efficient inducible CRISPR/Cas9 platform with improved specificity for multiple gene targeting. Nucleic Acids Res. 2016, 44, e149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Xie, W.; Liu, Z.; Shan, H.; Chen, M.; Song, Y.; Yu, H.; Lai, L.; Li, Z. CRISPR start-loss: A novel and practical alternative for gene silencing through base-editing-induced start codon mutations. Mol. Ther.-Nucl. Acids 2020, 21, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Histone modifications for human epigenome analysis. J. Hum. Genet. 2013, 58, 439–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Zhou, Z.; Huang, T.; Zhang, Z.; Li, W.; Ling, Z.; Jiang, T.; Yang, J.; Yang, B.; Huang, L. Mapping and analysis of a spatiotemporal H3K27ac and gene expression spectrum in pigs. Sci. China Life Sci. 2022, 65, 1517–1534. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Fu, A.K.; Ip, N.Y. APOE signaling in neurodegenerative diseases: An integrative approach targeting APOE coding and noncoding variants for disease intervention. Curr. Opin. Neurobiol. 2021, 69, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Slattery, M.L.; Sweeney, C.; Murtaugh, M.; Ma, K.N.; Potter, J.D.; Levin, T.R.; Samowitz, W.; Wolff, R. Associations between apoE genotype and colon and rectal cancer. Carcinogenesis 2005, 26, 1422–1429. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Zhang, H.; Sun, W.; Gong, H.; Wang, Y.; Ma, C.; Wang, J.; Cao, C.; Yang, X.; Tian, J. Hyperhomocysteinemia induces cardiac injury by up-regulation of p53-dependent Noxa and Bax expression through the p53 DNA methylation in ApoE−/− mice. Acta Biochim. Biophys. Sin. 2013, 45, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Etzerodt, A.; Moestrup, S.K. CD163 and inflammation: Biological, diagnostic, and therapeutic aspects. Antioxid. Redox Signal. 2013, 18, 2352–2363. [Google Scholar] [CrossRef] [Green Version]
- Slesarenko, Y.S.; Lavrov, A.V.; Smirnikhina, S.A. Off-target effects of base editors: What we know and how we can reduce it. Curr. Genet. 2022, 68, 39–48. [Google Scholar] [CrossRef]
- Zhou, C.; Sun, Y.; Yan, R.; Liu, Y.; Zuo, E.; Gu, C.; Han, L.; Wei, Y.; Hu, X.; Zeng, R. Off-target RNA mutation induced by DNA base editing and its elimination by mutagenesis. Nature 2019, 571, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.K.; Jo, D.H.; Lee, S.N.; Cho, C.S.; Jung, Y.; Bae, S. High-purity production and precise editing of DNA base editing ribonucleoproteins. Sci. Adv. 2021, 7, eabg2661. [Google Scholar] [CrossRef]
- Kim, D.; Kim, D.E.; Lee, G.; Cho, S.I.; Kim, J.S. Genome-wide target specificity of CRISPR RNA-guided adenine base editors. Nat. Biotechnol. 2019, 37, 430–435. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, Q.; Liu, W.; Jiang, H.; Liao, Y.; Yang, Q.; Xing, Y. Highly Efficient A-to-G Editing in PFFs via Multiple ABEs. Genes 2023, 14, 908. https://doi.org/10.3390/genes14040908
Jing Q, Liu W, Jiang H, Liao Y, Yang Q, Xing Y. Highly Efficient A-to-G Editing in PFFs via Multiple ABEs. Genes. 2023; 14(4):908. https://doi.org/10.3390/genes14040908
Chicago/Turabian StyleJing, Qiqi, Weiwei Liu, Haoyun Jiang, Yaya Liao, Qiang Yang, and Yuyun Xing. 2023. "Highly Efficient A-to-G Editing in PFFs via Multiple ABEs" Genes 14, no. 4: 908. https://doi.org/10.3390/genes14040908