
Strategies for the Generation of Gene Modified Avian Models: Advancement in Avian Germline Transmission, Genome Editing, and Applications

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Historical Overview of Genome Modification Strategies for Avian Model Development
2.1. Overview of Avian Transgenesis
2.2. Virus-Mediated Gene Insertion for Avian Transgenesis
2.3. Development of an Avian PGC Culture System and Transgenic Models
2.4. Development of Efficient Germline Chimeras by Depleting Endogenous PGCs for Avian Transgenesis
2.5. Transgenic Systems Using Other Germline-Competent Stem Cells in Avian Species
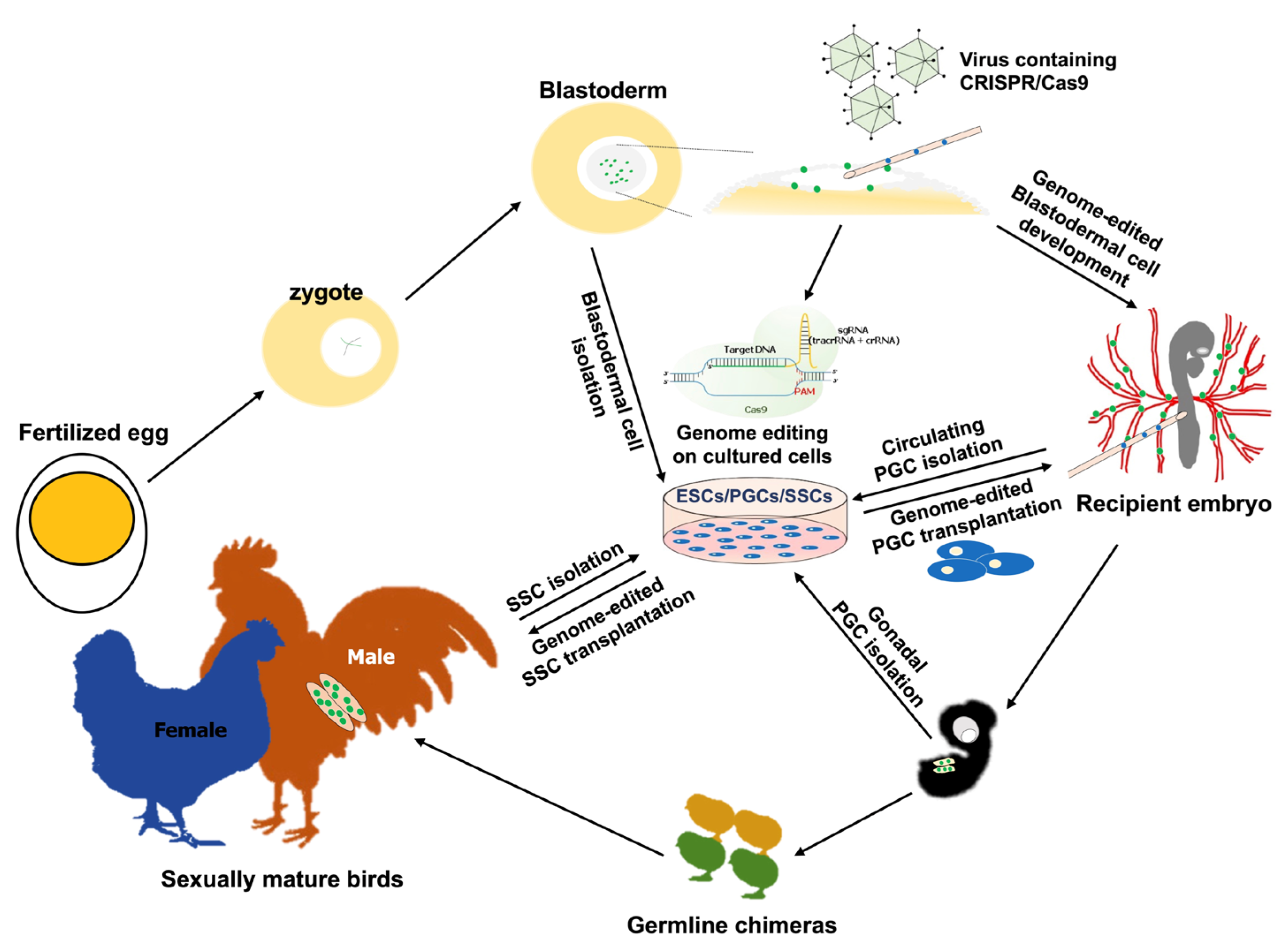
3. Progression of Precise Genome-Editing Systems for Avian Taxa
3.1. Programmable Genome Editing in the Avian System
3.2. CRISPR-Mediated Genome Editing in the Avian System
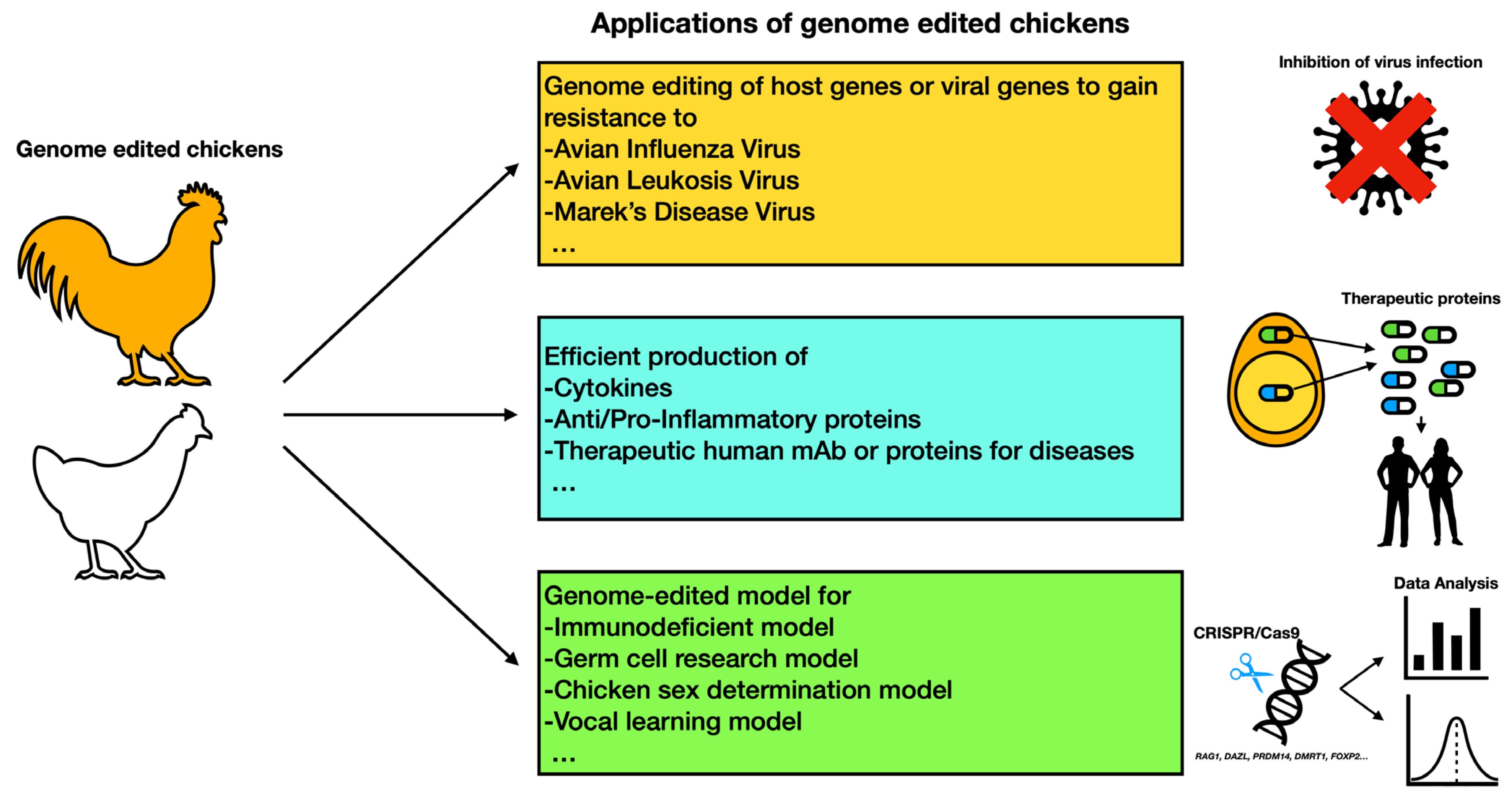
4. Potential Applications of Genome Editing for Avian Model Development
4.1. Genome Editing for the Development of Disease-Resistant Avian Models
4.2. Practical Bioreactor System for Recombinant Protein Production in Avian Systems
4.3. Genome-Edited Birds as Scientific Models
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Chicken Genome Sequencing Consortium. Sequencing and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, C.D. The chick; A great model system becomes even greater. Dev. Cell 2005, 8, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tizard, M.L.; Jenkins, K.A.; Cooper, C.A.; Woodcock, M.E.; Challagulla, A.; Doran, T.J. Potential benefits of gene editing for the future of poultry farming. Transgenic Res. 2019, 28, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Looi, F.Y.; Baker, M.L.; Townson, T.; Richard, M.; Novak, B.; Doran, T.J.; Short, K.R. Creating Disease Resistant Chickens: A Viable Solution to Avian Influenza? Viruses 2018, 10, 561. [Google Scholar] [CrossRef] [Green Version]
- Han, J.Y.; Park, Y.H. Primordial germ cell-mediated transgenesis and genome editing in birds. J. Anim. Sci. Biotechnol. 2018, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Herron, L.R.; Pridans, C.; Turnbull, M.L.; Smith, N.; Lillico, S.; Sherman, A.; Gilhooley, H.J.; Wear, M.; Kurian, D.; Papadakos, G.; et al. A chicken bioreactor for efficient production of functional cytokines. BMC Biotechnol. 2018, 18, 82. [Google Scholar] [CrossRef]
- Woodfint, R.M.; Hamlin, E.; Lee, K. Avian Bioreactor Systems: A Review. Mol. Biotechnol. 2018, 60, 975–983. [Google Scholar] [CrossRef]
- Mozdziak, P.E.; Petitte, J.N. Status of transgenic chicken models for developmental biology. Dev. Dyn. 2004, 229, 414–421. [Google Scholar] [CrossRef]
- Mottet, A.; Tempio, G. Global poultry production: Current state and future outlook and challenges. World’s Poult. Sci. J. 2017, 73, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Peacock, T.H.P.; James, J.; Sealy, J.E.; Iqbal, M. A Global Perspective on H9N2 Avian Influenza Virus. Viruses 2019, 11, 620. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.W. Transgenic Technology and Laboratory Animal Science. ILAR J. 1997, 38, 32–41. [Google Scholar] [CrossRef]
- Serralbo, O.; Picard, C.A.; Marcelle, C. Long-term, inducible gene loss-of-function in the chicken embryo. Genesis 2013, 51, 372–380. [Google Scholar] [CrossRef]
- Han, J.Y.; Lee, H.C.; Park, T.S. Germline-competent stem cell in avian species and its application. Asian J. Androl. 2015, 17, 421–426. [Google Scholar] [CrossRef]
- Gordon, J.W.; Scangos, G.A.; Plotkin, D.J.; Barbosa, J.A.; Ruddle, F.H. Genetic transformation of mouse embryos by microinjection of purified DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 7380–7384. [Google Scholar] [CrossRef] [Green Version]
- Hammer, R.E.; Pursel, V.G.; Rexroad, C.E., Jr.; Wall, R.J.; Bolt, D.J.; Ebert, K.M.; Palmiter, R.D.; Brinster, R.L. Production of transgenic rabbits, sheep and pigs by microinjection. Nature 1985, 315, 680–683. [Google Scholar] [CrossRef] [Green Version]
- McMahon, A.P.; Flytzanis, C.N.; Hough-Evans, B.R.; Katula, K.S.; Britten, R.J.; Davidson, E.H. Introduction of cloned DNA into sea urchin egg cytoplasm: Replication and persistence during embryogenesis. Dev. Biol. 1985, 108, 420–430. [Google Scholar] [CrossRef]
- Rusconi, S.; Schaffner, W. Transformation of frog embryos with a rabbit beta-globin gene. Proc. Natl. Acad. Sci. USA 1981, 78, 5051–5055. [Google Scholar] [CrossRef] [Green Version]
- Spradling, A.C.; Rubin, G.M. Transposition of cloned P elements into Drosophila germ line chromosomes. Science 1982, 218, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Rubin, G.M.; Spradling, A.C. Genetic transformation of Drosophila with transposable element vectors. Science 1982, 218, 348–353. [Google Scholar] [CrossRef] [Green Version]
- Lanigan, T.M.; Kopera, H.C.; Saunders, T.L. Principles of Genetic Engineering. Genes 2020, 11, 291. [Google Scholar] [CrossRef] [Green Version]
- Fofanova, K.A. Morphologic Data on Polyspermy in Chickens. Fed. Proc. Transl. Suppl. 1965, 24, 239–247. [Google Scholar] [PubMed]
- Lee, H.C.; Choi, H.J.; Park, T.S.; Lee, S.I.; Kim, Y.M.; Rengaraj, D.; Nagai, H.; Sheng, G.; Lim, J.M.; Han, J.Y. Cleavage events and sperm dynamics in chick intrauterine embryos. PLoS ONE 2013, 8, e80631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waddington, D.; Gribbin, C.; Sterling, R.J.; Sang, H.M.; Perry, M.M. Chronology of events in the first cell cycle of the polyspermic egg of the domestic fowl (Gallus domesticus). Int. J. Dev. Biol. 1998, 42, 625–628. [Google Scholar] [PubMed]
- Eyal-Giladi, H.; Kochav, S. From cleavage to primitive streak formation: A complementary normal table and a new look at the first stages of the development of the chick. I. General morphology. Dev. Biol. 1976, 49, 321–337. [Google Scholar] [CrossRef]
- Khaner, O.; Mitrani, E.; Eyal-Giladi, H. Developmental potencies of area opaca and marginal zone areas of early chick blastoderms. J. Embryol. Exp. Morphol. 1985, 89, 235–241. [Google Scholar] [CrossRef]
- Tsunekawa, N.; Naito, M.; Sakai, Y.; Nishida, T.; Noce, T. Isolation of chicken vasa homolog gene and tracing the origin of primordial germ cells. Development 2000, 127, 2741–2750. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yamamoto, Y.; Usui, F.; Mushika, T.; Ono, T.; Setioko, A.R.; Takeda, K.; Nirasawa, K.; Kagami, H.; Tagami, T. Migration and proliferation of primordial germ cells in the early chicken embryo. Poult. Sci. 2007, 86, 2182–2193. [Google Scholar] [CrossRef]
- Lee, H.C.; Choi, H.J.; Lee, H.G.; Lim, J.M.; Ono, T.; Han, J.Y. DAZL Expression Explains Origin and Central Formation of Primordial Germ Cells in Chickens. Stem Cells Dev. 2016, 25, 68–79. [Google Scholar] [CrossRef]
- Love, J.; Gribbin, C.; Mather, C.; Sang, H. Transgenic birds by DNA microinjection. Biotechnology 1994, 12, 60–63. [Google Scholar] [CrossRef]
- Salter, D.W.; Smith, E.J.; Hughes, S.H.; Wright, S.E.; Fadly, A.M.; Witter, R.L.; Crittenden, L.B. Gene insertion into the chicken germ line by retroviruses. Poult. Sci. 1986, 65, 1445–1458. [Google Scholar] [CrossRef]
- Salter, D.W.; Smith, E.J.; Hughes, S.H.; Wright, S.E.; Crittenden, L.B. Transgenic chickens: Insertion of retroviral genes into the chicken germ line. Virology 1987, 157, 236–240. [Google Scholar] [CrossRef]
- Bosselman, R.A.; Hsu, R.Y.; Boggs, T.; Hu, S.; Bruszewski, J.; Ou, S.; Kozar, L.; Martin, F.; Green, C.; Jacobsen, F.; et al. Germline transmission of exogenous genes in the chicken. Science 1989, 243, 533–535. [Google Scholar] [CrossRef]
- Harvey, A.J.; Speksnijder, G.; Baugh, L.R.; Morris, J.A.; Ivarie, R. Expression of exogenous protein in the egg white of transgenic chickens. Nat. Biotechnol. 2002, 20, 396–399. [Google Scholar] [CrossRef]
- McGrew, M.J.; Sherman, A.; Ellard, F.M.; Lillico, S.G.; Gilhooley, H.J.; Kingsman, A.J.; Mitrophanous, K.A.; Sang, H. Efficient production of germline transgenic chickens using lentiviral vectors. EMBO Rep. 2004, 5, 728–733. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kim, D.-H.; Karolak, M.C.; Shin, S.; Lee, K. Generation of genome-edited chicken and duck lines by adenovirus-mediated in vivo genome editing. Proc. Natl. Acad. Sci. USA 2022, 119, e2214344119. [Google Scholar] [CrossRef]
- Mizuarai, S.; Ono, K.; Yamaguchi, K.; Nishijima, K.; Kamihira, M.; Iijima, S. Production of transgenic quails with high frequency of germ-line transmission using VSV-G pseudotyped retroviral vector. Biochem. Biophys. Res. Commun. 2001, 286, 456–463. [Google Scholar] [CrossRef]
- Shin, S.S.; Kim, T.M.; Kim, S.Y.; Kim, T.W.; Seo, H.W.; Lee, S.K.; Kwon, S.C.; Lee, G.S.; Kim, H.; Lim, J.M.; et al. Generation of transgenic quail through germ cell-mediated germline transmission. FASEB J. 2008, 22, 2435–2444. [Google Scholar] [CrossRef]
- Agate, R.J.; Scott, B.B.; Haripal, B.; Lois, C.; Nottebohm, F. Transgenic songbirds offer an opportunity to develop a genetic model for vocal learning. Proc. Natl. Acad. Sci. USA 2009, 106, 17963–17967. [Google Scholar] [CrossRef] [Green Version]
- Abe, K.; Matsui, S.; Watanabe, D. Transgenic songbirds with suppressed or enhanced activity of CREB transcription factor. Proc. Natl. Acad. Sci. USA 2015, 112, 7599–7604. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.C.; Kohn, J.; Szwed, S.K.; Pariser, E.; Sepe, S.; Haripal, B.; Oshimori, N.; Marsala, M.; Miyanohara, A.; Lee, R. Human mutant huntingtin disrupts vocal learning in transgenic songbirds. Nat. Neurosci. 2015, 18, 1617–1622. [Google Scholar] [CrossRef]
- Gessara, I.; Dittrich, F.; Hertel, M.; Hildebrand, S.; Pfeifer, A.; Frankl-Vilches, C.; McGrew, M.; Gahr, M. Highly Efficient Genome Modification of Cultured Primordial Germ Cells with Lentiviral Vectors to Generate Transgenic Songbirds. Stem Cell Rep. 2021, 16, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Ma, J.; Lee, K. Direct delivery of adenoviral CRISPR/Cas9 vector into the blastoderm for generation of targeted gene knockout in quail. Proc. Natl. Acad. Sci. USA 2019, 116, 13288–13292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, B.E.; Lehmann, R. Mechanisms guiding primordial germ cell migration: Strategies from different organisms. Nat. Rev. Mol. Cell Biol. 2010, 11, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, Y.; Kagami, H.; Tagami, T. Development, differentiation and manipulation of chicken germ cells. Dev. Growth Differ. 2013, 55, 20–40. [Google Scholar] [CrossRef]
- Kang, K.S.; Lee, H.C.; Kim, H.J.; Lee, H.G.; Kim, Y.M.; Lee, H.J.; Park, Y.H.; Yang, S.Y.; Rengaraj, D.; Park, T.S.; et al. Spatial and temporal action of chicken primordial germ cells during initial migration. Reproduction 2015, 149, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Paksa, A.; Raz, E. Zebrafish germ cells: Motility and guided migration. Curr. Opin. Cell Biol. 2015, 36, 80–85. [Google Scholar] [CrossRef]
- Sasado, T.; Morinaga, C.; Niwa, K.; Shinomiya, A.; Yasuoka, A.; Suwa, H.; Hirose, Y.; Yoda, H.; Henrich, T.; Deguchi, T.; et al. Mutations affecting early distribution of primordial germ cells in Medaka (Oryzias latipes) embryo. Mech. Dev. 2004, 121, 817–828. [Google Scholar] [CrossRef]
- Hayashi, K.; de Sousa Lopes, S.M.; Surani, M.A. Germ cell specification in mice. Science 2007, 316, 394–396. [Google Scholar] [CrossRef]
- Lawson, K.A.; Hage, W.J. Clonal analysis of the origin of primordial germ cells in the mouse. In Ciba Foundation Symposium 182-Germline Development: Germline Development: Ciba Foundation Symposium 182; John Willey & Sons: Chichester, UK, 1994; Volume 182, pp. 68–84, discussion 84–91. [Google Scholar]
- Ginsburg, M.; Eyal-Giladi, H. Primordial germ cells of the young chick blastoderm originate from the central zone of the area pellucida irrespective of the embryo-forming process. Development 1987, 101, 209–219. [Google Scholar] [CrossRef]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef]
- Swift, C.H. Origin and early history of the primordial germ cells of the chick. Am. J. Anat. 1914, 15, 483–516. [Google Scholar] [CrossRef] [Green Version]
- Eyal-Giladi, H.; Ginsburg, M.; Farbarov, A. Avian primordial germ cells are of epiblastic origin. J. Embryol. Exp. Morphol. 1981, 65, 139–147. [Google Scholar] [CrossRef]
- Ukeshima, A.; Yoshinaga, K.; Fujimoto, T. Scanning and transmission electron microscopic observations of chick primordial germ cells with special reference to the extravasation in their migration course. J. Electron. Microsc. 1991, 40, 124–128. [Google Scholar]
- Fujimoto, T.; Ukeshima, A.; Kiyofuji, R. The origin, migration and morphology of the primordial germ cells in the chick embryo. Anat. Rec. 1976, 185, 139–145. [Google Scholar] [CrossRef]
- Meyer, D.B. The Migration of Primordial Germ Cells in the Chick Embryo. Dev. Biol. 1964, 10, 154–190. [Google Scholar] [CrossRef]
- Niewkoop, P.; Sutasurya, L. Primordial Germ Cells in the Chordates; Cambridge University Press: Cambridge, UK, 1979; pp. 118–123. [Google Scholar]
- Kim, Y.M.; Han, J.Y. The early development of germ cells in chicken. Int. J. Dev. Biol. 2018, 62, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Han, J.Y.; Cho, H.Y.; Kim, Y.M.; Park, K.J.; Jung, K.M.; Park, J.S. Production of quail (Coturnix japonica) germline chimeras by transfer of Ficoll-enriched spermatogonial stem cells. Theriogenology 2020, 154, 223–231. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Usui, F.; Nakamura, Y.; Ito, Y.; Tagami, T.; Nirasawa, K.; Matsubara, Y.; Ono, T.; Kagami, H. A Novel Method to Isolate Primordial Germ Cells and Its Use for the Generation of Germline Chimeras in Chicken1. Biol. Reprod. 2007, 77, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Chaipipat, S.; Prukudom, S.; Sritabtim, K.; Kuwana, T.; Piyasanti, Y.; Sinsiri, R.; Piantham, C.; Sangkalerd, S.; Boonsanong, S.; Pitiwong, K.; et al. Primordial germ cells isolated from individual embryos of red junglefowl and indigenous pheasants of Thailand. Theriogenology 2021, 165, 59–68. [Google Scholar] [CrossRef]
- Wade, A.J.; French, N.A.; Ireland, G.W. The potential for archiving and reconstituting valuable strains of turkey (Meleagris gallopavo) using primordial germ cells. Poult. Sci. 2014, 93, 799–809. [Google Scholar] [CrossRef]
- Jung, K.; Kim, Y.; Ono, T.; Han, J. Size-dependent isolation of primordial germ cells from avian species. Mol. Reprod. Dev. 2017, 84, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Lin, S.P.; Chang, Y.Y.; Chang, W.P.; Wei, L.Y.; Liu, H.C.; Huang, J.F.; Pain, B.; Wu, S.C. In vitro culture and characterization of duck primordial germ cells. Poult. Sci. 2019, 98, 1820–1832. [Google Scholar] [CrossRef] [PubMed]
- Mozdziak, P.E.; Angerman-Stewart, J.; Rushton, B.; Pardue, S.L.; Petitte, J.N. Isolation of chicken primordial germ cells using fluorescence-activated cell sorting. Poult. Sci. 2005, 84, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Ono, T.; Machida, Y. Immunomagnetic purification of viable primordial germ cells of Japanese quail (Coturnix japonica). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1999, 122, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.F.; Kuwana, T. Purification of avian circulating primordial germ cells by Nycodenz density gradient centrifugation. Br. Poult. Sci. 2003, 44, 30–35. [Google Scholar] [CrossRef]
- Chang, I.; Tajima, A.; Yasuda, Y.; Chikamune, T.; Ohno, T. Simple method for isolation of primordial germ cells from chick embryos. Cell Biol. Int. Rep. 1992, 16, 853–857. [Google Scholar] [CrossRef]
- Ono, T.; Yokoi, R.; Aoyama, H. Transfer of male or female primordial germ cells of quail into chick embryonic gonads. Exp. Anim. 1996, 45, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Reynaud, G. The transfer of turkey primordial germ cells to chick embryos by intravascular injection. J. Embryol. Exp. Morphol. 1969, 21, 485–507. [Google Scholar]
- Liu, C.; Khazanehdari, K.A.; Baskar, V.; Saleem, S.; Kinne, J.; Wernery, U.; Chang, I.K. Production of chicken progeny (Gallus gallus domesticus) from interspecies germline chimeric duck (Anas domesticus) by primordial germ cell transfer. Biol. Reprod. 2012, 86, 101. [Google Scholar] [CrossRef] [Green Version]
- Van de Lavoir, M.C.; Collarini, E.J.; Leighton, P.A.; Fesler, J.; Lu, D.R.; Harriman, W.D.; Thiyagasundaram, T.S.; Etches, R.J. Interspecific germline transmission of cultured primordial germ cells. PLoS ONE 2012, 7, e35664. [Google Scholar] [CrossRef]
- Kang, S.J.; Choi, J.W.; Kim, S.Y.; Park, K.J.; Kim, T.M.; Lee, Y.M.; Kim, H.; Lim, J.M.; Han, J.Y. Reproduction of wild birds via interspecies germ cell transplantation. Biol. Reprod. 2008, 79, 931–937. [Google Scholar] [CrossRef] [Green Version]
- Wernery, U.; Liu, C.; Baskar, V.; Guerineche, Z.; Khazanehdari, K.A.; Saleem, S.; Kinne, J.; Wernery, R.; Griffin, D.K.; Chang, I.K. Primordial germ cell-mediated chimera technology produces viable pure-line Houbara bustard offspring: Potential for repopulating an endangered species. PLoS ONE 2010, 5, e15824. [Google Scholar] [CrossRef] [Green Version]
- Van de Lavoir, M.C.; Diamond, J.H.; Leighton, P.A.; Mather-Love, C.; Heyer, B.S.; Bradshaw, R.; Kerchner, A.; Hooi, L.T.; Gessaro, T.M.; Swanberg, S.E.; et al. Germline transmission of genetically modified primordial germ cells. Nature 2006, 441, 766–769. [Google Scholar] [CrossRef]
- Choi, J.W.; Kim, S.; Kim, T.M.; Kim, Y.M.; Seo, H.W.; Park, T.S.; Jeong, J.W.; Song, G.; Han, J.Y. Basic fibroblast growth factor activates MEK/ERK cell signaling pathway and stimulates the proliferation of chicken primordial germ cells. PLoS ONE 2010, 5, e12968. [Google Scholar] [CrossRef]
- Macdonald, J.; Glover, J.D.; Taylor, L.; Sang, H.M.; McGrew, M.J. Characterisation and germline transmission of cultured avian primordial germ cells. PLoS ONE 2010, 5, e15518. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.C.; Lim, S.; Han, J.Y. Wnt/beta-catenin signaling pathway activation is required for proliferation of chicken primordial germ cells in vitro. Sci. Rep. 2016, 6, 34510. [Google Scholar] [CrossRef] [Green Version]
- Whyte, J.; Glover, J.D.; Woodcock, M.; Brzeszczynska, J.; Taylor, L.; Sherman, A.; Kaiser, P.; McGrew, M.J. FGF, Insulin, and SMAD Signaling Cooperate for Avian Primordial Germ Cell Self-Renewal. Stem Cell Rep. 2015, 5, 1171–1182. [Google Scholar] [CrossRef] [Green Version]
- Yakhkeshi, S.; Rahimi, S.; Sharafi, M.; Hassani, S.N.; Taleahmad, S.; Shahverdi, A.; Baharvand, H. In vitro improvement of quail primordial germ cell expansion through activation of TGF-beta signaling pathway. J. Cell. Biochem. 2018, 119, 4309–4319. [Google Scholar] [CrossRef]
- Jung, K.M.; Kim, Y.M.; Keyte, A.L.; Biegler, M.T.; Rengaraj, D.; Lee, H.J.; Mello, C.V.; Velho, T.A.F.; Fedrigo, O.; Haase, B.; et al. Identification and characterization of primordial germ cells in a vocal learning Neoaves species, the zebra finch. FASEB J. 2019, 33, 13825–13836. [Google Scholar] [CrossRef] [Green Version]
- Park, T.S.; Han, J.Y. piggyBac transposition into primordial germ cells is an efficient tool for transgenesis in chickens. Proc. Natl. Acad. Sci. USA 2012, 109, 9337–9341. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, J.; Taylor, L.; Sherman, A.; Kawakami, K.; Takahashi, Y.; Sang, H.M.; McGrew, M.J. Efficient genetic modification and germ-line transmission of primordial germ cells using piggyBac and Tol2 transposons. Proc. Natl. Acad. Sci. USA 2012, 109, E1466–E1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Lee, H.C.; Kim, Y.M.; Hwang, Y.S.; Park, Y.H.; Park, T.S.; Han, J.Y. Site-specific recombination in the chicken genome using Flipase recombinase-mediated cassette exchange. FASEB J. 2016, 30, 555–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivics, Z.; Li, M.A.; Mates, L.; Boeke, J.D.; Nagy, A.; Bradley, A.; Izsvak, Z. Transposon-mediated genome manipulation in vertebrates. Nat. Methods 2009, 6, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronovich, E.L.; Bell, J.B.; Belur, L.R.; Gunther, R.; Koniar, B.; Erickson, D.C.; Schachern, P.A.; Matise, I.; McIvor, R.S.; Whitley, C.B.; et al. Prolonged expression of a lysosomal enzyme in mouse liver after Sleeping Beauty transposon-mediated gene delivery: Implications for non-viral gene therapy of mucopolysaccharidoses. J. Gene Med. 2007, 9, 403–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, T.S.; Lee, H.G.; Moon, J.K.; Lee, H.J.; Yoon, J.W.; Yun, B.N.; Kang, S.C.; Kim, J.; Kim, H.; Han, J.Y.; et al. Deposition of bioactive human epidermal growth factor in the egg white of transgenic hens using an oviduct-specific minisynthetic promoter. FASEB J. 2015, 29, 2386–2396. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.M.; Park, J.S.; Kim, S.K.; Jung, K.M.; Hwang, Y.S.; Han, M.; Lee, H.J.; Seo, H.W.; Suh, J.Y.; Han, B.K.; et al. The transgenic chicken derived anti-CD20 monoclonal antibodies exhibits greater anti-cancer therapeutic potential with enhanced Fc effector functions. Biomaterials 2018, 167, 58–68. [Google Scholar] [CrossRef]
- Reynaud, G. Reproductive capacity and offspring of chickens submitted to a transfer of primordial germ cells during embryonic life. Wilhelm Roux’s Arch. Dev. Biol. 1976, 179, 85–110. [Google Scholar] [CrossRef]
- Wentworth, B.C.; Tsai, H.; Hallett, J.H.; Gonzales, D.S.; Rajcic-Spasojevic, G. Manipulation of avian primordial germ cells and gonadal differentiation. Poult. Sci. 1989, 68, 999–1010. [Google Scholar] [CrossRef]
- Vick, L.; Li, Y.; Simkiss, K. Transgenic birds from transformed primordial germ cells. Proc. Biol. Sci. 1993, 251, 179–182. [Google Scholar] [CrossRef]
- Yasuda, Y.; Tajima, A.; Fujimoto, T.; Kuwana, T. A method to obtain avian germ-line chimaeras using isolated primordial germ cells. J. Reprod. Fertil. 1992, 96, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Naito, M.; Tajima, A.; Tagami, T.; Yasuda, Y.; Kuwana, T. Preservation of chick primordial germ cells in liquid nitrogen and subsequent production of viable offspring. J. Reprod. Fertil. 1994, 102, 321–325. [Google Scholar] [CrossRef] [Green Version]
- Tajima, A.; Naito, M.; Yasuda, Y.; Kuwana, T. Production of germ-line chimeras by transfer of cryopreserved gonadal primordial germ cells (gPGCs) in chicken. J. Exp. Zool. 1998, 280, 265–267. [Google Scholar] [CrossRef]
- Trefil, P.; Aumann, D.; Kosolová, A.; Mucksová, J.; Benešová, B.; Kallina, J.; Wurmser, C.; Fries, R.; Elleder, D.; Schsser, B.; et al. Male fertility restored by transplanting primordial germ cells into testes: A new way towards efficient transgenesis in chicken. Sci. Rep. 2017, 7, 14246. [Google Scholar] [CrossRef]
- Carsience, R.S.; Clark, M.E.; Verrinder Gibbins, A.M.; Etches, R.J. Germline chimeric chickens from dispersed donor blastodermal cells and compromised recipient embryos. Development 1993, 117, 669–675. [Google Scholar] [CrossRef]
- Nakamura, Y.; Usui, F.; Ono, T.; Takeda, K.; Nirasawa, K.; Kagami, H.; Tagami, T. Germline replacement by transfer of primordial germ cells into partially sterilized embryos in the chicken. Biol. Reprod. 2010, 83, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y. Poultry genetic resource conservation using primordial germ cells. J. Reprod. Dev. 2016, 62, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.M.; Park, K.J.; Park, J.S.; Jung, K.M.; Han, J.Y. In vivo enrichment of busulfan-resistant germ cells for efficient production of transgenic avian models. Sci. Rep. 2021, 11, 9127. [Google Scholar] [CrossRef]
- Jung, K.M.; Kim, Y.M.; Han, J.Y. Transplantation and enrichment of busulfan-resistant primordial germ cells into adult testes for efficient production of germline chimeras in songbirds. Biol. Reprod. 2022, 108, 316–323. [Google Scholar] [CrossRef]
- Taylor, L.; Carlson, D.F.; Nandi, S.; Sherman, A.; Fahrenkrug, S.C.; McGrew, M.J. Efficient TALEN-mediated gene targeting of chicken primordial germ cells. Development 2017, 144, 928–934. [Google Scholar] [CrossRef] [Green Version]
- Ballantyne, M.; Woodcock, M.; Doddamani, D.; Hu, T.; Taylor, L.; Hawken, R.J.; McGrew, M.J. Direct allele introgression into pure chicken breeds using Sire Dam Surrogate (SDS) mating. Nat. Commun. 2021, 12, 659. [Google Scholar] [CrossRef]
- Rogers, C.S. Engineering Large Animal Species to Model Human Diseases. Curr. Protoc. Hum. Genet. 2016, 90, 15.9.1–15.9.14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, K.R.; Capecchi, M.R. Site-directed mutagenesis by gene targeting in mouse embryo-derived stem cells. Cell 1987, 51, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.; Kolber-Simonds, D.; Park, K.W.; Cheong, H.T.; Greenstein, J.L.; Im, G.S.; Samuel, M.; Bonk, A.; Rieke, A.; Day, B.N.; et al. Production of alpha-1,3-galactosyltransferase knockout pigs by nuclear transfer cloning. Science 2002, 295, 1089–1092. [Google Scholar] [CrossRef] [PubMed]
- Nottle, M.B.; Beebe, L.F.; Harrison, S.J.; McIlfatrick, S.M.; Ashman, R.J.; O’Connell, P.J.; Salvaris, E.J.; Fisicaro, N.; Pommey, S.; Cowan, P.J.; et al. Production of homozygous alpha-1,3-galactosyltransferase knockout pigs by breeding and somatic cell nuclear transfer. Xenotransplantation 2007, 14, 339–344. [Google Scholar] [CrossRef]
- Petitte, J.N.; Liu, G.; Yang, Z. Avian pluripotent stem cells. Mech. Dev. 2004, 121, 1159–1168. [Google Scholar] [CrossRef]
- Petitte, J.N.; Clark, M.E.; Liu, G.; Verrinder Gibbins, A.M.; Etches, R.J. Production of somatic and germline chimeras in the chicken by transfer of early blastodermal cells. Development 1990, 108, 185–189. [Google Scholar] [CrossRef]
- Pain, B.; Clark, M.E.; Shen, M.; Nakazawa, H.; Sakurai, M.; Samarut, J.; Etches, R.J. Long-term in vitro culture and characterisation of avian embryonic stem cells with multiple morphogenetic potentialities. Development 1996, 122, 2339–2348. [Google Scholar] [CrossRef]
- Van de Lavoir, M.C.; Mather-Love, C.; Leighton, P.; Diamond, J.H.; Heyer, B.S.; Roberts, R.; Zhu, L.; Winters-Digiacinto, P.; Kerchner, A.; Gessaro, T.; et al. High-grade transgenic somatic chimeras from chicken embryonic stem cells. Mech. Dev. 2006, 123, 31–41. [Google Scholar] [CrossRef]
- Lavial, F.; Acloque, H.; Bachelard, E.; Nieto, M.A.; Samarut, J.; Pain, B. Ectopic expression of Cvh (Chicken Vasa homologue) mediates the reprogramming of chicken embryonic stem cells to a germ cell fate. Dev. Biol. 2009, 330, 73–82. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Ji, G.; Mao, J.; Liu, M.; Wang, L.; Chen, C.; Liu, L. Generation of porcine-induced pluripotent stem cells by using OCT4 and KLF4 porcine factors. Cell. Reprogram. 2012, 14, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Zhong, B.; Trobridge, G.D.; Zhang, X.; Watts, K.L.; Ramakrishnan, A.; Wohlfahrt, M.; Adair, J.E.; Kiem, H.P. Efficient generation of nonhuman primate induced pluripotent stem cells. Stem Cells Dev. 2011, 20, 795–807. [Google Scholar] [CrossRef] [Green Version]
- Rossello, R.A.; Chen, C.C.; Dai, R.; Howard, J.T.; Hochgeschwender, U.; Jarvis, E.D. Mammalian genes induce partially reprogrammed pluripotent stem cells in non-mammalian vertebrate and invertebrate species. Elife 2013, 2, e00036. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; West, F.D.; Jordan, B.J.; Mumaw, J.L.; Jordan, E.T.; Gallegos-Cardenas, A.; Beckstead, R.B.; Stice, S.L. Avian-induced pluripotent stem cells derived using human reprogramming factors. Stem Cells Dev. 2012, 21, 394–403. [Google Scholar] [CrossRef]
- Kim, Y.M.; Park, Y.H.; Lim, J.M.; Jung, H.; Han, J.Y. Technical note: Induction of pluripotent stem cell-like cells from chicken feather follicle cells. J. Anim. Sci. 2017, 95, 3479–3486. [Google Scholar] [CrossRef]
- Zhao, R.; Zuo, Q.; Yuan, X.; Jin, K.; Jin, J.; Ding, Y.; Zhang, C.; Li, T.; Jiang, J.; Li, J.; et al. Production of viable chicken by allogeneic transplantation of primordial germ cells induced from somatic cells. Nat. Commun. 2021, 12, 2989. [Google Scholar] [CrossRef]
- Ryu, B.Y.; Orwig, K.E.; Oatley, J.M.; Lin, C.C.; Chang, L.J.; Avarbock, M.R.; Brinster, R.L. Efficient generation of transgenic rats through the male germline using lentiviral transduction and transplantation of spermatogonial stem cells. J. Androl. 2007, 28, 353–360. [Google Scholar] [CrossRef]
- Hamra, F.K.; Gatlin, J.; Chapman, K.M.; Grellhesl, D.M.; Garcia, J.V.; Hammer, R.E.; Garbers, D.L. Production of transgenic rats by lentiviral transduction of male germ-line stem cells. Proc. Natl. Acad. Sci. USA 2002, 99, 14931–14936. [Google Scholar] [CrossRef] [Green Version]
- Kanatsu-Shinohara, M.; Kato, M.; Takehashi, M.; Morimoto, H.; Takashima, S.; Chuma, S.; Nakatsuji, N.; Hirabayashi, M.; Shinohara, T. Production of transgenic rats via lentiviral transduction and xenogeneic transplantation of spermatogonial stem cells. Biol. Reprod. 2008, 79, 1121–1128. [Google Scholar] [CrossRef]
- Li, C.H.; Yan, L.Z.; Ban, W.Z.; Tu, Q.; Wu, Y.; Wang, L.; Bi, R.; Ji, S.; Ma, Y.H.; Nie, W.H.; et al. Long-term propagation of tree shrew spermatogonial stem cells in culture and successful generation of transgenic offspring. Cell Res. 2017, 27, 241–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanatsu-Shinohara, M.; Kato-Itoh, M.; Ikawa, M.; Takehashi, M.; Sanbo, M.; Morioka, Y.; Tanaka, T.; Morimoto, H.; Hirabayashi, M.; Shinohara, T. Homologous recombination in rat germline stem cells. Biol. Reprod. 2011, 85, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Izsvak, Z.; Frohlich, J.; Grabundzija, I.; Shirley, J.R.; Powell, H.M.; Chapman, K.M.; Ivics, Z.; Hamra, F.K. Generating knockout rats by transposon mutagenesis in spermatogonial stem cells. Nat. Methods 2010, 7, 443–445. [Google Scholar] [CrossRef] [PubMed]
- Tegelenbosch, R.A.; de Rooij, D.G. A quantitative study of spermatogonial multiplication and stem cell renewal in the C3H/101 F1 hybrid mouse. Mutat. Res./Fundam. Mol. Mech. Mutagen. 1993, 290, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Kossack, N.; Meneses, J.; Shefi, S.; Nguyen, H.N.; Chavez, S.; Nicholas, C.; Gromoll, J.; Turek, P.J.; Reijo-Pera, R.A. Isolation and characterization of pluripotent human spermatogonial stem cell-derived cells. Stem Cells 2009, 27, 138–149. [Google Scholar] [CrossRef] [Green Version]
- Nagano, M.; Ryu, B.Y.; Brinster, C.J.; Avarbock, M.R.; Brinster, R.L. Maintenance of mouse male germ line stem cells in vitro. Biol. Reprod. 2003, 68, 2207–2214. [Google Scholar] [CrossRef]
- Nasiri, Z.; Hosseini, S.M.; Hajian, M.; Abedi, P.; Bahadorani, M.; Baharvand, H.; Nasr-Esfahani, M.H. Effects of different feeder layers on short-term culture of prepubertal bovine testicular germ cells in-vitro. Theriogenology 2012, 77, 1519–1528. [Google Scholar] [CrossRef]
- Kuijk, E.W.; Colenbrander, B.; Roelen, B.A. The effects of growth factors on in vitro-cultured porcine testicular cells. Reproduction 2009, 138, 721–731. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.G.; Lee, Y.M.; Park, T.S.; Park, S.H.; Lim, J.M.; Han, J.Y. Identification, culture, and characterization of germline stem cell-like cells in chicken testes. Biol. Reprod. 2007, 76, 173–182. [Google Scholar] [CrossRef]
- Pramod, R.K.; Lee, B.R.; Kim, Y.M.; Lee, H.J.; Park, Y.H.; Ono, T.; Lim, J.M.; Han, J.Y. Isolation, Characterization, and In Vitro Culturing of Spermatogonial Stem Cells in Japanese Quail (Coturnix japonica). Stem Cells Dev. 2017, 26, 60–70. [Google Scholar] [CrossRef]
- Zhang, Y.; Sastre, D.; Wang, F. CRISPR/Cas9 Genome Editing: A Promising Tool for Therapeutic Applications of Induced Pluripotent Stem Cells. Curr. Stem Cell Res. Ther. 2018, 13, 243–251. [Google Scholar] [CrossRef]
- De Silva Feelixge, H.S.; Stone, D.; Roychoudhury, P.; Aubert, M.; Jerome, K.R. CRISPR/Cas9 and Genome Editing for Viral Disease-Is Resistance Futile? ACS Infect. Dis. 2018, 4, 871–880. [Google Scholar] [CrossRef]
- Ingusci, S.; Verlengia, G.; Soukupova, M.; Zucchini, S.; Simonato, M. Gene Therapy Tools for Brain Diseases. Front. Pharmacol. 2019, 10, 724. [Google Scholar] [CrossRef]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR-Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef]
- Schusser, B.; Collarini, E.J.; Yi, H.; Izquierdo, S.M.; Fesler, J.; Pedersen, D.; Klasing, K.C.; Kaspers, B.; Harriman, W.D.; van de Lavoir, M.C.; et al. Immunoglobulin knockout chickens via efficient homologous recombination in primordial germ cells. Proc. Natl. Acad. Sci. USA 2013, 110, 20170–20175. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, J.S. A guide to genome engineering with programmable nucleases. Nat. Rev. Genet. 2014, 15, 321–334. [Google Scholar] [CrossRef]
- Li, H.; Yang, Y.; Hong, W.; Huang, M.; Wu, M.; Zhao, X. Applications of genome editing technology in the targeted therapy of human diseases: Mechanisms, advances and prospects. Signal Transduct. Target. Ther. 2020, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., 3rd. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Park, T.S.; Lee, H.J.; Kim, K.H.; Kim, J.S.; Han, J.Y. Targeted gene knockout in chickens mediated by TALENs. Proc. Natl. Acad. Sci. USA 2014, 111, 12716–12721. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.W.; Kim, S.; Kim, J.M.; Kim, J.S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.H.; Kim, J.M.; Kim, H.T.; Lee, J.; Jeon, J.; Jin, Y.; Choi, J.H.; Ban, Y.H.; Ha, S.J.; Kim, C.H.; et al. Highly efficient gene knockout in mice and zebrafish with RNA-guided endonucleases. Genome Res. 2014, 24, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qiu, J.L.; et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Tsai, S.Q.; Sander, J.D.; Peterson, R.T.; Yeh, J.R.; Joung, J.K. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 227–229. [Google Scholar] [CrossRef] [Green Version]
- Oishi, I.; Yoshii, K.; Miyahara, D.; Kagami, H.; Tagami, T. Targeted mutagenesis in chicken using CRISPR/Cas9 system. Sci. Rep. 2016, 6, 23980. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, L.; Pedersen, D.; Ching, K.H.; Yi, H.; Collarini, E.J.; Izquierdo, S.; van de Lavoir, M.C.; Leighton, P.A. Germline Gene Editing in Chickens by Efficient CRISPR-Mediated Homologous Recombination in Primordial Germ Cells. PLoS ONE 2016, 11, e0154303. [Google Scholar] [CrossRef]
- Park, T.S.; Park, J.; Lee, J.H.; Park, J.W.; Park, B.C. Disruption of G0/G1 switch gene 2 (G0S2) reduced abdominal fat deposition and altered fatty acid composition in chicken. FASEB J. 2019, 33, 1188–1198. [Google Scholar] [CrossRef]
- Kim, G.D.; Lee, J.H.; Song, S.; Kim, S.W.; Han, J.S.; Shin, S.P.; Park, B.C.; Park, T.S. Generation of myostatin-knockout chickens mediated by D10A-Cas9 nickase. FASEB J. 2020, 34, 5688–5696. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kim, D.H.; Lee, K. Muscle Hyperplasia in Japanese Quail by Single Amino Acid Deletion in MSTN Propeptide. Int. J. Mol. Sci. 2020, 21, 1504. [Google Scholar] [CrossRef] [Green Version]
- Rieblinger, B.; Sid, H.; Duda, D.; Bozoglu, T.; Klinger, R.; Schlickenrieder, A.; Lengyel, K.; Flisikowski, K.; Flisikowska, T.; Simm, N.; et al. Cas9-expressing chickens and pigs as resources for genome editing in livestock. Proc. Natl. Acad. Sci. USA 2021, 118, e2022562118. [Google Scholar] [CrossRef]
- Challagulla, A.; Jenkins, K.A.; O’Neil, T.E.; Morris, K.R.; Wise, T.G.; Tizard, M.L.; Bean, A.G.D.; Schat, K.A.; Doran, T.J. Germline engineering of the chicken genome using CRISPR/Cas9 by in vitro transfection of PGCs. Anim. Biotechnol. 2020, 1–10. [Google Scholar] [CrossRef]
- Komor, A.C.; Badran, A.H.; Liu, D.R. Editing the Genome Without Double-Stranded DNA Breaks. ACS Chem. Biol. 2018, 13, 383–388. [Google Scholar] [CrossRef]
- Lee, K.Y.; Lee, H.J.; Choi, H.J.; Han, S.T.; Lee, K.H.; Park, K.J.; Park, J.S.; Jung, K.M.; Kim, Y.M.; Han, H.J.; et al. Highly elevated base excision repair pathway in primordial germ cells causes low base editing activity in chickens. FASEB J. 2020, 34, 15907–15921. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Atsuta, Y.; Suzuki, K.; Iikawa, H.; Yaguchi, H.; Saito, D. Prime editing in chicken fibroblasts and primordial germ cells. Dev. Growth Differ. 2022, 64, 548–557. [Google Scholar] [CrossRef]
- Koslová, A.; Trefil, P.; Mucksová, J.; Reinišová, M.; Plachý, J.; Kalina, J.; Kučerová, D.; Geryk, J.; Krchlíková, V.; Lejčková, B.; et al. Precise CRISPR/Cas9 editing of the NHE1 gene renders chickens resistant to the J subgroup of avian leukosis virus. Proc. Natl. Acad. Sci. USA 2020, 117, 2108–2112. [Google Scholar] [CrossRef]
- Park, Y.H.; Chungu, K.; Lee, S.B.; Woo, S.J.; Cho, H.Y.; Lee, H.J.; Rengaraj, D.; Lee, J.H.; Song, C.S.; Lim, J.M.; et al. Host-Specific Restriction of Avian Influenza Virus Caused by Differential Dynamics of ANP32 Family Members. J. Infect. Dis. 2020, 221, 71–80. [Google Scholar] [CrossRef]
- Hagag, I.T.; Wight, D.J.; Bartsch, D.; Sid, H.; Jordan, I.; Bertzbach, L.D.; Schusser, B.; Kaufer, B.B. Abrogation of Marek’s disease virus replication using CRISPR/Cas9. Sci. Rep. 2020, 10, 10919. [Google Scholar] [CrossRef] [PubMed]
- Fadly, A.M. Isolation and identification of avian leukosis viruses: A review. Avian Pathol. 2000, 29, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, K.Y.; Jung, K.M.; Park, K.J.; Lee, K.O.; Suh, J.Y.; Yao, Y.; Nair, V.; Han, J.Y. Precise gene editing of chicken Na+/H+ exchange type 1 (chNHE1) confers resistance to avian leukosis virus subgroup J (ALV-J). Dev. Comp. Immunol. 2017, 77, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, K.Y.; Park, Y.H.; Choi, H.J.; Yao, Y.; Nair, V.; Han, J.Y. Acquisition of resistance to avian leukosis virus subgroup B through mutations on tvb cysteine-rich domains in DF-1 chicken fibroblasts. Vet. Res. 2017, 48, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Park, K.J.; Lee, K.Y.; Yao, Y.; Nair, V.; Han, J.Y. Sequential disruption of ALV host receptor genes reveals no sharing of receptors between ALV subgroups A, B, and J. J. Anim. Sci. Biotechnol. 2019, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- Koslova, A.; Kucerova, D.; Reinisova, M.; Geryk, J.; Trefil, P.; Hejnar, J. Genetic Resistance to Avian Leukosis Viruses Induced by CRISPR/Cas9 Editing of Specific Receptor Genes in Chicken Cells. Viruses 2018, 10, 605. [Google Scholar] [CrossRef] [Green Version]
- Hellmich, R.; Sid, H.; Lengyel, K.; Flisikowski, K.; Schlickenrieder, A.; Bartsch, D.; Thoma, T.; Bertzbach, L.D.; Kaufer, B.B.; Nair, V.; et al. Acquiring Resistance Against a Retroviral Infection via CRISPR/Cas9 Targeted Genome Editing in a Commercial Chicken Line. Front. Genome. Ed. 2020, 2, 3. [Google Scholar] [CrossRef]
- Long, J.S.; Giotis, E.S.; Moncorge, O.; Frise, R.; Mistry, B.; James, J.; Morisson, M.; Iqbal, M.; Vignal, A.; Skinner, M.A.; et al. Species difference in ANP32A underlies influenza A virus polymerase host restriction. Nature 2016, 529, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Long, J.S.; Idoko-Akoh, A.; Mistry, B.; Goldhill, D.; Staller, E.; Schreyer, J.; Ross, C.; Goodbourn, S.; Shelton, H.; Skinner, M.A.; et al. Species specific differences in use of ANP32 proteins by influenza A virus. Elife 2019, 8, e45066. [Google Scholar] [CrossRef]
- Park, Y.H.; Woo, S.J.; Chungu, K.; Lee, S.B.; Shim, J.H.; Lee, H.J.; Kim, I.; Rengaraj, D.; Song, C.S.; Suh, J.Y.; et al. Asp149 and Asp152 in chicken and human ANP32A play an essential role in the interaction with influenza viral polymerase. FASEB J. 2021, 35, e21630. [Google Scholar] [CrossRef]
- Barber, M.R.W.; Aldridge, J.R., Jr.; Webster, R.G.; Magor, K.E. Association of RIG-I with innate immunity of ducks to influenza. Proc. Natl. Acad. Sci. USA 2010, 107, 5913–5918. [Google Scholar] [CrossRef] [Green Version]
- Ichikawa, K.; Motoe, Y.; Ezaki, R.; Matsuzaki, M.; Horiuchi, H. Knock-in of the duck retinoic acid-inducible gene I (RIG-I) into the Mx gene in DF-1 cells enables both stable and immune response-dependent RIG-I expression. Biochem. Biophys. Rep. 2021, 27, 101084. [Google Scholar] [CrossRef]
- Woo, S.J.; Choi, H.J.; Park, Y.H.; Rengaraj, D.; Kim, J.K.; Han, J.Y. Amplification of immunity by engineering chicken MDA5 combined with the C terminal domain (CTD) of RIG-I. Appl. Microbiol. Biotechnol. 2022, 106, 1599–1613. [Google Scholar] [CrossRef]
- Challagulla, A.; Jenkins, K.A.; O’Neil, T.E.; Shi, S.; Morris, K.R.; Wise, T.G.; Paradkar, P.N.; Tizard, M.L.; Doran, T.J.; Schat, K.A. In Vivo Inhibition of Marke’s Disease Virus in Transgenic Chickens Expressing Cas9 and gRNA against ICP4. Microorganisms 2021, 9, 164. [Google Scholar] [CrossRef]
- Ivarie, R. Avian transgenesis: Progress towards the promise. Trends Biotechnol. 2003, 21, 14–19. [Google Scholar] [CrossRef]
- Han, J.Y. Germ cells and transgenesis in chickens. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 61–80. [Google Scholar] [CrossRef]
- Sang, H.M. Transgenics, chickens and therapeutic proteins. Vox Sang. 2004, 87 (Suppl. 2), 164–166. [Google Scholar] [CrossRef]
- Zhu, L.; van de Lavoir, M.C.; Albanese, J.; Beenhouwer, D.O.; Cardarelli, P.M.; Cuison, S.; Deng, D.F.; Deshpande, S.; Diamond, J.H.; Green, L.; et al. Production of human monoclonal antibody in eggs of chimeric chickens. Nat. Biotechnol. 2005, 23, 1159–1169. [Google Scholar] [CrossRef]
- Lillico, S.; Sherman, A.; McGrew, M.; Robertson, C.; Smith, J.; Haslam, C.; Barnard, P.; Radcliffe, P.; Mitrophanous, K.; Elliot, E.; et al. Oviduct-specific expression of two therapeutic proteins in transgenic hens. Proc. Natl. Acad. Sci. USA 2007, 104, 1771–1776. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.; Wu, H.; Li, Q.; Sun, Y.; Liu, T.; Fei, J.; Zhao, Y.; Wu, S.; Hu, X.; Li, N. Expression of recombinant human lysozyme in egg whites of transgenic hens. PLoS ONE 2015, 10, e0118626. [Google Scholar] [CrossRef]
- Oishi, I.; Yoshii, K.; Miyahara, D.; Tagami, T. Efficient production of human interferon beta in the white of eggs from ovalbumin gene-targeted hens. Sci. Rep. 2018, 8, 10203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.M.; Park, J.S.; Choi, H.J.; Jung, K.M.; Lee, K.Y.; Shim, J.H.; Park, K.J.; Han, J.Y. Efficient production of recombinant human adiponectin in egg white using genome edited chickens. Front. Nutr. 2022, 9, 1068558. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Shim, J.H.; Park, J.S.; Choi, H.J.; Jung, K.M.; Lee, K.Y.; Park, K.J.; Han, J.Y. Sequential verification of exogenous protein production in OVA gene-targeted chicken bioreactors. Poult. Sci. 2023, 102, 102247. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, C. FDA approves ‘farmaceutical’ drug from transgenic chickens. Nat. Biotechnol. 2016, 34, 117–119. [Google Scholar] [CrossRef]
- Sly, W.S.; Vogler, C.; Grubb, J.H.; Levy, B.; Galvin, N.; Tan, Y.; Nishioka, T.; Tomatsu, S. Enzyme therapy in mannose receptor-null mucopolysaccharidosis VII mice defines roles for the mannose 6-phosphate and mannose receptors. Proc. Natl. Acad. Sci. USA 2006, 103, 15172–15177. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, E.F. Enzyme replacement therapy-a brief history. In Fabry Disease: Perspectives from 5 Years of FOS; Mehta, A., Beck, M., Sunder-Plassmann, G., Eds.; Oxford PharmaGenesis: Oxford, UK, 2006. [Google Scholar]
- Hintze, S.; Limmer, S.; Dabrowska-Schlepp, P.; Berg, B.; Krieghoff, N.; Busch, A.; Schaaf, A.; Meinke, P.; Schoser, B. Moss-Derived Human Recombinant GAA Provides an Optimized Enzyme Uptake in Differentiated Human Muscle Cells of Pompe Disease. Int. J. Mol. Sci. 2020, 21, 2642. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.W.; Huy, N.X.; Kim, H.B.; Kim, N.S.; van Giap, D.; Yang, M.S. Production of recombinant human acid alpha-glucosidase with high-mannose glycans in gnt1 rice for the treatment of Pompe disease. J. Biotechnol. 2017, 249, 42–50. [Google Scholar] [CrossRef]
- Shields, R.L.; Lai, J.; Keck, R.; O’Connell, L.Y.; Hong, K.; Meng, Y.G.; Weikert, S.H.; Presta, L.G. Lack of fucose on human IgG1 N-linked oligosaccharide improves binding to human Fcgamma RIII and antibody-dependent cellular toxicity. J. Biol. Chem. 2002, 277, 26733–26740. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.Y.; Choi, H.J.; Park, K.J.; Woo, S.J.; Kim, Y.M.; Han, J.Y. Development and characterization of a CRISPR/Cas9-mediated RAG1 knockout chicken model lacking mature B and T cells. Front. Immunol. 2022, 13, 892476. [Google Scholar] [CrossRef]
- Johnson, P.A.; Giles, J.R. The hen as a model of ovarian cancer. Nat. Rev. Cancer 2013, 13, 432–436. [Google Scholar] [CrossRef]
- Rengaraj, D.; Cha, D.G.; Lee, H.J.; Lee, K.Y.; Choi, Y.H.; Jung, K.M.; Kim, Y.M.; Choi, H.J.; Choi, H.J.; Yoo, E.; et al. Dissecting chicken germ cell dynamics by combining a germ cell tracing transgenic chicken model with single-cell RNA sequencing. Comput. Struct. Biotechnol. J. 2022, 20, 1654–1669. [Google Scholar] [CrossRef]
- Hagihara, Y.; Qkuzaki, Y.; Matsubayashi, K.; Kaneoka, H.; Suzuki, T.; Iijima, S.; Nishijima, K.-I. Primordial germ cell-specific expression of eGFP in transgenic chickens. Genesis 2020, 58, e23388. [Google Scholar] [CrossRef]
- Lee, H.J.; Seo, M.; Choi, H.J.; Rengaraj, D.; Jung, K.M.; Park, J.S.; Lee, K.Y.; Kim, Y.M.; Park, K.J.; Han, S.T.; et al. DMRT1 gene disruption alone induces incomplete gonad feminization in chicken. FASEB J. 2021, 35, e21876. [Google Scholar] [CrossRef]
- Ioannidis, J.; Taylor, G.; Zhao, D.; Liu, L.; Idoko-Akoh, A.; Gong, D.; Lovell-Badge, R.; Guioli, S.; McGrew, M.J.; Clinton, M. Primary sex determination in birds depends on DMRT1 dosage, but gonadal sex does not determine adult secondary sex characteristics. Proc. Natl. Acad. Sci. USA 2021, 118, e2020909118. [Google Scholar] [CrossRef]
- Lai, C.S.; Fisher, S.E.; Hurst, J.A.; Vargha-Khadem, F.; Monaco, A.P. A forkhead-domain gene is mutated in a severe speech and language disorder. Nature 2001, 413, 519–523. [Google Scholar] [CrossRef]
- Haesler, S.; Rochefort, C.; Georgi, B.; Licznerski, P.; Osten, P.; Scharff, C. Incomplete and inaccurate vocal imitation after knockdown of FoxP2 in songbird basal ganglia nucleus Area X. PLoS Biol. 2007, 5, e321. [Google Scholar] [CrossRef] [Green Version]
- Scott, B.B.; Velho, T.A.; Sim, S.; Lois, C. Applications of avian transgenesis. ILAR J. 2010, 51, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.M.; Seo, M.; Kim, Y.M.; Kim, J.L.; Han, J.Y. Single-Cell RNA Sequencing Revealed the Heterogeneity of Gonadal Primordial Germ Cells in Zebra Finch (Taeniopygia guttata). Front. Cell Dev. Biol. 2021, 9, 791335. [Google Scholar] [CrossRef]
- Jung, K.M.; Seo, M.; Han, J.Y. Comparative single-cell transcriptomic analysis reveals differences in signaling pathways in gonadal primordial germ cells between chicken (Gallus gallus) and zebra finch (Taeniopygia guttata). FASEB J. 2023, 37, e22706. [Google Scholar] [CrossRef]
- Jung, K.M.; Kim, Y.M.; Yoo, E.; Han, J.Y. Generation and characterization of genome-modified chondrocyte-like cells from the zebra finch cell line immortalized by c-MYC expression. Front. Zool. 2022, 19, 18. [Google Scholar] [CrossRef]
- Wray-Cahen, D.; Bodnar, A.; Rexroad, C.; Siewerdt, F.; Kovich, D. Advancing genome editing to improve the sustainability and resiliency of animal agriculture. CABI Agric. Biosci. 2022, 3, 21. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-M.; Woo, S.-J.; Han, J.-Y. Strategies for the Generation of Gene Modified Avian Models: Advancement in Avian Germline Transmission, Genome Editing, and Applications. Genes 2023, 14, 899. https://doi.org/10.3390/genes14040899
Kim Y-M, Woo S-J, Han J-Y. Strategies for the Generation of Gene Modified Avian Models: Advancement in Avian Germline Transmission, Genome Editing, and Applications. Genes. 2023; 14(4):899. https://doi.org/10.3390/genes14040899
Chicago/Turabian StyleKim, Young-Min, Seung-Je Woo, and Jae-Yong Han. 2023. "Strategies for the Generation of Gene Modified Avian Models: Advancement in Avian Germline Transmission, Genome Editing, and Applications" Genes 14, no. 4: 899. https://doi.org/10.3390/genes14040899