Identification and Functional Analysis of ToBPI1/LBP and ToBPI2/LBP in Anti-Bacterial Infection of Trachinotus ovatus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Bacterial Strain
2.2. Fish Challenge Experiments and Tissue Sample
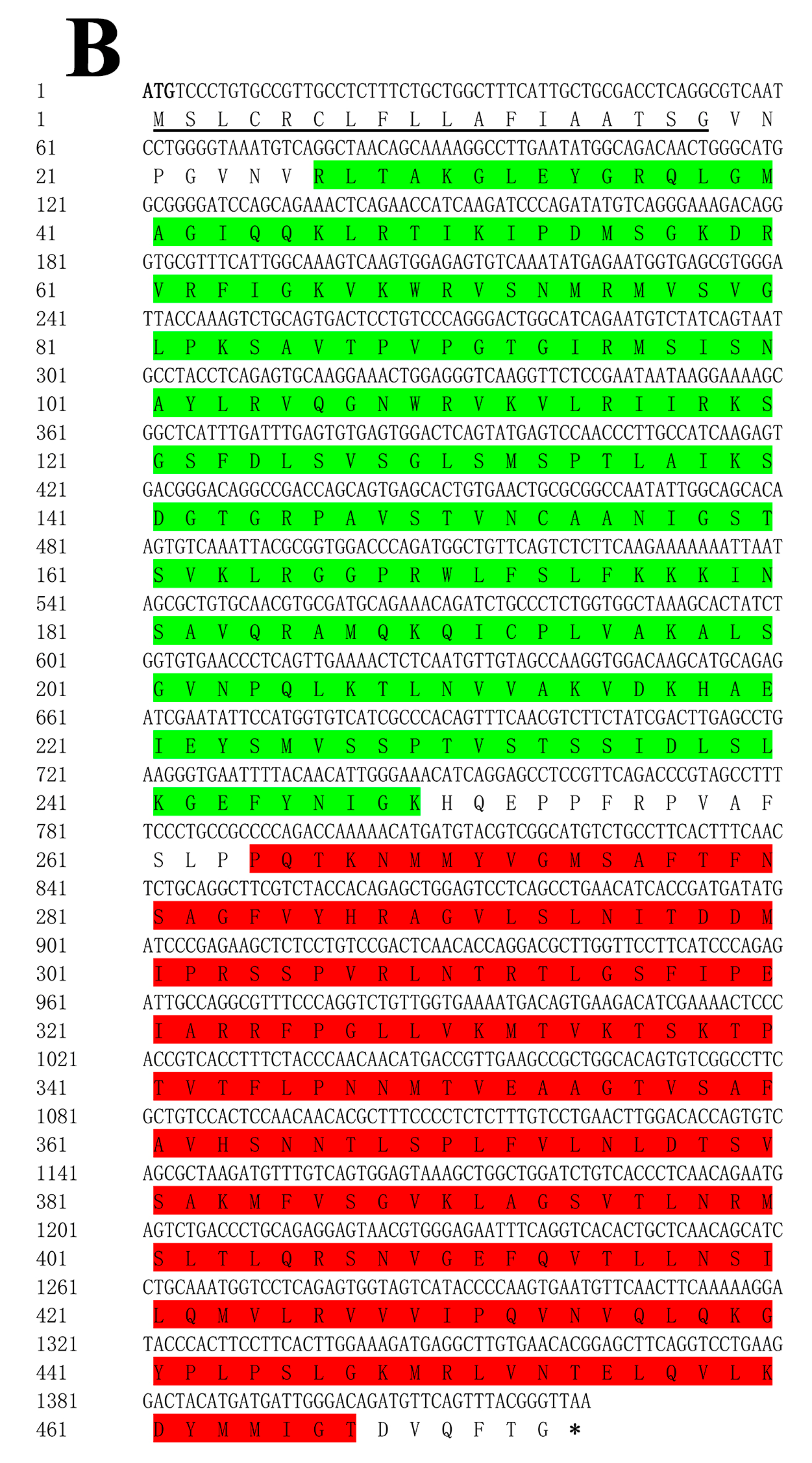
2.3. Gene Cloning
2.4. Bioinformatics Analysis
2.5. Expression of ToBPI1/LBP and ToBPI2/LBP in Healthy and Challenged T. ovatus
2.6. Prokaryotic Expression, Purification and Western Blotting of the Recombinant ToBPI1/LBP(rToBPI1/LBP) and ToBPI2/LBP(rToBPI2/LBP)
2.7. Antimicrobial Activity
2.8. Cell Membrane Permeability Assay
2.9. Statistical Analyses
3. Results
3.1. ToBPI1/LBP and ToBPI2/LBP Sequence Analysis
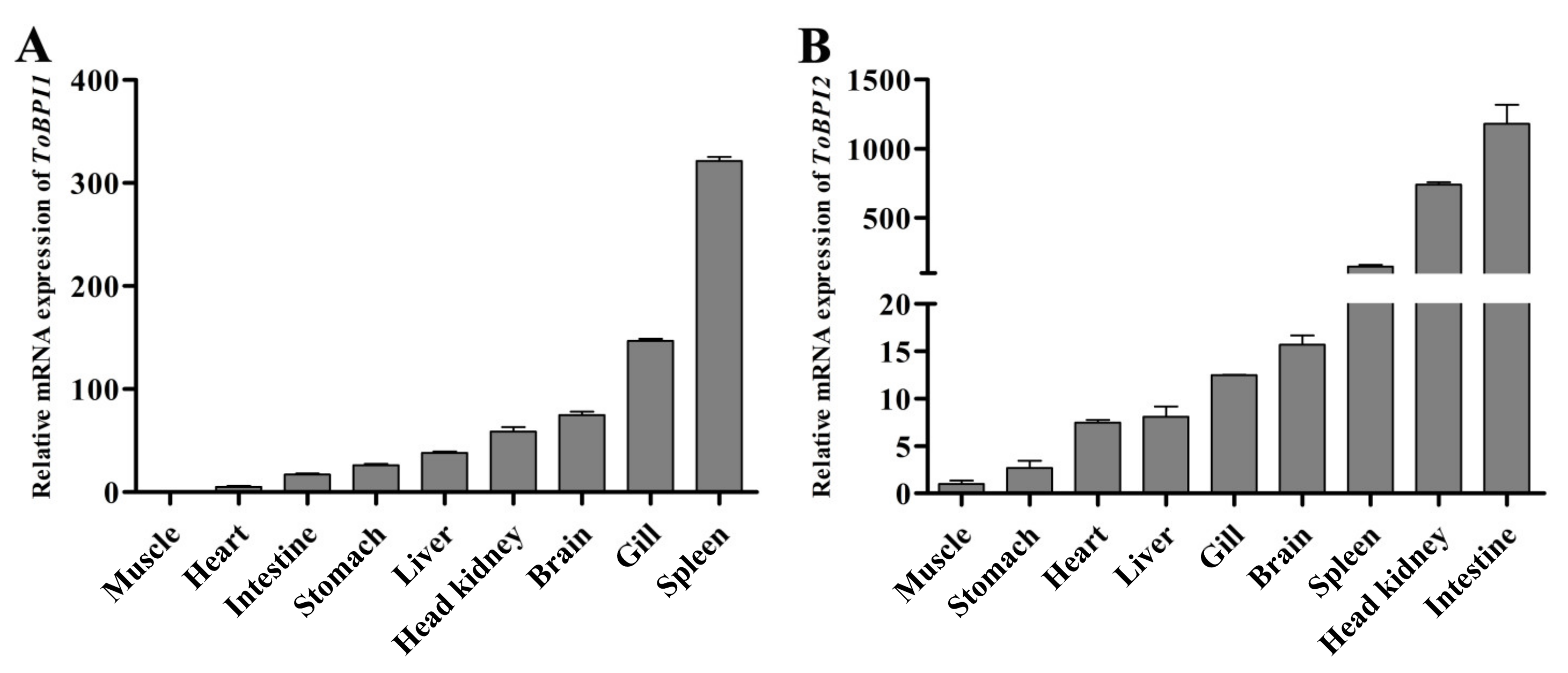
3.2. Tissue Distribution of ToBPI1/LBP and ToBPI2/LBP in Healthy T. ovatus
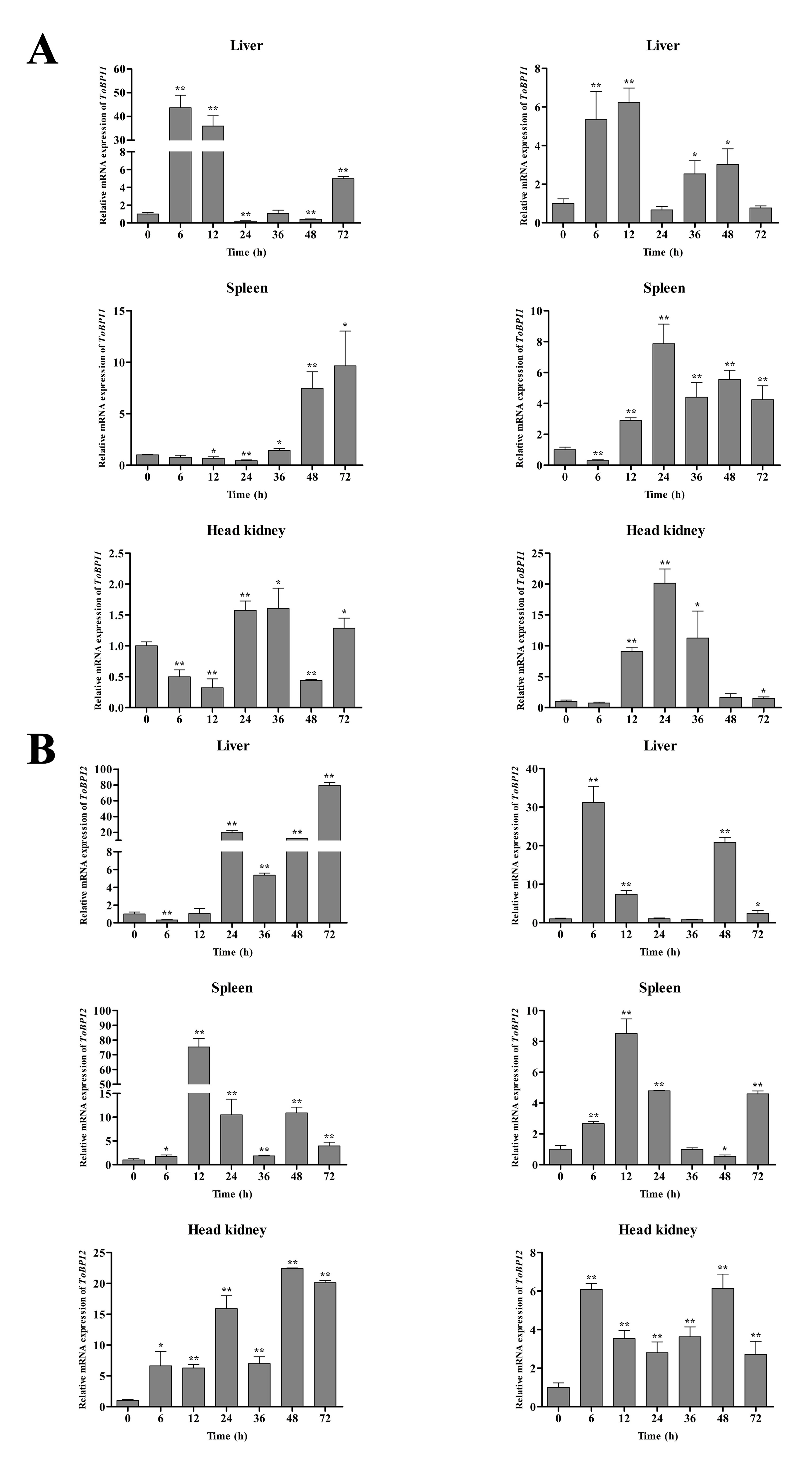
3.3. Expression of ToBPI1/LBP and ToBPI2/LBP in Different Tissues after Bacterial Infection
3.4. Recombinant Expression and Purification of rToBPI1/LBP and rToBPI2/LBP
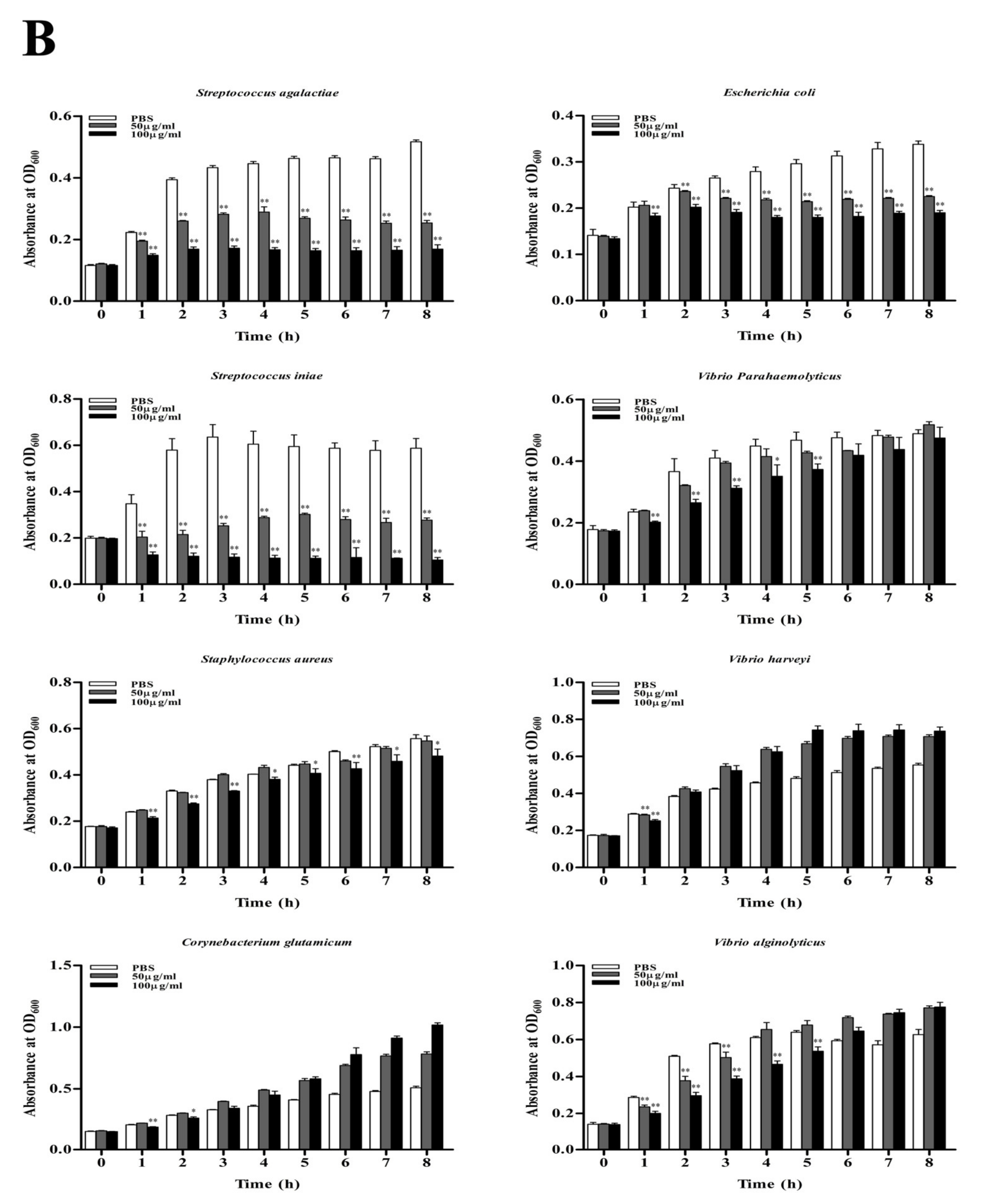
3.5. Antimicrobial Activity
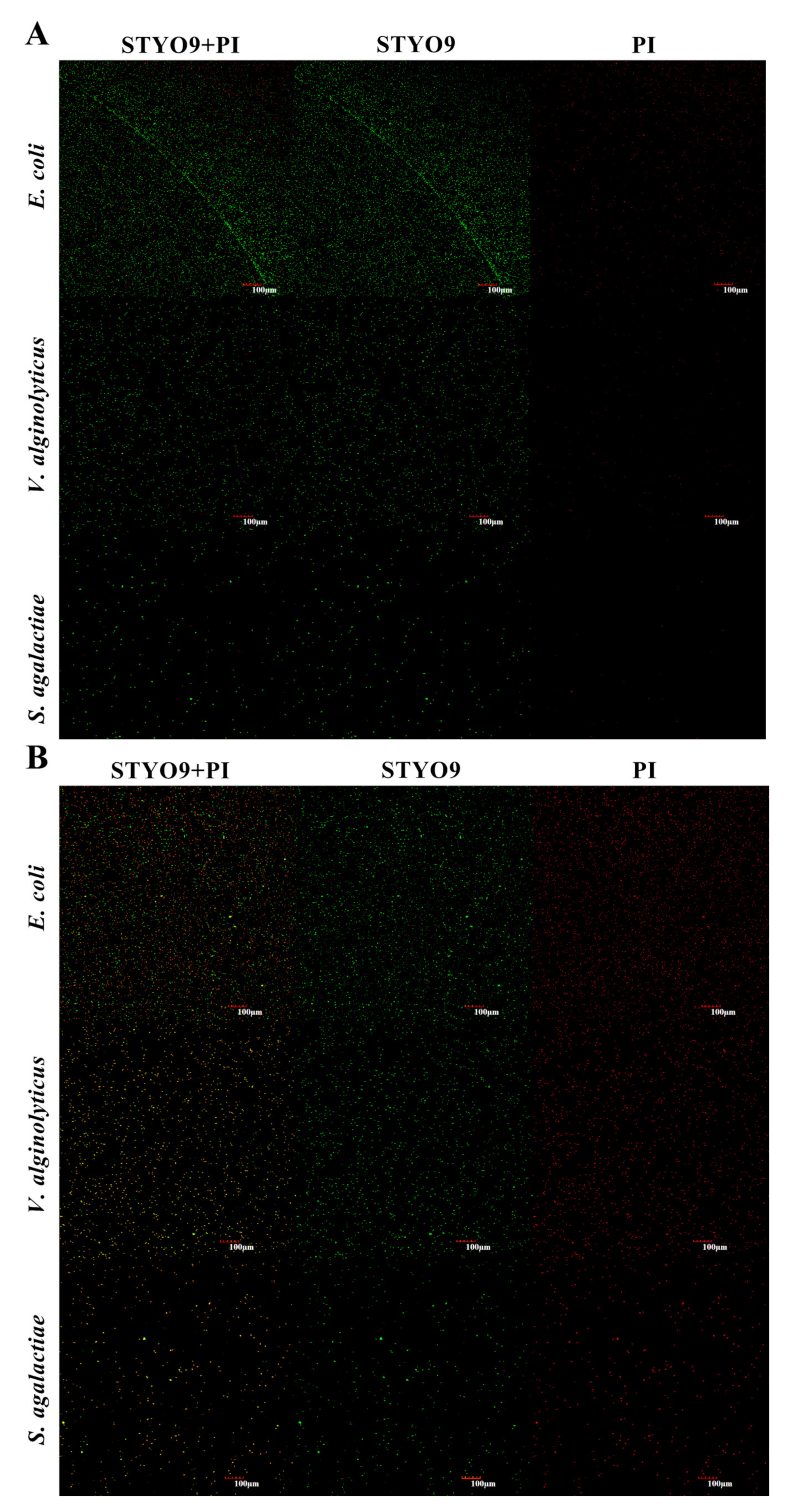
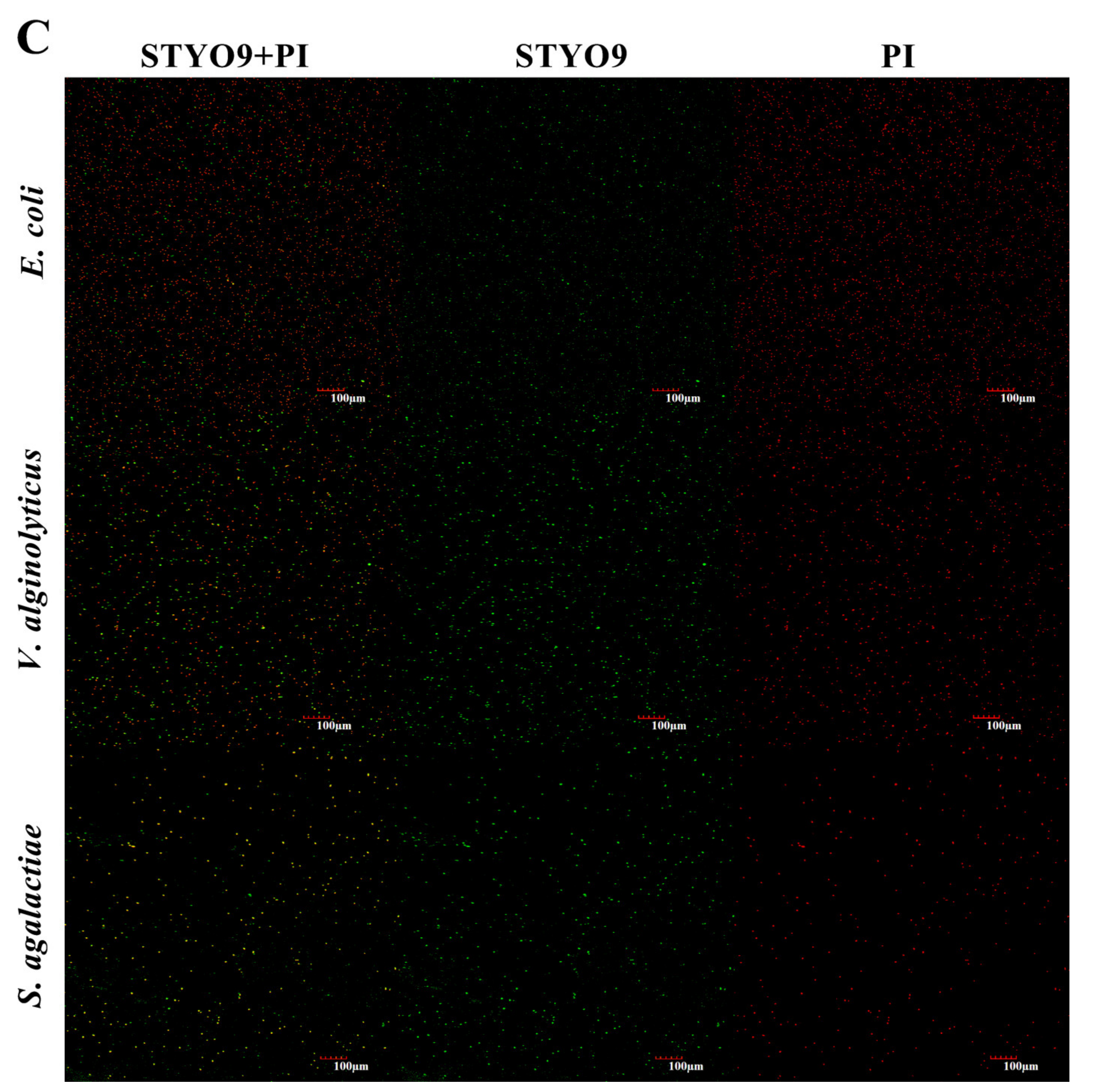
3.6. Permeability of Cell Membrane
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shao, Y.; Li, C.; Che, Z.; Zhang, P.; Zhang, W.; Duan, X.; Li, Y. Cloning and characterization of two lipopolysaccharide-binding protein/bactericidal permeability-increasing protein (LBP/BPI) genes from the sea cucumber Apostichopus japonicus with diversified function in modulating ROS production. Dev. Comp. Immunol. 2015, 52, 88–97. [Google Scholar] [CrossRef]
- Mao, Y.; Zhou, C.; Zhu, L.; Huang, Y.; Yan, T.; Fang, J.; Zhu, W. Identification and expression analysis on bactericidal permeability-increasing protein (BPI)/lipopolysaccharide-binding protein (LBP) of ark shell, Scapharca broughtonii. Fish Shellfish. Immunol. 2013, 35, 642–652. [Google Scholar] [CrossRef]
- Tang, L.; Liang, Y.; Jiang, Y.; Liu, S.; Zhang, F.; He, X.; Wang, T.; Zhou, Y.; Zhong, H.; Yan, J. Identification and expression analysis on bactericidal permeability-increasing protein/lipopolysaccharide-binding protein of blunt snout bream, Megalobrama amblycephala. Fish Shellfish. Immunol. 2015, 45, 630–640. [Google Scholar] [CrossRef]
- Su, J.; Li, H.; Hu, J.; Wang, D.; Zhang, F.; Fu, Z.; Han, F. LcCCL28-25, Derived from Piscine Chemokine, Exhibits Antimicrobial Activity against Gram-Negative and Gram-Positive Bacteria In Vitro and In Vivo. Microbiol. Spectr. 2022, 10, e0251521. [Google Scholar] [CrossRef]
- Bin Hafeez, A.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial Peptides: An Update on Classifications and Databases. Int. J. Mol. Sci. 2021, 22, 11691. [Google Scholar] [CrossRef]
- Kim, J.W.; Gerwick, L.; Park, C.I. Molecular identification and expression analysis of two distinct BPI/LBPs (bactericidal permeability-increasing protein/LPS-binding protein) from rock bream, Oplegnathus fasciatus. Fish Shellfish. Immunol. 2012, 33, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J. Bactericidal/permeability-increasing protein (BPI) and lipopolysaccharide-binding protein (LBP): Structure, function and regulation in host defence against Gram-negative bacteria. Biochem. Soc. Trans. 2003, 31, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Schultz, H.; Weiss, J.P. The bactericidal/permeability-increasing protein (BPI) in infection and inflammatory disease. Clin. Chim. Acta 2007, 384, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.P.; Xia, X.Y.; Guo, Q.S.; Xu, C. Bactericidal/permeability-increasing protein originates in both the testis and the epididymis and localizes in mouse spermatozoa. Asian J. Androl. 2014, 16, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, I.; Schonefeld, M.; Aichele, D.; Groer, G.; Gessner, A.; Schnare, M. Murine bactericidal/permeability-increasing protein inhibits the endotoxic activity of lipopolysaccharide and gram-negative bacteria. J. Immunol. 2008, 180, 7546–7552. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, H.; Gogami, A.; Miyagawa, Y.; Nanbo, A.; Murakami, Y.; Baba, T.; Nagasawa, S. Bactericidal/permeability-increasing protein promotes complement activation for neutrophil-mediated phagocytosis on bacterial surface. Immunology 2001, 103, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Elvitigala, D.A.S.; Lee, S.; Kim, H.C.; Park, H.C.; Lee, J. Molecular characterization of a bactericidal permeability-increasing protein/lipopolysaccharide-binding protein from black rockfish (Sebastes schlegelii): Deciphering its putative antibacterial role. Dev. Comp. Immunol. 2017, 67, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Y.; Sun, L. A Teleost Bactericidal Permeability-Increasing Protein Kills Gram-Negative Bacteria, Modulates Innate Immune Response, and Enhances Resistance against Bacterial and Viral Infection. PLoS ONE 2016, 11, e0154045. [Google Scholar] [CrossRef]
- Zhou, S.; Jiang, G.; Zhu, Y.; Liu, L.; Liu, D.; Diao, J.; Liu, H.; Xiu, Y. Molecular identification and function analysis of bactericidal permeability-increasing protein/LPS-binding protein 1 (BPI/LBP1) from turbot (Scophthalmus maximus). Fish Shellfish. Immunol. 2019, 87, 499–506. [Google Scholar] [CrossRef]
- Cai, X.H.; Peng, Y.H.; Wang, Z.C.; Huang, T.; Xiong, X.Y.; Huang, Y.C.; Wang, B.; Xu, L.W.; Wu, Z.H. Characterization and identification of streptococci from golden pompano in China. Dis. Aquat. Organ 2016, 119, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Guo, H.-Y.; Liu, M.-J.; Zhu, K.-C.; Liu, B.; Liu, B.-S.; Zhang, N.; Jiang, S.-G.; Zhang, D.-C. Transcriptome Analysis of the Immune Process of Golden Pompano (Trachinotus ovatus) Infected with Streptococcus agalactiae. Fishes 2023, 8, 52. [Google Scholar] [CrossRef]
- Wu, Y.; Du, H.H.; Zhu, L.; Zhao, N.; Zhang, S.; Cao, Z.; Zhou, Y.; Sun, Y. Bactericidal permeability-increasing protein/LPS-binding protein (BPI/LBP) enhances resistance of golden pompano Trachinotus ovatus against bacterial infection. Fish Shellfish. Immunol. 2022, 131, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Lei, Y.; Cao, Z.; Chen, X.; Sun, Y.; Xu, Y.; Guo, W.; Wang, S.; Liu, C. A β-defensin gene of Trachinotus ovatus might be involved in the antimicrobial and antiviral immune response. Dev. Comp. Immunol. 2019, 92, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Fan, J.; Wang, W.; Wang, H.; Liu, H. Molecular characterization, expression and antimicrobial activity of complement factor D in Megalobrama amblycephala. Fish Shellfish. Immunol. 2019, 89, 43–51. [Google Scholar] [CrossRef]
- Liu, Y.; Zha, H.; Han, X.; Yu, S.; Chai, Y.; Zhong, J.; Zhu, Q. Molecular characterization and functional analysis of the bactericidal permeability-increasing protein/LPS-binding protein (BPI/LBP) from roughskin sculpin (Trachidermus fasciatus). Dev. Comp. Immunol. 2021, 123, 104133. [Google Scholar] [CrossRef]
- Inagawa, H.; Honda, T.; Kohchi, C.; Nishizawa, T.; Yoshiura, Y.; Nakanishi, T.; Yokomizo, Y.; Soma, G. Cloning and characterization of the homolog of mammalian lipopolysaccharide-binding protein and bactericidal permeability-increasing protein in rainbow trout Oncorhynchus mykiss. J. Immunol. 2002, 168, 5638–5644. [Google Scholar] [CrossRef]
- Kono, T.; Sakai, M. Molecular cloning of a novel bactericidal permeability-increasing protein/lipopolysaccharide-binding protein (BPI/LBP) from common carp Cyprinus carpio L. and its expression. Mol. Immunol. 2003, 40, 269–278. [Google Scholar] [CrossRef]
- Theprungsirikul, J.; Skopelja-Gardner, S.; Rigby, W.F.C. Killing three birds with one BPI: Bactericidal, opsonic, and anti-inflammatory functions. J. Transl. Autoimmun. 2021, 4, 100105. [Google Scholar] [CrossRef] [PubMed]
- Beamer, L.J.; Carroll, S.F.; Eisenberg, D. The BPI/LBP family of proteins: A structural analysis of conserved regions. Protein Sci. 1998, 7, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Nam, B.H.; Ahn, K.J.; Kim, Y.O.; Kong, H.J.; Kim, W.J.; Kim, H.S.; Lee, S.J.; Kim, K.K. Molecular cloning and characterization of LPS-binding protein/bactericidal permeability-increasing protein (LBP/BPI) from olive flounder, Paralichthys olivaceus. Vet. Immunol. Immunopathol. 2010, 133, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Flajnik, M.F. A cold-blooded view of adaptive immunity. Nat. Rev. Immunol. 2018, 18, 438–453. [Google Scholar] [CrossRef] [PubMed]
- Ofek, T.; Lalzar, M.; Izhaki, I.; Halpern, M. Intestine and spleen microbiota composition in healthy and diseased tilapia. Anim. Microbiome 2022, 4, 50. [Google Scholar] [CrossRef]
- Press, C.M.; Evensen, Ø. The morphology of the immune system in teleost fishes. Fish Shellfish. Immunol. 1999, 9, 309–318. [Google Scholar] [CrossRef]
- Jiang, C.; Wang, S. Identification and functional characterization of bactericidal permeability/increasing protein (BPI) from frog Nanorana yunnanensis (Paa yunnanensis). Dev. Comp. Immunol. 2022, 137, 104517. [Google Scholar] [CrossRef]
- Horwitz, A.H.; Williams, R.E.; Liu, P.S.; Nadell, R. Bactericidal/permeability-increasing protein inhibits growth of a strain of Acholeplasma laidlawii and L forms of the gram-positive bacteria Staphylococcus aureus and Streptococcus pyogenes. Antimicrob. Agents Chemother. 1999, 43, 2314–2316. [Google Scholar] [CrossRef]
- Wiese, A.; Brandenburg, K.; Carroll, S.F.; Rietschel, E.T.; Seydel, U. Mechanisms of action of bactericidal/permeability-increasing protein BPI on reconstituted outer membranes of gram-negative bacteria. Biochemistry 1997, 36, 10311–10319. [Google Scholar] [CrossRef] [PubMed]
- Bulow, S.; Zeller, L.; Werner, M.; Toelge, M.; Holzinger, J.; Entzian, C.; Schubert, T.; Waldow, F.; Gisch, N.; Hammerschmidt, S.; et al. Bactericidal/Permeability-Increasing Protein Is an Enhancer of Bacterial Lipoprotein Recognition. Front. Immunol. 2018, 9, 2768. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers Name | Sequence 5′–3′ | Purpose |
|---|---|---|
| ToBPI1/LBP -F | ATGTTTCTAAACCTAATAGTAGTGCTCATGC | Cloning |
| ToBPI1/LBP -R | TCAGTTGAAGTCTCTGTCTGTTAGG | Cloning |
| ToBPI2/LBP -F | ATGTCCCTGTGCCGTTG | Cloning |
| ToBPI2/LBP -R | TTAACCCGTAAACTGAACATCTGTC | Cloning |
| ToBPI1/LBP -F | CCTTTGCCATCCAGCCAAAT | Real-time PCR |
| ToBPI1/LBP -R | CCACTTCACTTCCTGCCAAC | Real-time PCR |
| ToBPI2/LBP -F | GCCCACAGTTTCAACGTCTT | Real-time PCR |
| ToBPI2/LBP -R | GTGTTGAGTCGGACAGGAGA | Real-time PCR |
| β-action-F | TACGAGCTGCCTGACGGACA | Real-time PCR |
| β-action-R | GGCTGTGATCTCCTTCTGC | Real-time PCR |
| mToBPI1 LBP-F | CGCGGATCCGAAAATCCTGCAATACAAGTC | Prokaryotic expression |
| mToBPI1/LBP-R | CCGCTCGAGGTTGAAGTCTCTGTCTGTTAGG | Prokaryotic expression |
| mToBPI2/LBP-F | CGGAATTCGTCAATCCTGGGGTAAATGT | Prokaryotic expression |
| mToBPI2/LBP-R | CCCTCGAGACCCGTAAACTGAACATCTGT | Prokaryotic expression |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bian, Z.-C.; Cai, X.-H.; Tan, K.A.; Wang, Y.-D.; Huang, Z.; Kwan, K.Y.; Xu, P. Identification and Functional Analysis of ToBPI1/LBP and ToBPI2/LBP in Anti-Bacterial Infection of Trachinotus ovatus. Genes 2023, 14, 826. https://doi.org/10.3390/genes14040826
Bian Z-C, Cai X-H, Tan KA, Wang Y-D, Huang Z, Kwan KY, Xu P. Identification and Functional Analysis of ToBPI1/LBP and ToBPI2/LBP in Anti-Bacterial Infection of Trachinotus ovatus. Genes. 2023; 14(4):826. https://doi.org/10.3390/genes14040826
Chicago/Turabian StyleBian, Ze-Chang, Xiao-Hui Cai, Kian Ann Tan, Ya-Dan Wang, Zhuang Huang, Kit Yue Kwan, and Peng Xu. 2023. "Identification and Functional Analysis of ToBPI1/LBP and ToBPI2/LBP in Anti-Bacterial Infection of Trachinotus ovatus" Genes 14, no. 4: 826. https://doi.org/10.3390/genes14040826