Association of Single Nucleotide Polymorphism in the DGAT1 Gene with the Fatty Acid Composition of Cows Milked Once and Twice a Day

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Farm and Feeding
2.2. Cows and Milk Sampling
2.3. Determination of Milk Composition and Fatty Acid Composition
2.4. Genotyping
2.5. Statistical Analysis
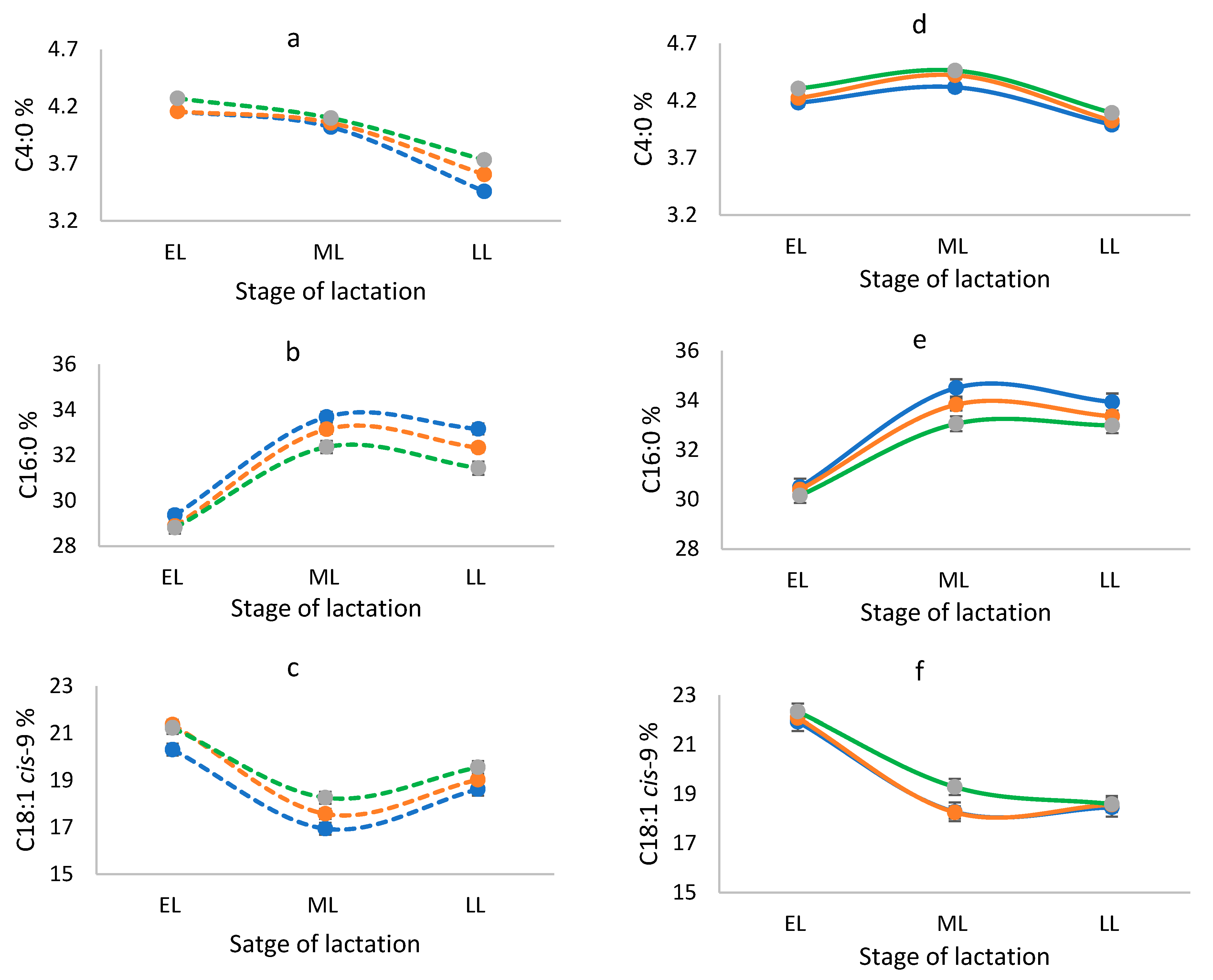
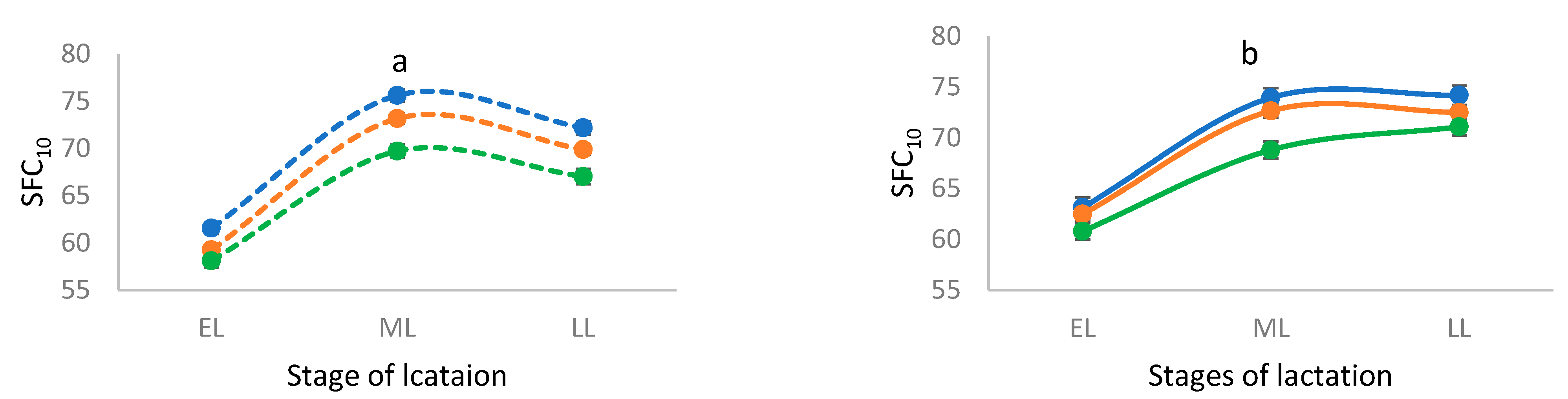
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davis, S.R.; Farr, V.C.; Stelwagen, K. Regulation of yield loss and milk composition during once-daily milking: A review. Livest. Prod. Sci. 1999, 59, 77–94. [Google Scholar] [CrossRef]
- Clark, D.A.; Phyn, C.V.C.; Tong, M.J.; Collis, S.J.; Dalley, D.E. A systems comparison of once-versus twice daily milking of pastured dairy cows. J. Dairy Sci. 2006, 89, 1854–1862. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.P. Comparison of milk production and herd characteristics in New Zealand herds milked once or twice a day. Anim. Prod. Sci. 2019, 59, 570–580. [Google Scholar] [CrossRef]
- Rémond, B.; Pomiès, D.; Dupont, D.; Chilliard, Y. Once-a-day milking of multiparous Holstein cows throughout the entire lactation: Milk yield and composition, and nutritional status. Anim. Res. 2004, 53, 201–212. [Google Scholar] [CrossRef]
- Lembeye, F.; López-Villalobos, N.; Burke, J.L.; Davis, S.R. Estimation of breed and heterosis effects for milk traits and somatic cell scores in cows milked once and twice daily in New Zealand. Proc. N. Z. Soc. Anim. Prod. 2015, 75, 60–63. [Google Scholar]
- Sneddon, N.W.; Lopez-Villalobos, N.; Hickson, R.E.; Shalloo, L.; Garrick, D.J. Estimation of crossbreeding effects on yields of dairy products and value of milk processed in different product portfolios. Proc. N. Z. Soc. Anim. Prod. 2015, 75, 48–53. [Google Scholar]
- Dalgleish, D.G. Bovine milk protein properties and the manufacturing quality of milk. Livest. Prod. Sci. 1993, 35, 75–93. [Google Scholar] [CrossRef]
- Walstra, P.; Wouters, J.T.; Geurts, T.J. Dairy Science and Technology; CRC Press, Taylor & Francis: Boca Raton, FL, USA, 2005. [Google Scholar]
- MacGibbon, A.K.H. Composition and Structure of Bovine Milk Lipids. In Advanced Dairy Chemistry, 4th ed.; McSweeney, P.L.H., Fox, P.F., O’Mahony, J.A., Eds.; Springer: New York, NY, USA, 2020; Volume 2, pp. 1–32. [Google Scholar]
- Soyeurt, H.; Dardenne, P.; Gillon, A.; Croquet, C.; Vanderick, S.; Mayeres, P.; Bertozzi, C.; Gengler, N. Variation in fatty acid contents of milk and milk fat within and across breeds. J. Dairy Sci. 2006, 89, 4858–4865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palladino, R.A.; Buckley, F.; Prendiville, R.; Murphy, J.J.; Callan, J.; Kenny, D.A. A comparison between Holstein-Friesian and Jersey dairy cows and their F1 hybrid on milk fatty acid composition under grazing conditions. J. Dairy Sci. 2010, 93, 2176–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Villalobos, N.; Spelman, R.J.; Melis, J.; Davis, S.R.; Berry, S.D.; Lehnert, K.; Holroyd, S.E.; MacGibbon, A.K.; Snell, R.G. Estimation of genetic and crossbreeding parameters of fatty acid concentrations in milk fat predicted by mid-infrared spectroscopy in New Zealand dairy cattle. J. Dairy Sci. 2014, 81, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Back, P.J.; Thomson, N.A. Exploiting cow genotype to increase milk value through production of minor milk components. Proc. N. Z. Soc. Anim. Prod. 2005, 65, 53–58. [Google Scholar]
- Stoop, W.M.; Bovenhuis, H.; Heck, J.M.L.; van Arendonk, J.A.M. Effect of lactation stage and energy status on milk fat composition of Holstein-Friesian cows. J. Dairy Sci. 2009, 92, 1469–1478. [Google Scholar] [CrossRef] [Green Version]
- Palmquist, D.L.; Beaulieu, A.D.; Barbano, D.M. Feed and animal factors influencing milk fat composition. J. Dairy Sci. 1993, 76, 1753–1771. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Shingfield, K.J.; Lee, M.R.; Scollan, N.D. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems. Anim. Feed Sci. Technol. 2006, 131, 168–206. [Google Scholar] [CrossRef]
- Heck, J.M.; Van Valenberg, H.J.; Dijkstra, J.; Van Hooijdonk, A.C. Seasonal variation in the Dutch bovine raw milk composition. J. Dairy Sci. 2009, 92, 4745–4755. [Google Scholar] [CrossRef] [Green Version]
- Schwendel, B.H.; Morel, P.C.; Wester, T.J.; Tavendale, M.H.; Deadman, C.; Fong, B.; Shadbolt, N.M.; Thatcher, A.; Otter, D.E. Fatty acid profile differs between organic and conventionally produced cow milk independent of season or milking time. J. Dairy Sci. 2015, 98, 1411–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacGibbon, A.K. Herd-to-herd variations in the properties of milkfat. Proc. N. Z. Soc. Anim. Prod. 1996, 56, 224–227. [Google Scholar]
- MacGibbon, A.K.; McLennan, W.D. Hardness of New-Zealand patted butter-seasonal and regional variations. N. Z. J. Dairy Sci. Technol. 1987, 22, 143–156. [Google Scholar]
- Stoop, W.M.; Schennink, A.; Visker, M.H.P.W.; Mullaart, E.; Van Arendonk, J.A.M.; Bovenhuis, H. Genome-wide scan for bovine milk-fat composition. I. Quantitative trait loci for short-and medium-chain fatty acids. J. Dairy Sci. 2009, 92, 4664–4675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutsen, T.M.; Olsen, H.G.; Tafintseva, V.; Svendsen, M.; Kohler, A.; Kent, M.P.; Lien, S. Unravelling genetic variation underlying de novo-synthesis of bovine milk fatty acids. Sci. Rep. 2018, 8, 2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spelman, R.J.; Ford, C.A.; McElhinney, P.; Gregory, G.C.; Snell, R.G. Characterization of the DGAT1 gene in the New Zealand dairy population. J. Dairy Sci. 2002, 85, 3514–3517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bovenhuis, H.; Visker, M.H.; Poulsen, N.A.; Sehested, J.; Van Valenberg, H.J.; Van Arendonk, J.A.; Larsen, L.B.; Buitenhuis, A.J. Effects of the diacylglycerol o-acyltransferase 1 (DGAT1) K232A polymorphism on fatty acid, protein, and mineral composition of dairy cattle milk. J. Dairy Sci. 2016, 99, 3113–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhou, H.; Cheng, L.; Edwards, G.R.; Hickford, J.G. Effect of DGAT1 variant (K232A) on milk traits and milk fat composition in outdoor pasture-grazed dairy cattle. N. Z. J. Agric. Res. 2021, 64, 101–113. [Google Scholar] [CrossRef]
- Schennink, A.; Stoop, W.M.; Visker, M.W.; Heck, J.M.; Bovenhuis, H.; Van Der Poel, J.J.; Van Valenberg, H.J.; Van Arendonk, J.A. DGAT1 underlies large genetic variation in milk-fat composition of dairy cows. Anim. Genet. 2007, 38, 467–473. [Google Scholar] [CrossRef]
- Tumino, S.; Criscione, A.; Moltisanti, V.; Marletta, D.; Bordonaro, S.; Avondo, M.; Valenti, B. Feeding System Resizes the Effects of DGAT1 Polymorphism on Milk Traits and Fatty Acids Composition in Modicana Cows. Animals 2021, 11, 1616. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Villalobos, N.; Ariyarathne, H.B.P.C.; Gedye, K.; Correa-Luna, M.; Donaghy, D.J. Association of a SNP in the DGAT1 gene with productive and reproductive performance and profitability in grazing cows milked once and twice a day. J. Dairy Sci. 2020, 103, 74. [Google Scholar]
- Sukhija, P.S.; Palmquist, D.L. Rapid method for determination of total fatty acid content and composition of feedstuffs and feces. J. Agric. Food Chem. 1988, 36, 1202–1206. [Google Scholar] [CrossRef]
- Wigginton, J.E.; Cutler, D.J.; Abecasis, G.R. A note on exact tests of Hardy-Weinberg equilibrium. Am. J. Hum. Genet. 2005, 76, 887–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Ma, L.; Prakapenka, D.; VanRaden, P.M.; Cole, J.B.; Da, Y. A large-scale genome-wide association study in US Holstein cattle. Front. Genet. 2019, 10, 412. [Google Scholar] [CrossRef]
- Grisart, B.; Farnir, F.; Karim, L.; Cambisano, N.; Kim, J.J.; Kvasz, A.; Mni, M.; Simon, P.; Frere, J.M.; Coppieters, W.; et al. Genetic and functional confirmation of the causality of the DGAT1 K232A quantitative trait nucleotide in affecting milk yield and composition. Proc. Natl. Acad. Sci. USA 2004, 101, 2398–2403. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Villalobos, N.; Spelman, R.J.; Melis, J.; Davis, S.R.; Berry, S.D.; Lehnert, K.; Sneddon, N.W.; Holroyd, S.E.; MacGibbon, A.K.; Snell, R.G. Genetic correlations of milk fatty acid contents predicted from milk mid-infrared spectra in New Zealand dairy cattle. J. Dairy Sci. 2020, 103, 7238–7248. [Google Scholar] [CrossRef]
- Soyeurt, H.; Gillon, A.; Vanderick, S.; Mayeres, P.; Bertozzi, C.; Gengler, N. Estimation of heritability and genetic correlations for the major fatty acids in bovine milk. J. Dairy Sci. 2007, 90, 4435–4442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, M.O.; Knudsen, J. Specificity of diacylglycerol acyltrans-ferase from bovine mammary gland, liver and adipose tissue towards acyl-CoA esters. Eur. J. Biochem. 1979, 94, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Ariyarathne, H.B.P.C.; Correa-Luna, M.; Blair, H.T.; Garrick, D.J.; Lopez-Villalobos, N. Identification of genomic regions associated with concentrations of milk fat, protein, urea and efficiency of crude protein utilization in grazing dairy cows. Genes 2021, 12, 456. [Google Scholar] [CrossRef]
- Mackle, T.R.; Petch, S.F.; Bryant, A.M.; Auldist, M.J.; Henderson, H.V.; MacGibbon, A.K.H. Variation in the characteristics of milkfat from pasture-fed dairy cows during late spring and the effects of grain supplementation. N. Z. J. Agric. Res. 1997, 40, 349–359. [Google Scholar] [CrossRef]
- Kaylegian, K.E. The production of specialty milk fat ingredients. J. Dairy Sci. 1999, 82, 1433–1439. [Google Scholar] [CrossRef]
- Sanjayaranj, I.; Lopez-Villalobos, N.; Blair, H.T.; Janssen, P.W.M.; Holroyd, S.E.; MacGibbon, A.K.H. Fatty Acid Composition of Dairy Milk: A Case Study Comparing Once- and Twice-a-Day Milking of Pasture-Fed Cows at Different Stages of Lactation. Dairy 2022, 3, 174–189. [Google Scholar] [CrossRef]
- Falconer, D.; Mackay, M. Introduction to Quantitative Genetics, 4th ed.; Logman Group: London, UK, 1996; 464p. [Google Scholar]
- Lopez-Villalobos, N. Analysing the genetic basis of milk production traits. CABI Rev. 2012, 1–18. [Google Scholar] [CrossRef]
- Delamaire, E.; Guinard-Flament, J. Increasing milking intervals decreases the mammary blood flow and mammary uptake of nutrients in dairy cows. J. Dairy Sci. 2006, 89, 3439–3446. [Google Scholar] [CrossRef] [Green Version]
- Bionaz, M.; Periasamy, K.; Rodriguez-Zas, S.L.; Hurley, W.L.; Loor, J.J. A novel dynamic impact approach (DIA) for functional analysis of time-course omics studies: Validation using the bovine mammary transcriptome. PLoS ONE 2012, 7, e32455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kęsek-Woźniak, M.; Wojtas, E.; Zielak-Steciwko, A.E. Impact of SNPs in ACACA, SCD1, and DGAT1 genes on fatty acid profile in bovine milk with regard to lactation phases. Animals 2020, 10, 997. [Google Scholar] [CrossRef] [PubMed]
- Auldist, M.J.; Walsh, B.J.; Thomson, N.A. Seasonal and lactational influences on bovine milk composition in New Zealand. J. Dairy Res. 1998, 65, 401–411. [Google Scholar] [CrossRef] [PubMed]




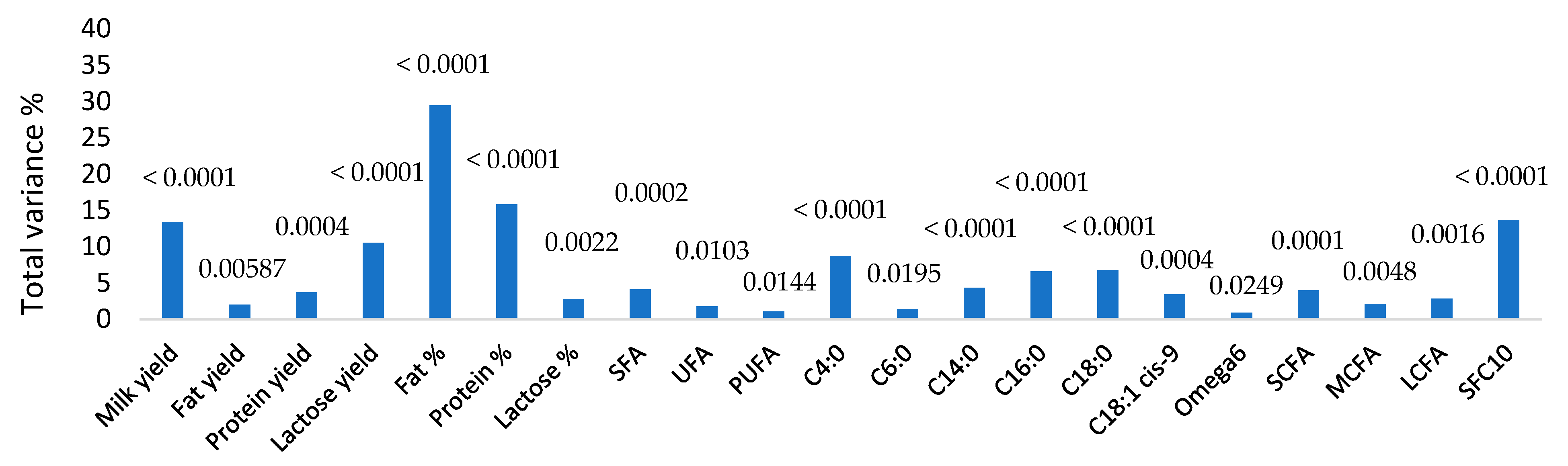{kind=link}
{kind=link}
{kind=link}
| Farm | No. 1 Dairy (OAD 12) | No. 4 Dairy (TAD 13) |
| Feed allocation (kg DM per cow per day) | ||
| Pasture 1 | 8.2 | 14.7 |
| Herb mix crop 2 | 3.7 | - |
| Maize silage | 1.0 | 4.0 |
| DDG 3 | 1.5 | 1.0 |
| Tapioca pellets | 0.8 | - |
| Concentrate 4 | 2.0 | 2.5 |
| Dry roughage 5 | - | 0.6 |
| Baleage 6 | 4.0 | 5.5 |
| Feed chemical composition | ||
| ME 7 MJ/kg DM | 11.33 | 11.07 |
| CP 8 g/100 g DM | 20.49 | 19.95 |
| NDF 9 g/100 g DM | 38.91 | 44.43 |
| ADF 10 g/100 g DM | 22.44 | 23.24 |
| SSS 11 g/100 g DM | 13.22 | 15.09 |
| Lipid g/100 g DM | 4.44 | 4.42 |
| Milking Frequency | OAD | TAD | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | N | Mean | SD | CV% | Min | Max | N | Mean | SD | CV% | Min | Max |
| Milk yield (L/cow/day) | 668 | 17.89 | 6.3 | 35.2 | 4.6 | 39.9 | 543 | 19.50 | 5.56 | 28.5 | 6.40 | 39.50 |
| Fat yield (kg/cow/day) | 668 | 0.91 | 0.32 | 35.4 | 0.26 | 5.22 | 543 | 0.88 | 0.22 | 24.6 | 0.31 | 1.92 |
| Protein yield (kg/cow/day) | 668 | 0.73 | 0.22 | 30.3 | 0.21 | 1.48 | 543 | 0.71 | 0.17 | 23.6 | 0.29 | 1.54 |
| Lactose yield (kg/cow/day) | 668 | 0.87 | 0.33 | 38 | 0.19 | 2.13 | 543 | 0.99 | 0.30 | 30.5 | 0.29 | 2.08 |
| Fat percentage | 694 | 4.86 | 1.04 | 21.3 | 1.72 | 8.53 | 543 | 3.98 | 0.85 | 21.4 | 1.27 | 6.89 |
| Protein percentage | 694 | 4.14 | 0.4 | 9.6 | 3.19 | 5.67 | 543 | 3.76 | 0.37 | 9.9 | 2.74 | 5.01 |
| Lactose percentage | 694 | 4.83 | 0.26 | 5.3 | 3.7 | 5.43 | 543 | 5.03 | 0.24 | 4.7 | 4.29 | 5.51 |
| Fatty acid (% of the total FA) | ||||||||||||
| SFA 1 | 694 | 70.65 | 3.31 | 4.7 | 51.57 | 81.78 | 543 | 70.08 | 2.96 | 4.2 | 58.64 | 77.94 |
| UFA 2 | 694 | 29.85 | 2.68 | 9 | 21.71 | 43.96 | 543 | 29.94 | 2.54 | 8.5 | 23.70 | 38.84 |
| PUFA 3 | 694 | 3.13 | 0.52 | 16.7 | 2.01 | 4.61 | 543 | 2.87 | 0.41 | 14.5 | 1.49 | 3.85 |
| C4:0 | 694 | 3.95 | 0.34 | 8.7 | 2.5 | 4.89 | 543 | 4.11 | 0.25 | 6.0 | 2.52 | 4.65 |
| C6:0 | 694 | 2.87 | 0.25 | 8.7 | 1.37 | 3.49 | 543 | 2.83 | 0.20 | 6.9 | 1.71 | 3.29 |
| C8:0 | 694 | 1.6 | 0.17 | 10.7 | 0.56 | 2.05 | 543 | 1.50 | 0.12 | 8.3 | 0.68 | 1.82 |
| C10:0 | 694 | 3.7 | 0.46 | 12.3 | 0.93 | 4.93 | 543 | 3.33 | 0.34 | 10.3 | 1.19 | 4.38 |
| C12:0 | 694 | 4.08 | 0.37 | 9.1 | 1.9 | 5.17 | 543 | 3.72 | 0.29 | 7.7 | 2.46 | 4.46 |
| C14:0 | 693 | 13.07 | 1.35 | 10.3 | 5.44 | 16.05 | 543 | 12.58 | 1.22 | 9.7 | 7.14 | 15.27 |
| C16:0 | 694 | 31.52 | 2.83 | 9 | 22.86 | 38.69 | 543 | 32.10 | 2.47 | 7.7 | 25.17 | 39.81 |
| C18:0 | 694 | 12.62 | 1.59 | 12.6 | 7.87 | 22.23 | 543 | 13.28 | 1.22 | 9.2 | 9.94 | 21.37 |
| C18:1 cis-9 | 694 | 19.18 | 2.76 | 14.4 | 11.81 | 37.88 | 543 | 20.54 | 2.69 | 13.1 | 14.75 | 31.99 |
| Omega-6 | 694 | 1.74 | 0.51 | 29.5 | 0.47 | 4.03 | 543 | 1.55 | 0.31 | 20.1 | 0.37 | 2.28 |
| SCFA 4 | 694 | 8.41 | 0.72 | 8.6 | 4.43 | 10.02 | 543 | 8.44 | 0.53 | 6.3 | 5.37 | 9.71 |
| MCFA 5 | 694 | 20.83 | 1.91 | 9.2 | 8.64 | 25.43 | 543 | 19.63 | 1.64 | 8.4 | 11.57 | 23.37 |
| LCFA 6 | 694 | 65.06 | 2.38 | 3.7 | 58.25 | 79.77 | 543 | 67.46 | 2.41 | 3.6 | 62.12 | 82.02 |
| Milk income (NZD/cow/day) | 668 | 14.8 | 4.64 | 31.3 | 4.26 | 57.33 | 543 | 14.34 | 3.37 | 23.5 | 6.27 | 31.16 |
| SFC10 7 | 694 | 67.53 | 8.6 | 12.7 | 17.7 | 89.57 | 543 | 67.44 | 7.13 | 10.6 | 41.03 | 86.96 |
| Fatty Acids | Milking Frequency | |||
|---|---|---|---|---|
| OAD | p-Value | TAD | p-Value | |
| SFA 1 | −0.09 | 0.0228 | −0.04 | 0.3954 |
| UFA 2 | 0.2 | <0.0001 | 0.14 | 0.0027 |
| PUFA 3 | −0.12 | 0.0022 | −0.11 | 0.0117 |
| C4:0 | −0.15 | <0.0001 | −0.43 | <0.0001 |
| C6:0 | −0.12 | 0.0017 | −0.34 | <0.0001 |
| C8:0 | 0.01 | 0.8817 | −0.20 | <0.0001 |
| C10:0 | −0.02 | 0.5528 | −0.18 | <0.0001 |
| C12:0 | −0.01 | 0.7613 | −0.11 | 0.0148 |
| C14:0 | −0.04 | 0.3424 | 0.04 | 0.3271 |
| C16:0 | 0.24 | <0.0001 | 0.32 | <0.0001 |
| C18:0 | 0.06 | 0.1455 | 0.23 | <0.0001 |
| C18:1 cis-9 | 0.17 | <0.0001 | 0.11 | 0.0136 |
| Omega6 | −0.21 | <0.0001 | −0.21 | <0.0001 |
| SCFA 4 | −0.13 | 0.0014 | −0.38 | <0.0001 |
| MCFA 5 | −0.03 | 0.4321 | −0.03 | 0.5355 |
| LCFA 6 | 0.37 | <0.0001 | 0.45 | <0.0001 |
| SFC10 7 | 0.03 | 0.5057 | 0.21 | <0.0001 |
| Variables | OAD | TAD | p-Value MF 8 ×SNP | ||||
|---|---|---|---|---|---|---|---|
| CC | CT | TT | CC | CT | TT | ||
| Milk yield (L/cow/day) | 16.57 ± 0.3 e | 18.23 ± 0.24 d | 18.83 ± 0.36 d | 22.17 ± 0.46 c | 23.56 ± 0.33 b | 25.00 ± 0.4 a | <0.0001 |
| Fat yield (kg/cow/day) | 0.93 ± 0.02 b | 0.91 ± 0.01 b | 0.85 ± 0.02 c | 1.06 ± 0.03 a | 1.06 ± 0.02 a | 1.03 ± 0.02 a | 0.015 |
| Protein yield (kg/cow/day) | 0.70 ± 0.01 d | 0.75 ± 0.01 c | 0.75 ± 0.01 c | 0.84 ± 0.02 b | 0.87 ± 0.01 ab | 0.89 ± 0.02 a | 0.0025 |
| Lactose yield (kg/cow/day) | 0.80 ± 0.02 e | 0.89 ± 0.01 e | 0.91 ± 0.02 d | 1.13 ± 0.02 c | 1.19 ± 0.02 b | 1.25 ± 0.02 a | <0.0001 |
| Fat percentage | 5.34 ± 0.07 a | 4.80 ± 0.05 b | 4.33 ± 0.08 c | 4.08 ± 0.1 c | 3.67 ± 0.07 d | 3.15 ± 0.09 e | <0.0001 |
| Protein percentage | 4.27 ± 0.03 a | 4.15 ± 0.02 b | 4.05 ± 0.03 c | 3.84 ± 0.04 d | 3.74 ± 0.03 e | 3.60 ± 0.04 f | <0.0001 |
| Lactose percentage | 4.80 ± 0.02 d | 4.85 ± 0.01 c | 4.79 ± 0.02 d | 5.07 ± 0.03 a | 5.04 ± 0.02 a | 4.97 ± 0.02 b | 0.0016 |
| Fatty acid (% of the total FA) | |||||||
| SFA 1 | 71.4 ± 0.21 a | 70.51 ± 0.17 bc | 70.1 ± 0.25 b | 71.55 ± 0.32 a | 71.42 ± 0.23 a | 70.88 ± 0.29 ac | 0.0016 |
| UFA 2 | 29.29 ± 0.18 b | 29.9 ± 0.14 a | 30.18 ± 0.21 a | 28.53 ± 0.27 c | 28.59 ± 0.19 c | 28.79 ± 0.24 bc | 0.0171 |
| PUFA 3 | 3.05 ± 0.03 b | 3.15 ± 0.02 a | 3.16 ± 0.03 a | 2.82 ± 0.04 c | 2.87 ± 0.03 c | 2.84 ± 0.04 c | 0.0291 |
| C4:0 | 3.89 ± 0.02 e | 3.94 ± 0.02 d | 4.03 ± 0.02 c | 4.17 ± 0.03 b | 4.23 ± 0.02 b | 4.29 ± 0.03 a | <0.0001 |
| C6:0 | 2.85 ± 0.01 c | 2.86 ± 0.01 c | 2.89 ± 0.02 bc | 2.89 ± 0.02 bc | 2.93 ± 0.02 ab | 2.95 ± 0.02 a | 0.0543 |
| C8:0 | 1.60 ± 0.01 a | 1.59 ± 0.01 ac | 1.59 ± 0.01 ac | 1.54 ± 0.01 b | 1.57 ± 0.01 bc | 1.56 ± 0.01 bc | 0.4008 |
| C10:0 | 3.72 ± 0.03 a | 3.67 ± 0.02 a | 3.67 ± 0.03 a | 3.41 ± 0.04 c | 3.5 ± 0.03 b | 3.48 ± 0.04 bc | 0.1831 |
| C12:0 | 4.12 ± 0.03 a | 4.05 ± 0.02 b | 4.03 ± 0.03 b | 3.77 ± 0.04 c | 3.81 ± 0.03 c | 3.80 ± 0.04 c | 0.133 |
| C14:0 | 13.31 ± 0.09 a | 13.06 ± 0.07b | 12.77 ± 0.1 cd | 12.91 ± 0.13 bc | 12.95 ± 0.09 bd | 12.63 ± 0.12 c | 0.0002 |
| C16:0 | 32.05 ± 0.18 b | 31.45 ± 0.14 c | 30.9 ± 0.21 d | 32.97 ± 0.27 a | 32.52 ± 0.19 ab | 32.06 ± 0.24 b | <0.0001 |
| C18:0 | 13.03 ± 0.12 b | 12.62 ± 0.09 c | 12.35 ± 0.14 cd | 13.59 ± 0.18 a | 13.12 ± 0.13 bc | 12.82 ± 0.16 bc | <0.0001 |
| C18:1 cis-9 | 18.59 ± 0.18 d | 19.30 ± 0.15 bc | 19.65 ± 0.21 acb | 19.53 ± 0.28 b | 19.62 ± 0.2 b | 20.06 ± 0.24 a | 0.0009 |
| Omega-6 | 1.66 ± 0.02 b | 1.73 ± 0.02 a | 1.77 ± 0.03 a | 1.58 ± 0.03 c | 1.61 ± 0.02 cb | 1.58 ± 0.03 cb | 0.0168 |
| SCFA 4 | 8.33 ± 0.04 c | 8.39 ± 0.03 c | 8.51 ± 0.04 b | 8.60 ± 0.06 b | 8.72 ± 0.04 a | 8.81 ± 0.05 a | 0.0007 |
| MCFA 5 | 21.13 ± 0.14 a | 20.78 ± 0.11 b | 20.39 ± 0.16 c | 20.06 ± 0.21 cd | 20.24 ± 0.15 cd | 19.88 ± 0.18 d | 0.0053 |
| LCFA 6 | 65.33 ± 0.2 d | 65.12 ± 0.16 de | 64.67 ± 0.24 e | 67.68 ± 0.31 b | 66.88 ± 0.22 ac | 66.53 ± 0.27 c | 0.0048 |
| Milk income (NZD/cow/day) | 12.58 ± 0.26 b | 12.50 ± 0.21 b | 11.74 ± 0.30 c | 14.59 ± 0.39 a | 14.67 ± 0.28 a | 14.61 ± 0.34 a | 0.2208 |
| SFC10 7 | 69.83 ± 0.49 a | 67.49 ± 0.39 b | 65.03 ± 0.57 c | 70.45 ± 0.73 a | 69.21 ± 0.53 a | 66.89 ± 0.65 b | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanjayaranj, I.; MacGibbon, A.K.H.; Holroyd, S.E.; Janssen, P.W.M.; Blair, H.T.; Lopez-Villalobos, N. Association of Single Nucleotide Polymorphism in the DGAT1 Gene with the Fatty Acid Composition of Cows Milked Once and Twice a Day. Genes 2023, 14, 767. https://doi.org/10.3390/genes14030767
Sanjayaranj I, MacGibbon AKH, Holroyd SE, Janssen PWM, Blair HT, Lopez-Villalobos N. Association of Single Nucleotide Polymorphism in the DGAT1 Gene with the Fatty Acid Composition of Cows Milked Once and Twice a Day. Genes. 2023; 14(3):767. https://doi.org/10.3390/genes14030767
Chicago/Turabian StyleSanjayaranj, Inthujaa, Alastair K. H. MacGibbon, Stephen E. Holroyd, Patrick W. M. Janssen, Hugh T. Blair, and Nicolas Lopez-Villalobos. 2023. "Association of Single Nucleotide Polymorphism in the DGAT1 Gene with the Fatty Acid Composition of Cows Milked Once and Twice a Day" Genes 14, no. 3: 767. https://doi.org/10.3390/genes14030767