
Floral Development Stage-Specific Transcriptomic Analysis Reveals the Formation Mechanism of Different Shapes of Ray Florets in Chrysanthemum

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. RNA-Seq, Functional Annotation, and Data Processing
2.3. Identification of Differentially Expressed Genes (DEGs)
2.4. Weighted Gene Co-Expression Correlation Network Analysis (WGCNA)
2.5. Real-Time Quantitative Polymerase Chain Reaction (qRT-PCR) Analysis
3. Results
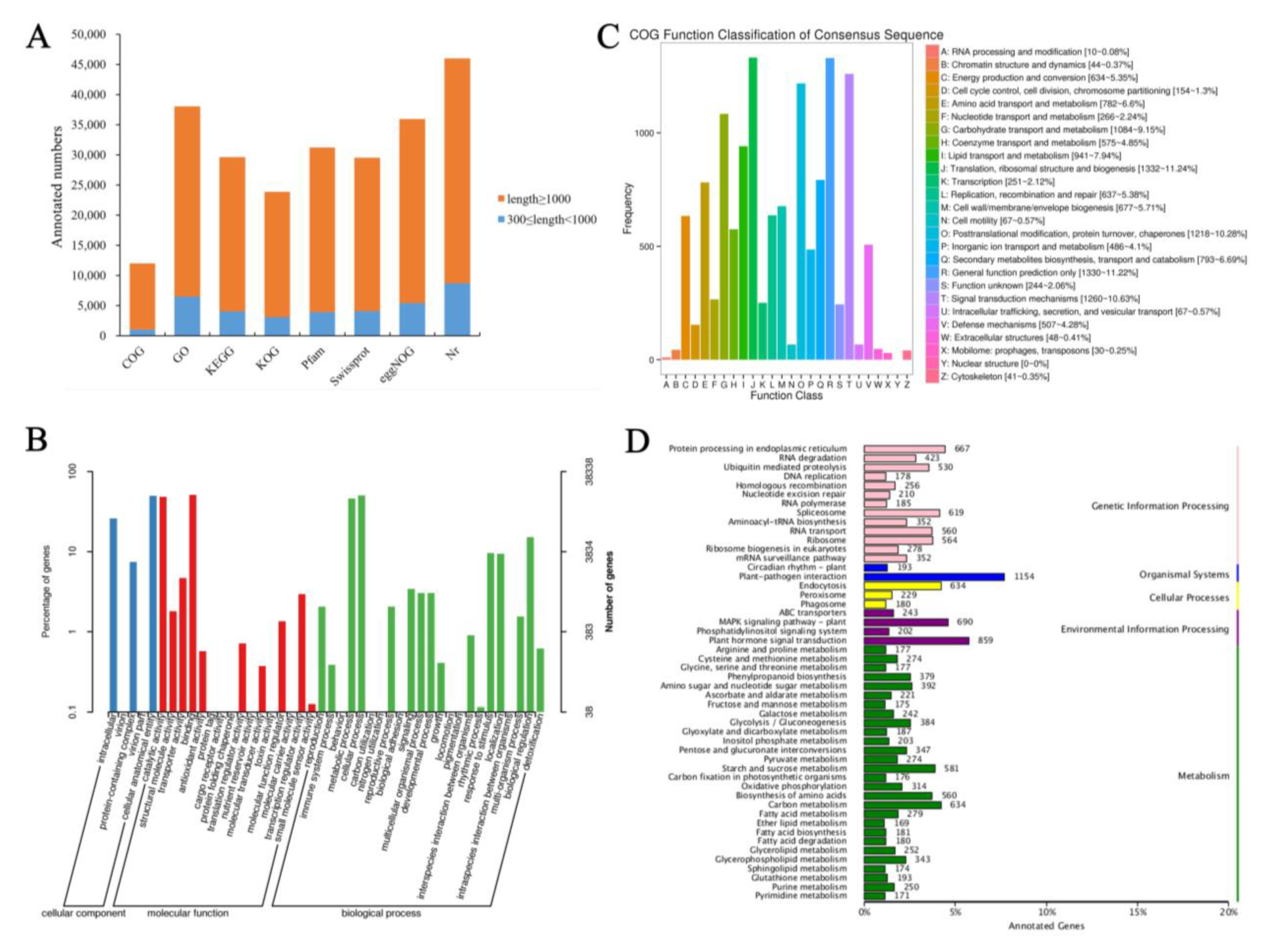
3.1. Sequencing, Assembly, and Functional Annotation of the Transcriptome
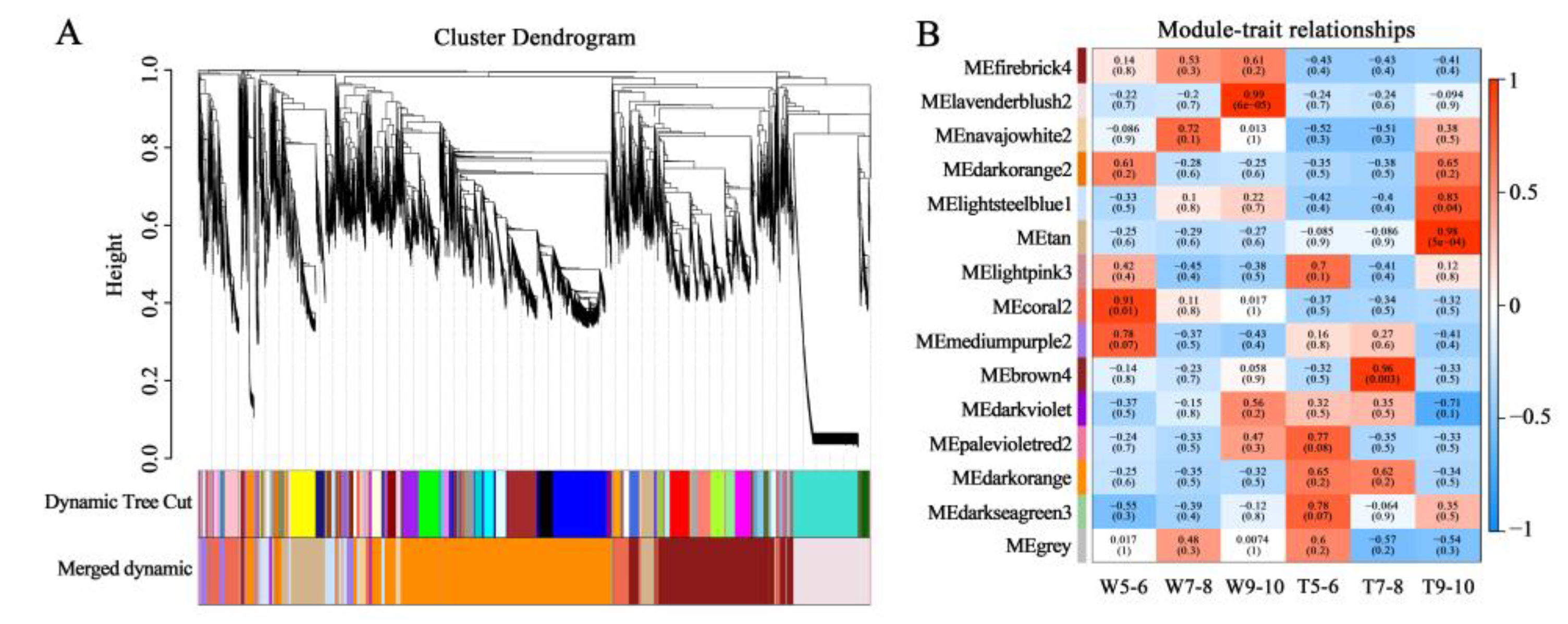
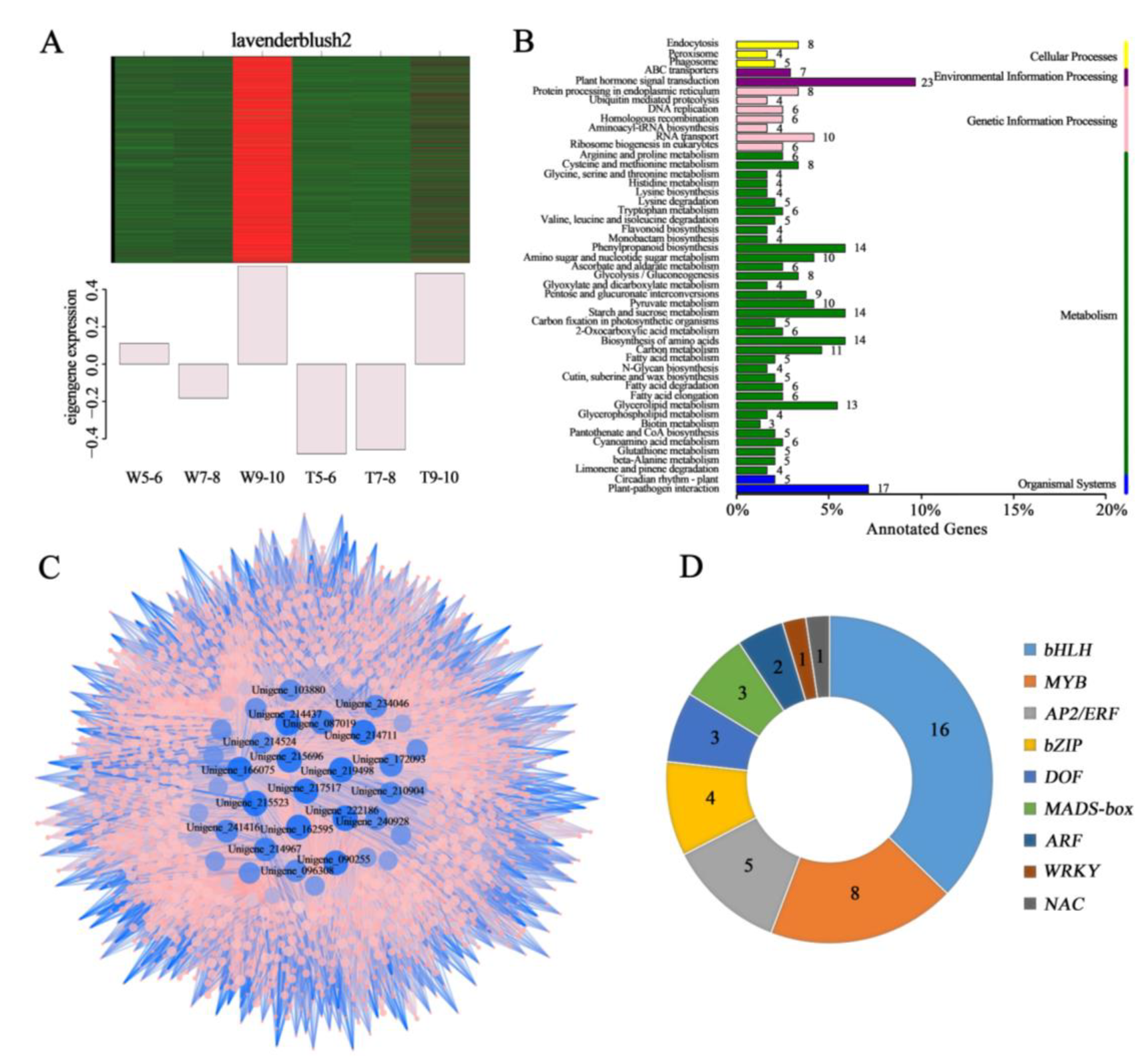
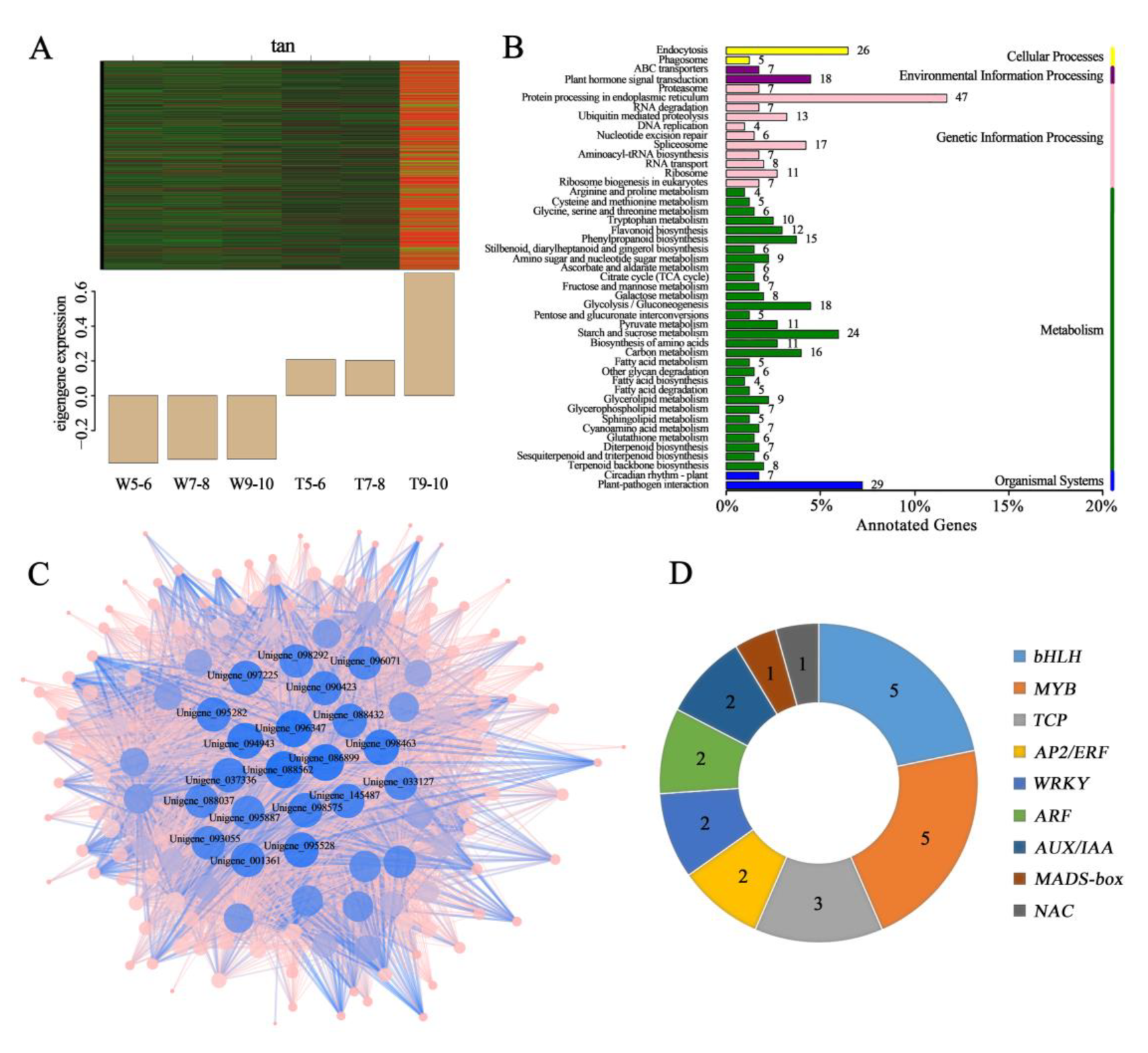
3.2. Screening for the Genes Related to the Development of Different Ray Floret Types through WGCNA
3.3. DEGs Identified by K-Means Cluster Analysis
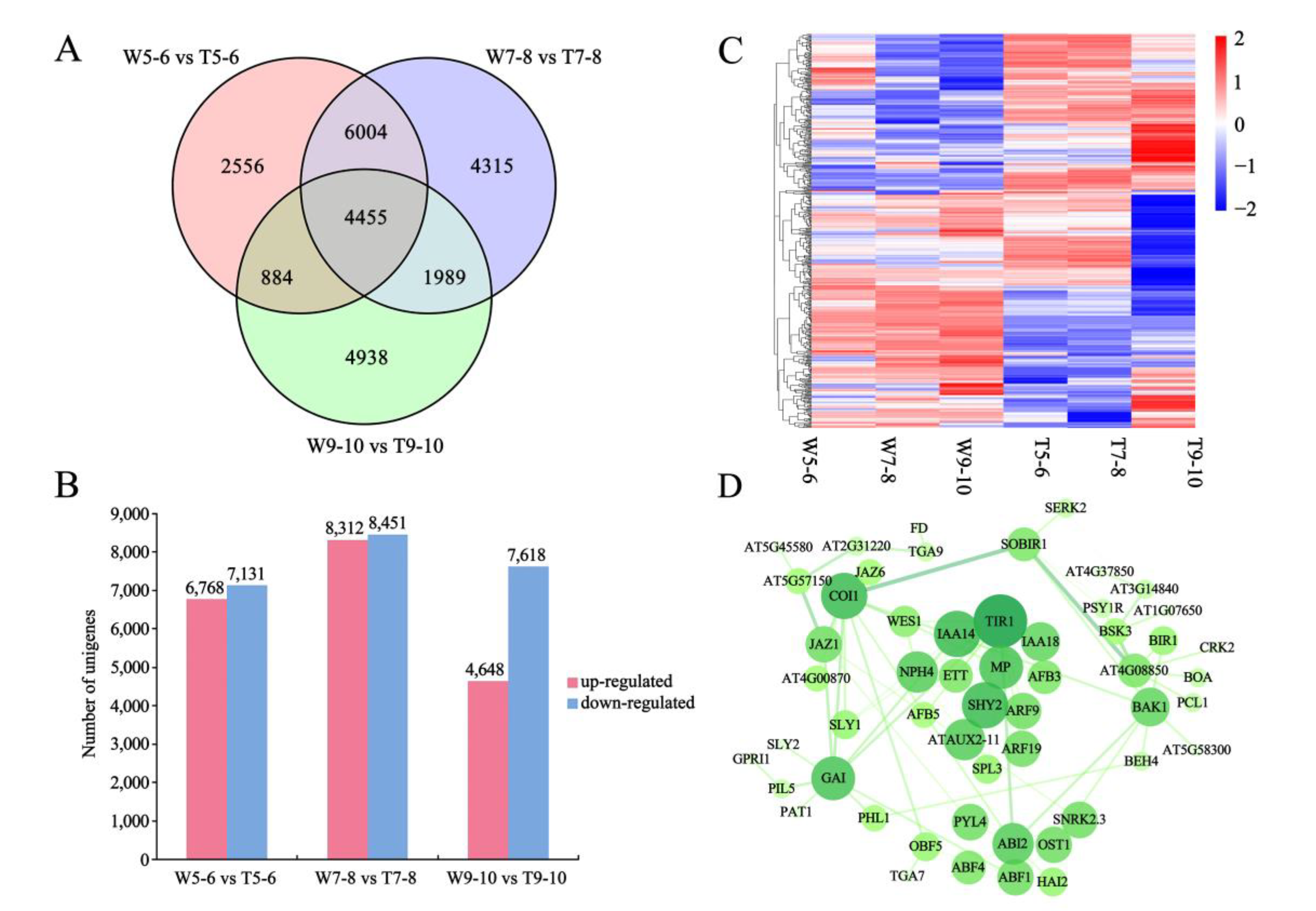
3.4. Screening for Candidate DEGs Regulating Morphological Differences in Ray Florets by Pairwise Comparison
3.5. Validation of Key DEGs Related to Morphological Difference in Ray Florets
4. Discussion
4.1. Auxin and Auxin-Related Genes Are Involved in Regulating the Formation of Different Ray Floret Types
4.2. Multiple Plant Hormones Are Involved in Regulating the Morphology of Ray Florets
4.3. MADS-box Genes Affect the Morphogenesis of Ray Florets
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nasri, F.; Zakizadeh, H.; Vafaee, Y.; Mozafari, A.A. In vitro mutagenesis of Chrysanthemum morifolium cultivars using ethylmethanesulphonate (EMS) and mutation assessment by ISSR and IRAP markers. Plant Cell Tissue Organ Cult. 2021, 149, 657–673. [Google Scholar] [CrossRef]
- Reynolds, J.; Tampion, J. Double Flowers: A Scientific Study; Pembridge Press: London, UK, 1983. [Google Scholar]
- Dejong, J.; Drennan, D.L. Genetic analysis in Chrysanthemum morifolium. II. Flower doubleness and ray floret corolla splitting. Euphytica 1984, 33, 465–470. [Google Scholar] [CrossRef]
- Song, X.B.; Gao, K.; Fan, G.X.; Zhao, X.G.; Liu, Z.L.; Dai, S.L. Quantitative classification of the morphological traits of ray florets in large-flowered Chrysanthemum. Hortscience 2018, 53, 1258–1265. [Google Scholar] [CrossRef] [Green Version]
- Shan, H.Y.; Cheng, J.; Zhang, R.; Yao, X.; Kong, H.Z. Developmental mechanisms involved in the diversification of flowers. Nat. Plants 2019, 5, 917–923. [Google Scholar] [CrossRef]
- Huang, T.; Irish, V.F. Temporal control of plant organ growth by TCP transcription factors. Curr. Biol. 2015, 25, 1765–1770. [Google Scholar] [CrossRef] [Green Version]
- Varaud, E.; Brioudes, F.; Szecsi, J.; Leroux, J.; Brown, S.; Perrot-Rechenmann, C.; Bendahmane, M. AUXIN RESPONSE FACTOR8 regulates Arabidopsis petal growth by interacting with the bHLH transcription factor BIGPETALp. Plant Cell 2011, 23, 973–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavallini-Speisser, Q.; Morel, P.; Monniaux, M. Petal cellular identities. Front. Plant Sci. 2021, 12, 745507. [Google Scholar] [CrossRef] [PubMed]
- Riglet, L.; Gatti, S.; Moyroud, E. Sculpting the surface: Structural patterning of plant epidermis. iScience 2021, 24, 103346. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.H.; Shan, H.Y.; Yao, X.; Cheng, J.; Jiang, Y.C.; Yin, X.F.; Kong, H.Z. Petal development and elaboration. J. Exp. Bot. 2022, 73, 3308–3318. [Google Scholar] [CrossRef] [PubMed]
- Song, X.B.; Zhao, X.G.; Fan, G.X.; Gao, K.; Dai, S.L.; Zhang, M.M.; Ma, C.F.; Wu, X.Y. Genetic analysis of the corolla tube merged degree and the relative number of ray florets in chrysanthemum (Chrysanthemum × morifolium Ramat.). Sci. Hortic. 2018, 242, 214–224. [Google Scholar] [CrossRef]
- Song, X.B.; Xu, Y.H.; Gao, K.; Fan, G.X.; Zhang, F.; Deng, C.Y.; Dai, S.L.; Huang, H.; Xin, H.G.; Li, Y.Y. High-density genetic map construction and identification of the locus controlling flower-type traits in Chrysanthemum (Chrysanthemum × morifolium Ramat.). Hortic. Res. 2020, 7, 108. [Google Scholar] [CrossRef] [PubMed]
- Kloos, W.E.; George, C.G.; Sorge, L.K. Inheritance of the flower types of Gerbera hybrida. J. Am. Soc. Hortic. Sci. 2004, 129, 802–810. [Google Scholar]
- Chapman, M.A.; Tang, S.; Draeger, D.; Nambeesan, S.; Shaffer, H.; Barb, J.B.; Knapp, S.J.; Burke, J.M. Genetic analysis of floral symmetry in Van Gogh’s sunflowers reveals independent recruitment of CYCLOIDEA genes in the Asteraceae. PLoS Genet. 2012, 8, e1002628. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Cui, M.L.; Cubas, P.; Gillies, A.; Lee, K.; Chapman, M.A.; Abbott, R.J.; Coen, E. Regulatory genes control a key morphological and ecological trait transferred between species. Science 2008, 322, 1116–1119. [Google Scholar] [CrossRef] [PubMed]
- Fambrini, M.; Salvini, M.; Pugliesi, C. A transposon-mediate inactivation of a CYCLOIDEA-like gene originates polysymmetric and androgynous ray flowers in Helianthus annuus. Genetica 2011, 139, 1521–1529. [Google Scholar] [CrossRef]
- Fambrini, M.; Bellanca, M.; Costa Muñoz, M.C.; Usai, G.; Cavallini, A.; Pugliesi, C. Ligulate inflorescence of Helianthus × multiflorus, cv. Soleil d’Or, correlates with a mis-regulation of a CYCLOIDEA gene characterised by insertion of a transposable element. Plant Biol. 2018, 20, 956–967. [Google Scholar] [PubMed]
- Huang, D.; Li, X.W.; Sun, M.; Zhang, T.X.; Pan, H.T.; Cheng, T.R.; Wang, J.; Zhang, Q.X. Identification and characterization of CYC-like genes in regulation of ray floret development in Chrysanthemum morifolium. Front. Plant Sci. 2016, 7, 1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Shen, C.Z.; Guo, Y.P.; Rao, G.Y. Patterning the Asteraceae capitulum duplications and differential expression of the flower symmetry CYC2-like genes. Front. Plant Sci. 2018, 9, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.Z.; Chen, J.; Zhang, C.J.; Rao, G.Y.; Guo, Y.P. Dysfunction of CYC2g is responsible for the evolutionary shift from radiate to disciform flowerheads in the Chrysanthemum group (Asteraceae: Anthemideae). Plant J. 2021, 106, 1024–1038. [Google Scholar] [CrossRef]
- Broholm, S.K.; TäHtiharju, S.; Laitinen, R.A.E.; Albert, V.A.; Teeri, T.H.; Elomaa, P. A TCP domain transcription factor controls flower type specification along the radial axis of the Gerbera (Asteraceae) inflorescence. Proc. Natl. Acad. Sci. USA 2008, 105, 9117–9122. [Google Scholar] [CrossRef] [Green Version]
- Bello, M.A.; Pilar, C.; Álvarez, I.; Sanjuanbenito, G.; Fuertes-Aguilar, J. Evolution and expression patterns of CYC/TB1 genes in Anacyclus: Phylogenetic insights for floral symmetry genes in Asteraceae. Front. Plant Sci. 2017, 8, 589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Cieslak, M.; Owens, A.; Wang, F.; Broholm, S.K.; Teeri, T.H.; Elomaa, P.; Prusinkiewicz, P. Phyllotactic patterning of gerbera flower heads. Proc. Natl. Acad. Sci. USA 2021, 118, e2016304118. [Google Scholar] [CrossRef]
- Zoulias, N.; Duttke, S.H.C.; Garcês, H.; Spencer, V.; Kim, M. The role of auxin in the pattern formation of the Asteraceae flower head (capitulum). Plant Physiol. 2019, 179, 391–401. [Google Scholar] [CrossRef] [Green Version]
- Cheng, P.P.; Liu, Y.N.; Yang, Y.M.; Chen, H.; Cheng, H.; Hu, Q.; Zhang, Z.X.; Gao, J.J.; Zhang, J.X.; Ding, L.; et al. CmBES1 is a regulator of boundary formation in chrysanthemum ray florets. Hortic. Res. 2020, 7, 129. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.X.; Ding, L.; Jiang, J.F.; Jia, D.W.; Li, S.; Jin, L.; Zhao, W.Q.; Zhang, X.; Song, A.P.; Chen, S.M.; et al. The TIFY family protein CmJAZ1-like negatively regulates petal size via interaction with the bHLH transcription factor CmBPE2 in Chrysanthemum morifolium. Plant J. 2022, 122, 1489–1506. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.L.; Wang, W.K.; Li, M.X.; Xu, Y.X. Phylogenetic relationship of Dendranthema (DC.) des moul. revealed by fluorescent in situ hybridization. J. Integr. Plant Biol. 2005, 47, 783–791. [Google Scholar]
- Chen, J.Y.; Wang, C.Y.; Zhao, H.E.; Zhou, J. The Origin of Garden Chrysanthemum; Anhui Science & Technology Press: Hefei, China, 2012; ISBN 978-7-5337-5691-8. [Google Scholar]
- Luo, C.; Chen, D.L.; Cheng, X.; Liu, H.; Li, Y.H.; Huang, C.L. SSR Analysis of genetic relationship and classification in Chrysanthemum germplasm collection. Hortic. Plant J. 2018, 4, 73–82. [Google Scholar] [CrossRef]
- Pu, Y.; Huang, H.; Wen, X.H.; Lu, C.F.; Zhang, B.H.; Gu, X.Q.; Qi, S.; Fan, G.X.; Wang, W.K.; Dai, S.L. Comprehensive transcriptomic analysis provides new insights into the mechanism of ray floret morphogenesis in Chrysanthemum. BMC Genom. 2020, 21, 728. [Google Scholar] [CrossRef]
- Luo, J.; Wang, H.; Chen, S.J.; Ren, S.J.; Fu, H.S.; Li, R.R.; Wang, C.Y. CmNAC73 mediates the formation of green color in chrysanthemum flowers by directly activating the expression of chlorophyll biosynthesis genes HEMA1 and CRD1. Genes 2021, 12, 704. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, D.Q.; et al. Full length transcriptome assembly from RNA Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [PubMed] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of shor tDNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewer, C.N. RSEM: Accurate transcript quantification from RNA Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.F.; Pu, Y.; Liu, Y.T.; Li, Y.J.; Qu, J.P.; Huang, H.; Dai, S.L. Comparative transcriptomics and weighted gene co-expression correlation network analysis (WGCNA) reveal potential regulation mechanism of carotenoid accumulation in Chrysanthemum × morifolium. Plant Physiol. Biochem. 2019, 142, 415–428. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2013, 13, 2498–2504. [Google Scholar] [CrossRef]
- Qi, S.; Yang, L.W.; Wen, X.H.; Hong, Y.; Song, X.B.; Zhang, M.M.; Dai, S.L. Reference gene selection for RT-qPCR analysis of flower development in Chrysanthemum morifolium and Chrysanthemum lavandulifolium. Front. Plant Sci. 2016, 7, 287. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Blakeslee, J.J.; Rossi, T.S.; Kriechbaumer, V. Auxin biosynthesis: Spatial regulation and adaptation to stress. J. Exp. Bot. 2019, 70, 5041–5049. [Google Scholar] [CrossRef]
- Báez, R.R.; Nemhauser, J.L. Expansion and innovation in auxin signaling: Where do we grow from here? Development 2021, 148, dev187120. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.W.; Jacobs, B.; Cole, M.; Comelli, P.; Werr, W. DORNRÖSCHEN-LIKE expression marks Arabidopsis floral organ founder cells and precedes auxin response maxima. Plant Mol. Biol. 2011, 76, 171–185. [Google Scholar] [CrossRef]
- Heisler, M.G.; Ohno, C.; Das, P.; Sieber, P.; Reddy, G.V.; Long, J.A.; Meyerowitz, E.M. Patterns of auxin transport and gene expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem. Curr. Biol. 2005, 15, 1899–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.F.; Dai, X.H.; Zhao, Y.D. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Genes Dev. 2006, 20, 1790–1799. [Google Scholar] [CrossRef] [Green Version]
- Pekker, I.; Alvarez, J.P.; Eshed, Y. Auxin response factors mediate Arabidopsis organ asymmetry via modulation of KANADI activity. Plant Cell 2005, 17, 2899–2910. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.G.; Dai, X.H.; De-Paoli, H.; Cheng, Y.F.; Takebayashi, Y.; Kasahara, H.; Kamiya, Y.; Zhao, Y.D. Auxin overproduction in shoots cannot rescue auxin deficiencies in Arabidopsis roots. Plant Cell Physiol. 2014, 55, 1072–1079. [Google Scholar] [CrossRef] [Green Version]
- Bennett, S.R.M.; Alvarez, J.; Bossinger, G.; Smyth, D.R. Morphogenesis in PINOID mutants of Arabidopsis thaliana. Plant J. 1995, 8, 505–520. [Google Scholar] [CrossRef]
- Brewer, P.B.; Howles, P.A.; Dorian, K.; Griffith, M.E.; Ishida, T.; Kaplan-Levy, R.N.; Kilinc, A.; Smyth, D.R. PETAL LOSS, a trihelix transcription factor gene, regulates perianth architecture in the Arabidopsis flower. Development 2004, 131, 4035–4045. [Google Scholar] [CrossRef] [Green Version]
- Ding, B.Q.; Xia, R.; Lin, Q.S.; Gurung, V.; Sagawa, J.M.; Stanley, L.E.; Strobel, M.; Diggle, P.K.; Meyers, B.C.; Yuan, Y.W. Developmental genetics of corolla tube formation: Role of the tasiRNA-ARF pathway and a conceptual model. Plant Cell 2020, 32, 3452–3468. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xie, Q.; Chua, N.H. The Arabidopsis auxin-inducible gene ARGOS controls lateral organ size. Plant Cell 2003, 15, 1951–1961. [Google Scholar] [CrossRef] [Green Version]
- Mizukami, Y.; Fischer, R.L. Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 942–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horstman, A.; Willemsen, V.; Boutilier, K.; Heidstra, R. AINTEGUMENTA-LIKE proteins: Hubs in a plethora of networks. Trends Plant Sci. 2014, 19, 146–157. [Google Scholar] [CrossRef]
- Krizek, B.A. AINTEGUMENTA-LIKE genes have partly overlapping functions with AINTEGUMENTA but make distinct contributions to Arabidopsis thaliana flower development. J. Exp. Bot. 2015, 66, 4537–4549. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, N.; Wu, M.F.; Winter, C.M.; Berns, M.C.; Nole-Wilson, S.; Yamaguchi, A.; Coupland, G.; Krizek, B.A.; Wagner, D. A molecular framework for auxin-mediated initiation of flower primordia. Dev. Cell 2013, 24, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Krizek, B.A. AINTEGUMENTA and AINTEGUMENTA-LIKE6 act redundantly to regulate Arabidopsis floral growth and patterning. Plant Physiol. 2009, 150, 1916–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.F.; Zhang, W.B.; Zhang, L.L.; Li, N.; Peng, J.Z.; Wang, Y.Q.; Zhong, C.M.; Yang, Y.P.; Sun, S.L.; Liang, S.; et al. Transcriptomic insights into antagonistic effects of gibberellin and abscisic acid on petal growth in Gerbera hybrida. Front. Plant Sci. 2015, 6, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.; Han, M.X.; Yao, W.; Wang, Y.Q. Transcriptome analysis reveals the regulation of brassinosteroids on petal growth in Gerbera hybrida. PeerJ 2017, 5, e3382. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.P.; Li, L.F.; Huang, Y.H.; Wang, Y.Q.; Zhang, W.B.; Zheng, R.Y.; Zhong, C.M.; Wang, X.J. GhWIP2, a WIP zinc finger protein, suppresses cell expansion in Gerbera hybrida by mediating crosstalk between gibberellin, abscisic acid, and auxin. New Phytolpgist 2018, 219, 728–742. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Wang, H.B.; Ding, L.; Song, A.P.; Shen, F.; Jiang, J.F.; Chen, S.M.; Chen, F.D. Transcriptomic and hormone analyses reveal mechanisms underlying petal elongation in Chrysanthemum morifolium ‘Jinba’. Plant Mol. Biol. 2017, 93, 593–606. [Google Scholar] [CrossRef]
- Alvarez-Buylla, E.R.; Benítez, M.; Corvera-Poiré, A.; Chaos Cador, A.; de Folter, S.; Gamboa de Buen, A.; Garay-Arroyo, A.; Garcia-Ponce, B.; Jaimes-Miranda, F.; Perez-Ruiz, R.; et al. Flower development. The Arabidopsis Book 2010, 8, e0127. [Google Scholar] [CrossRef] [Green Version]
- Theißen, G.; Melzer, R.; Florian, R. MADS-domain transcription factors and the floral quartet model of flower development: Linking plant development and evolution. Development 2016, 143, 3259–3271. [Google Scholar] [CrossRef] [Green Version]
- Irish, V. The ABC model of floral development. Curr. Biol. 2017, 27, R887–R890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laitinen, R.A.; Broholm, S.; Albert, V.A.; Teeri, T.H.; Elomaa, P. Patterns of MADS-box gene expression mark flower-type development in Gerbera hybrida (Asteraceae). BMC Plant Biol. 2006, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.; Kotilainen, M.; Pöllänen, E.; Mehto, M.; Elomaa, P.; Helariutta, Y.; Albert, V.A.; Teeri, T.H. Organ identity genes and modified patterns of flower development in Gerbera hybrida (Asteraceae). Plant J. Ceel Mol. Biol. 1999, 17, 51–62. [Google Scholar] [CrossRef]
- Uimari, A.; Kotilainen, M.; Elomaa, P.; Yu, D.; Albert, V.A.; Teeri, T.H. Integration of reproductive meristem fates by a SEPALLATA-like MADS-box gene. Proc. Natl. Acad. Sci. USA 2004, 101, 15817–15822. [Google Scholar] [CrossRef] [Green Version]
- Elomaa, P.; Zhao, Y.F.; Zhang, T. Flower heads in Asteraceae-recruitment of conserved developmental regulators to control the flower-like inflorescence architecture. Hortic. Res. 2018, 5, 36. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhao, Y.F.; Juntheikki, I.; Mouhu, K.; Broholm, S.K.; Rijpkema, A.S.; Kins, L.; Lan, T.Y.; Albert, V.A.; Teeri, T.H.; et al. Dissecting functions of SEPALLATA-like MADS box genes in patterning of the pseudanthial inflorescence of Gerbera hybrida. New Phytol. 2017, 216, 939–954. [Google Scholar] [CrossRef] [Green Version]
- Broholm, S.K.; Pöllänen, E.; Ruokolainen, S.; Tähtiharju, S.; Kotilainen, M.; Albert, V.A.; Elomaa, P.; Teeri, T.H. Functional characterization of B class MADS-box transcription factors in Gerbera hybrida. J. Exp. Bot. 2010, 61, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.H.; Wang, J.H.; Gu, K.D.; Zhang, P.; Zhang, C.S.; Hu, D.G.; Ma, F.F. New insights into the role of MADS-box transcription factor gene CmANR1 on root and shoot development in chrysanthemum (Chrysanthemum morifolium). BMC Plant Biol. 2021, 21, 79. [Google Scholar] [CrossRef]
- Sasaki, K.; Yoshioka, S.; Aida, R.; Ohtsubo, N. Production of petaloid phenotype in the reproductive organs of compound flowerheads by the co-suppression of class-C genes in hexaploid Chrysanthemum morifolium. Planta 2021, 253, 100. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jia, Y.; Chai, Y.H.; Wang, S.; Chen, H.X.; Zhou, X.M.; Huang, C.L.; Guo, S.; Chen, D.L. Whole-transcriptome analysis of differentially expressed genes between ray and disc florets and identification of flowering regulatory genes in Chrysanthemum morifolium. Front. Plant Sci. 2022, 13, 947331. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.W.; Huang, J.L.; Pu, Y.; Niu, Y.J.; Zhang, M.M.; Dai, S.L.; Huang, H. Transcriptomic analysis reveals the formation mechanism of anemone-type flower in chrysanthemum. BMC Genom. 2022, 23, 846. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.H.; Qi, S.; Huang, H.; Wu, X.Y.; Zhang, B.H.; Fan, G.X.; Yang, L.W.; Hong, Y.; Dai, S.L. The expression and interactions of ABCE-class and CYC2-like genes in the capitulum development of Chrysanthemum lavandulifolium and C. × morifolium. Plant Growth Regul. 2019, 88, 205–214. [Google Scholar] [CrossRef]
- Ding, L.; Song, A.P.; Zhang, X.; Li, S.; Su, J.S.; Xia, W.K.; Zhao, K.K.; Zhao, W.Q.; Guan, Y.X.; Fang, W.M.; et al. The core regulatory networks and hub genes regulating flower development in Chrysanthemum morifolium. Plant Mol. Biol. 2020, 103, 669–688. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| #ID | FPKM of Each Sample | Annotation | |||||
|---|---|---|---|---|---|---|---|
| W5-6 | W7-8 | W9-10 | T5-6 | T7-8 | T9-10 | ||
| ARF | |||||||
| Unigene_093663 | 0.97 | 1.22 | 0.77 | 2.23 | 1.57 | 5.49 | Auxin response factor 19 |
| Unigene_149292 | 10.00 | 10.42 | 11.45 | 29.13 | 28.39 | 39.25 | Auxin response factor |
| Unigene_237143 | 20.92 | 23.70 | 23.73 | 13.30 | 15.75 | 10.88 | Auxin response factor 3 |
| Unigene_174210 | 2.89 | 3.65 | 3.37 | 8.14 | 11.27 | 11.45 | Auxin response factor 5 |
| AUX/IAA | |||||||
| Unigene_034051 | 4.45 | 2.25 | 1.22 | 9.58 | 10.39 | 3.79 | Auxin-responsive protein |
| Unigene_085389 | 3.19 | 2.30 | 3.80 | 5.67 | 4.32 | 9.10 | Auxin-induced protein 22D |
| Unigene_009970 | 0.90 | 2.88 | 1.75 | 1.44 | 0.68 | 7.70 | Indole-3-acetic acid 14 |
| AP2/ERF | |||||||
| Unigene_168891 | 3.48 | 7.13 | 5.40 | 5.95 | 5.83 | 0.86 | AP2-like ethylene-responsive transcription factor |
| Unigene_209218 | 2.23 | 5.12 | 4.72 | 0.18 | 0.18 | 1.09 | AP2/ERF protein |
| Unigene_033078 | 0.17 | 0.11 | 0.06 | 1.72 | 2.00 | 0.66 | AP2/ERF protein |
| Unigene_053843 | 48.23 | 3.91 | 0.33 | 29.64 | 21.68 | 2.11 | AP2/ERF protein |
| Unigene_173391 | 22.79 | 0.36 | 0.23 | 5.68 | 4.68 | 1.60 | Ethylene-responsive transcription factor 5 |
| Unigene_237527 | 0.34 | 0.32 | 7.63 | 0.40 | 1.07 | 0.95 | Ethylene-responsive transcription factor 14 |
| MADS-box | |||||||
| Unigene_086333 | 0.40 | 0.00 | 0.46 | 0.00 | 0.00 | 2.04 | MADS-box protein defh21 |
| Unigene_240064 | 7.67 | 43.22 | 131.43 | 105.67 | 180.61 | 348.32 | MADS-box transcription factor CDM77 |
| Unigene_164772 | 29.55 | 75.39 | 88.59 | 0.71 | 11.79 | 20.35 | MADS-box transcription factor CDM37 |
| Unigene_007650 | 6.53 | 5.91 | 3.62 | 6.83 | 6.39 | 7.54 | flower development transporter AP3 |
| Unigene_209803 | 0.06 | 1.60 | 2.96 | 6.58 | 7.36 | 13.96 | GRCD5 protein/SEP3 |
| Unigene_176186 | 7.27 | 8.76 | 8.90 | 1.39 | 1.24 | 1.54 | Flower development transporter AG2 |
| TCP | |||||||
| Unigene_240675 | 1.55 | 2.38 | 2.02 | 0.60 | 1.28 | 6.74 | Flower asymmetry transporter CYC2b |
| Unigene_166758 | 0.64 | 1.47 | 1.31 | 0.09 | 0.49 | 7.18 | Flower asymmetry transporter CYC2d |
| Unigene_039861 | 2.04 | 2.82 | 2.26 | 0.96 | 1.95 | 6.72 | Flower asymmetry transporter CYC2e |
| Unigene_168575 | 2.00 | 2.64 | 2.26 | 0.33 | 0.60 | 0.59 | Flower asymmetry transporter CYC2f |
| NAC | |||||||
| Unigene_154909 | 54.56 | 9.71 | 5.95 | 72.13 | 88.18 | 16.07 | NAC1 |
| Unigene_231589 | 3.87 | 4.57 | 4.49 | 1.56 | 1.54 | 1.95 | NAC transcription factor 25 |
| Unigene_096105 | 0.42 | 0.17 | 0.12 | 1.52 | 1.49 | 1.44 | No apical meristem protein |
| Unigene_037350 | 2.39 | 0.13 | 0.08 | 4.47 | 5.33 | 13.66 | No apical meristem protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pu, Y.; Liao, M.; Li, J.; Tian, Y.; Wang, Z.; Song, X.; Dai, S. Floral Development Stage-Specific Transcriptomic Analysis Reveals the Formation Mechanism of Different Shapes of Ray Florets in Chrysanthemum. Genes 2023, 14, 766. https://doi.org/10.3390/genes14030766
Pu Y, Liao M, Li J, Tian Y, Wang Z, Song X, Dai S. Floral Development Stage-Specific Transcriptomic Analysis Reveals the Formation Mechanism of Different Shapes of Ray Florets in Chrysanthemum. Genes. 2023; 14(3):766. https://doi.org/10.3390/genes14030766
Chicago/Turabian StylePu, Ya, Minling Liao, Junzhuo Li, Yuankai Tian, Zhongman Wang, Xiang Song, and Silan Dai. 2023. "Floral Development Stage-Specific Transcriptomic Analysis Reveals the Formation Mechanism of Different Shapes of Ray Florets in Chrysanthemum" Genes 14, no. 3: 766. https://doi.org/10.3390/genes14030766