
Coronatine-Based Gene Expression Changes Impart Partial Resistance to Fall Armyworm (Spodoptera frugiperda) in Seedling Maize
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Insect Materials
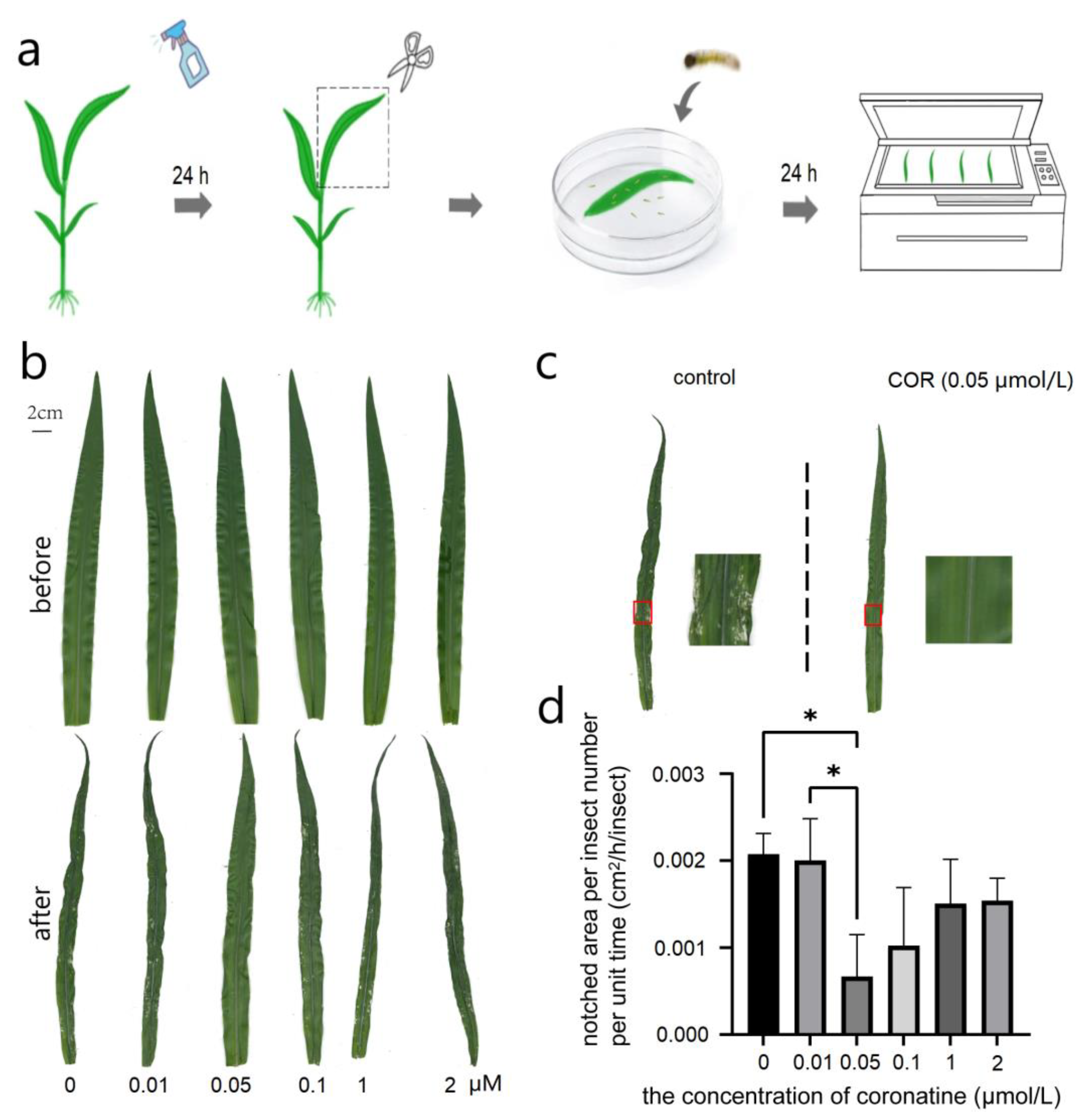
2.2. The Method of COR Treatment
2.3. Determination of the Feeding Amount of the Newly Hatched Larvae of S. frugiperda on the Leaves of Maize Sprayed with COR
2.4. Determination of Endogenous Plant Hormones in Maize Leaves
2.5. RNA Extraction and Library Preparing
2.6. RNA-Seq Data Analysis
2.7. The Analysis of Differential Expression
2.8. Functional Enrichment Analysis
3. Results
3.1. Insect Resistance of COR with Different Concentrations
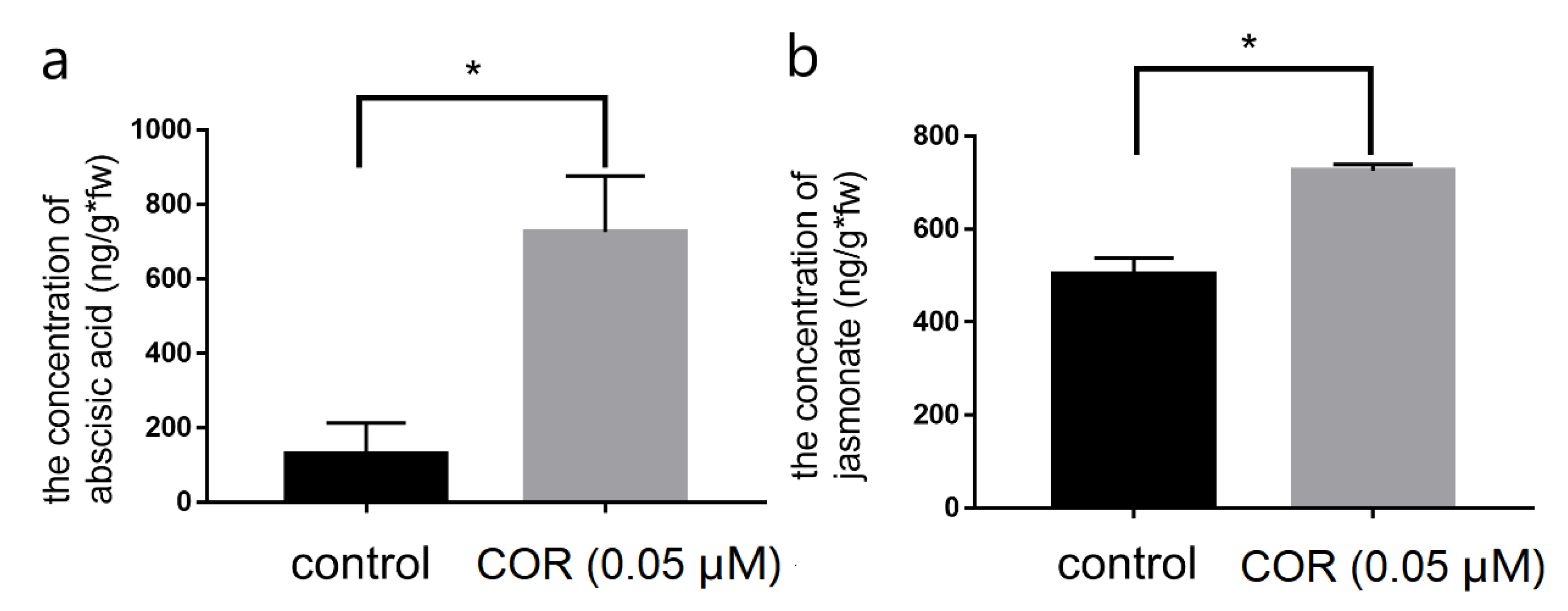
3.2. Changes in Endogenous Plant Hormones
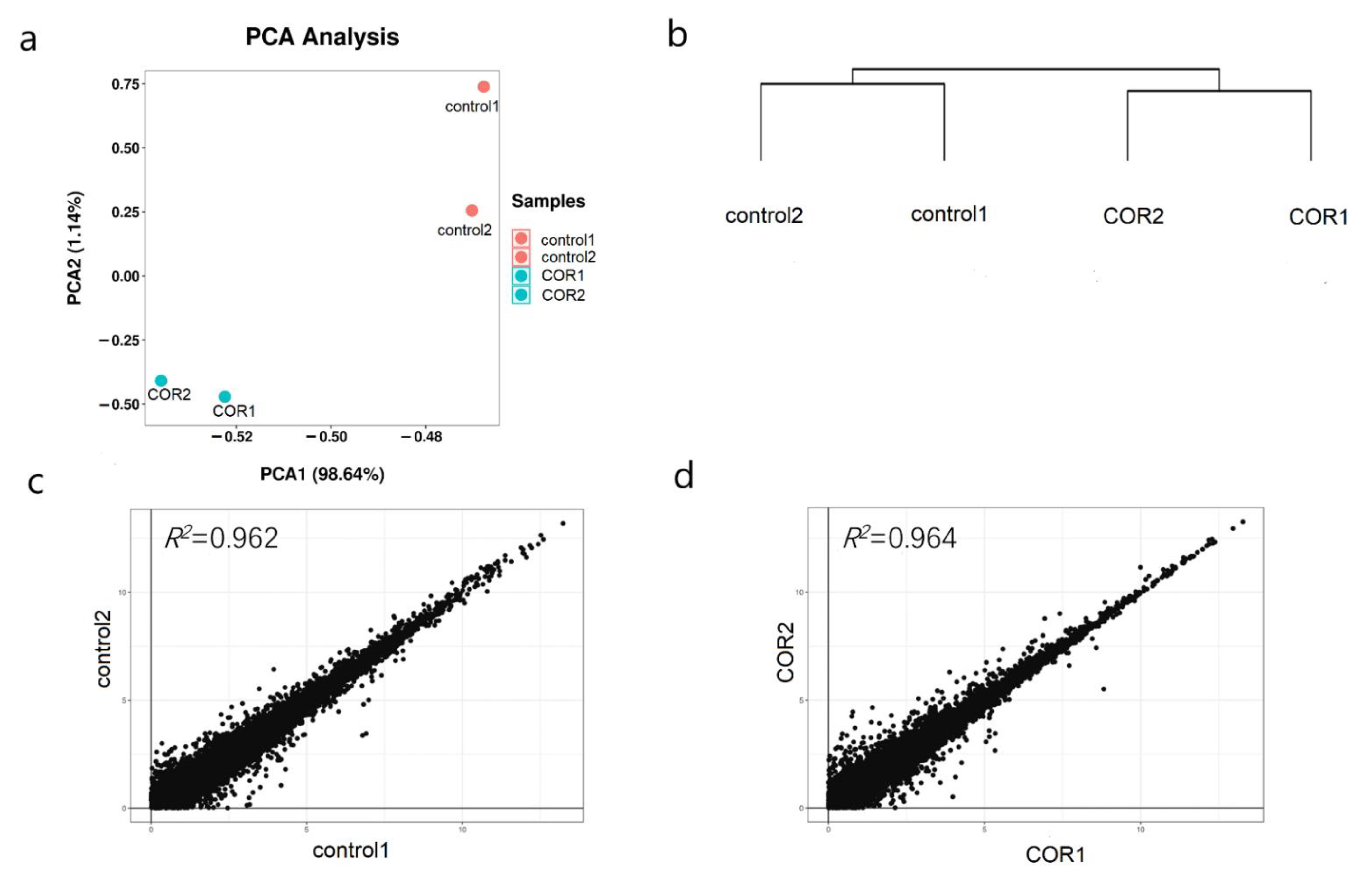
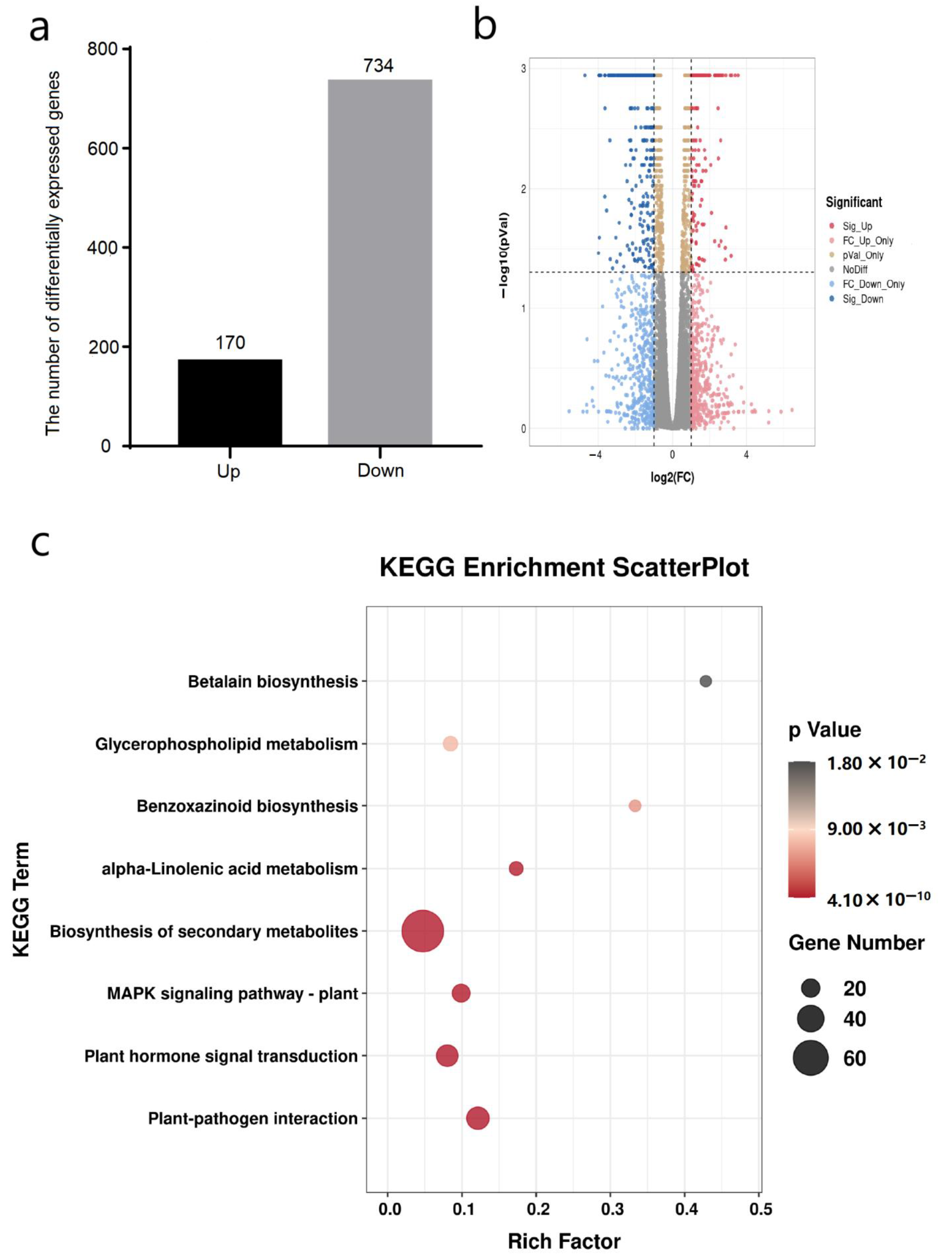
3.3. Overall Analysis of Transcriptome in Response to COR Treatment
3.4. Identification of Differentially Expressed Genes after COR Treatment
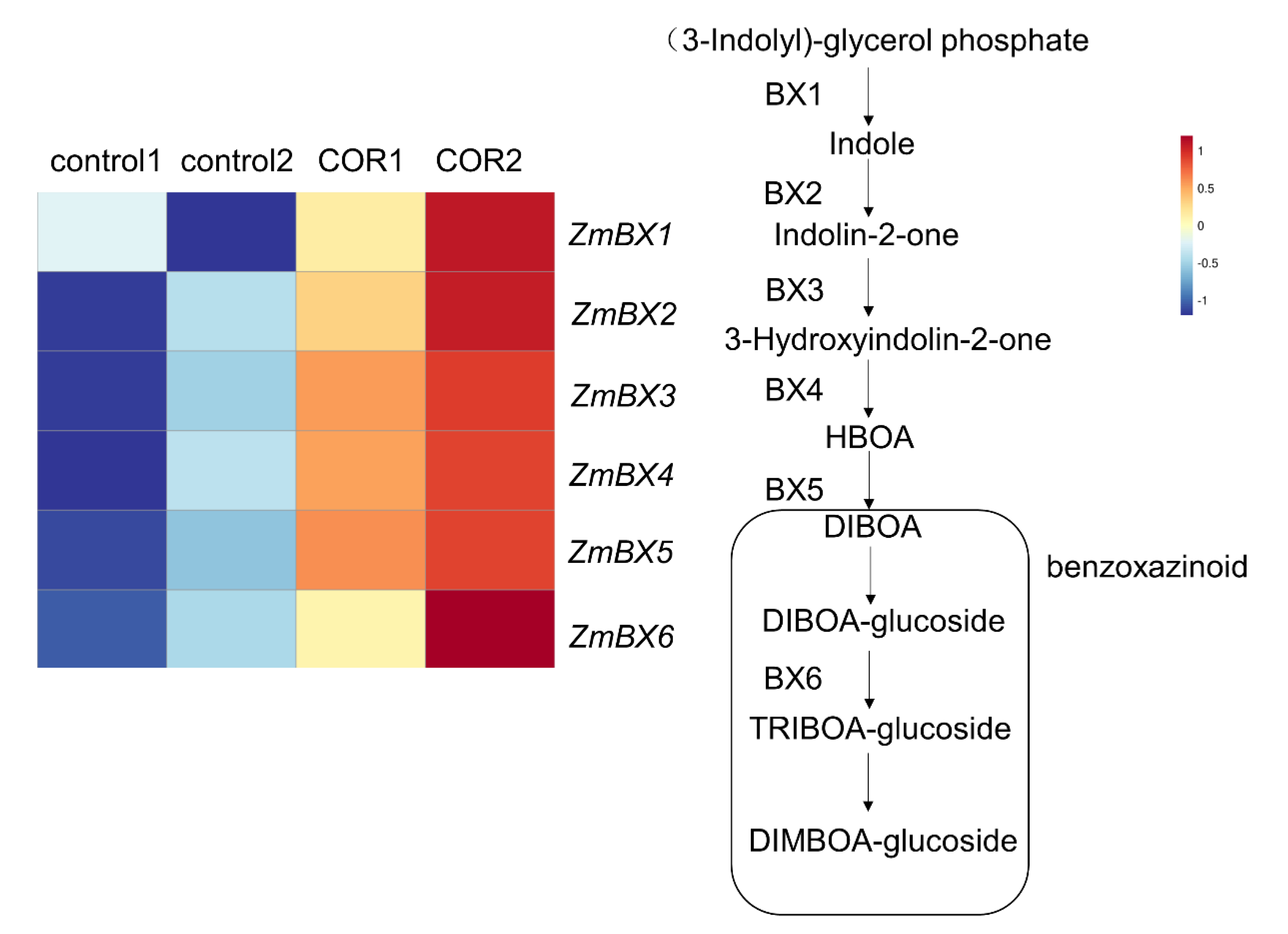
3.5. Expression of Genes Related to the Biosynthesis of Benzoxazines
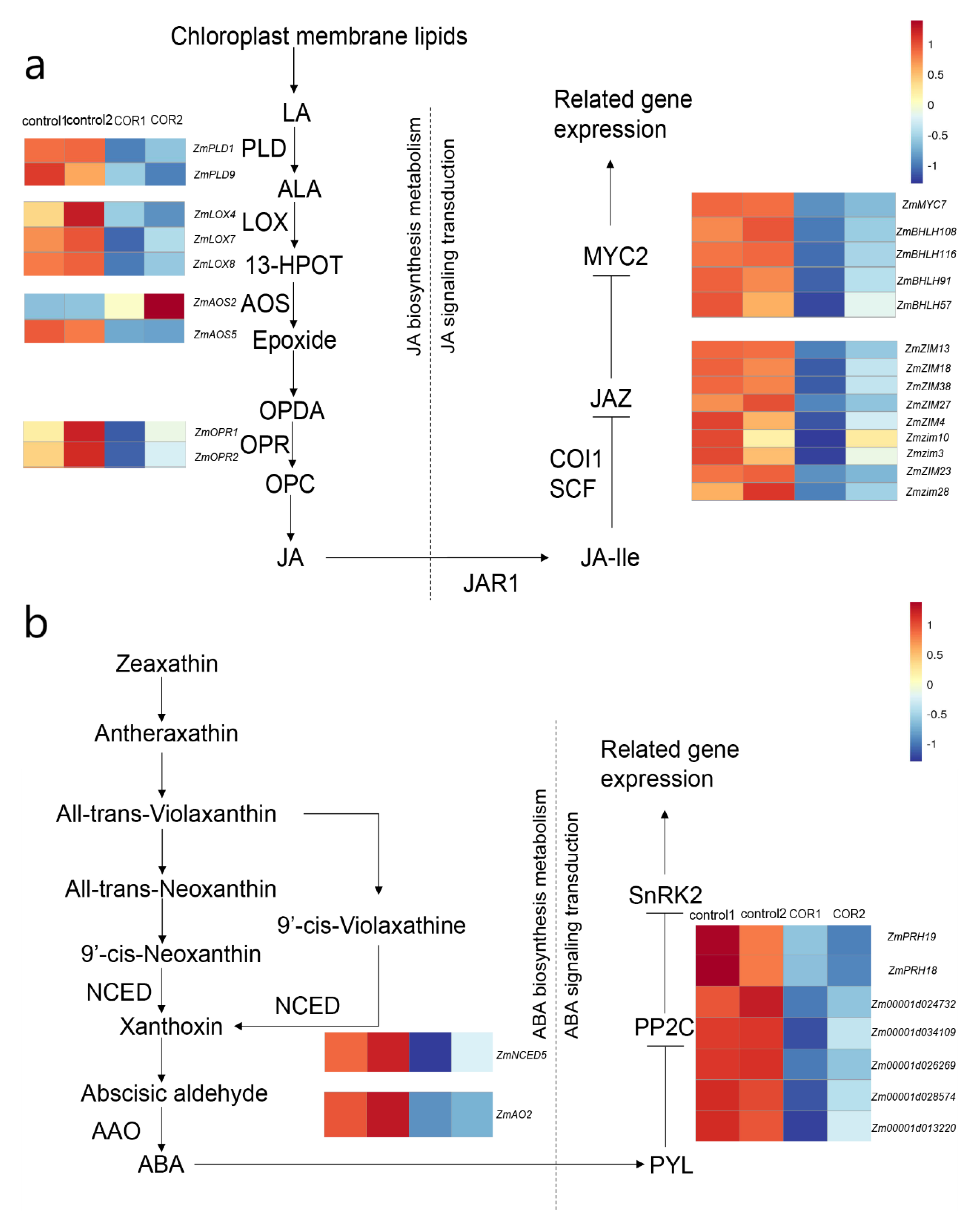
3.6. Expression of Genes Related to JA and ABA in Maize
4. Discussion
4.1. COR can Effectively Improve the Insect Resistance of Maize at the Seedling Stage
4.2. The Enhanced Synthesis of Benzoxazines May Be One of the Reasons for the Improvement of the Insect Resistance of Maize at the Seedling Stage
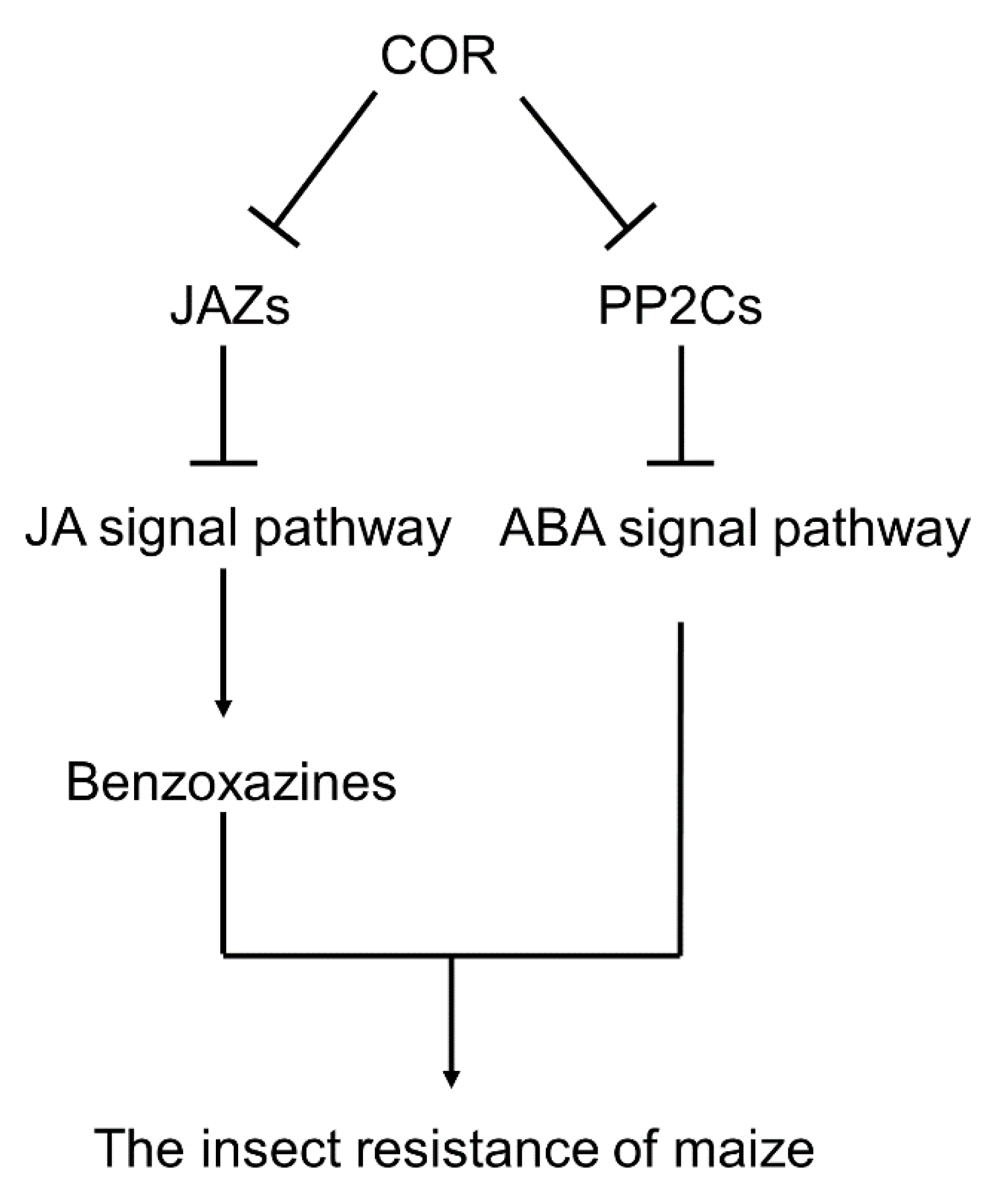
4.3. COR May Improve the Insect Resistance of Maize at the Seedling Stage through the ABA and JA Signal Pathways
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Yajun, C.; Yonglin, L.; Xingchuan, J.; Guirong, W.; Bin, Y. Advances in research on Spodoptera frugiperda and its functional genomics. Plant Prot. 2019, 45, 1–7. [Google Scholar]
- Xiangmin, W.; Yong, C.; Guojun, Y.; Kesen, Z.; Hui, X. On the host adaptability, population dynamics of Spodoptera frugiperda: Advances and prospect in efficient control. J. Environ. Entomol. 2020, 41, 42–51. [Google Scholar]
- Li, Q.; Men, X.; Jing, C.; Yu, Y.; Zhou, X.; Dai, X.; Li, S.; Li, L. Research progress in emergency prevention and control of Spodoptera frugiperda in China. Plant Prot. 2021, 47, 21–27, (in Chinese with English abstract). [Google Scholar]
- Montazano, D.G.; Specht, A.; Sosa-Gomez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–301. [Google Scholar] [CrossRef] [Green Version]
- Early, R.; Gonzalez-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. Neobiota 2018, 40, 25–50. [Google Scholar] [CrossRef] [Green Version]
- Wyckhuys, K.A.G.; O’Neil, R.J. Population dynamics of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) and associated arthropod natural enemies in Honduran subsistence maize. Crop Prot. 2006, 25, 1180–1190. [Google Scholar] [CrossRef]
- Murua, G.; Molina-Ochoa, J.; Coviella, C. Population dynamics of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) and its parasitoids in northwestern Argentina. Fla. Entomol. 2006, 89, 175–182. [Google Scholar] [CrossRef]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.L.; Shen, X.J.; Zhao, S.Y.; Jiang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm Spodoptera frugiperda invading into China. J. Entomol. Zool. Stud. 2018, 6, 2089–2901. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Liu, J.; Xie, M.C.; Li, Y.H.; Yang, J.J.; Zhang, M.L.; Qiu, K. Observation on law of diffusion damage of Spodoptera frugiperda in China in 2019. Plant Prot. 2019, 45, 10–19. [Google Scholar]
- Wu, K. Management strategies of fall armyworm (Spodoptera frugiperda) in China. Plant Prot. 2020, 46, 1–5. [Google Scholar]
- Li, Y.P.; Zhang, S.; Wang, X.J.; Xie, X.P.; Liang, P.; Zhang, L.; Gu, S.H.; Gao, X.W. Current status of insecticide resistance in Spodoptera frugiperda and strategies for its chemical control. Plant Prot. 2019, 45, 14–19. [Google Scholar]
- Gutierrez-Moreno, R.; Mota-Sanchez, D.; Blanco, C.A.; Whalon, M.E.; Teran-Santofimio, H.; Rodriguez-Maciel, J.C.; DiFonzo, C. Field-evolved resistance of the fall armyworm (Lepidoptera: Noctuidae) to synthetic insecticides in Puerto Rico and Mexico. J. Econ. Entomol. 2019, 112, 792–802. [Google Scholar] [CrossRef]
- Ichihara, A.; Shiraishi, K.; Sato, H.; Sakamura, S.; Nishiyama, K.; Sakai, R. The structure of coronatine. J. Am. Chem. Soc. 1977, 99, 636–637. [Google Scholar] [CrossRef]
- Katsir, L.; Schilmiller, A.L.; Staswick, P.E.; He, S.Y.; Howe, G.A. COI1 is a critical component of a receptor for jasmonate and the bacterial virulence factor coronatine. Proc. Natl. Acad. Sci. USA 2008, 105, 7100–7105. [Google Scholar] [CrossRef] [Green Version]
- Santner, A.; Estelle, M. Recent advances and emerging trends in plant hormone signalling. Nature 2009, 459, 1071–1078. [Google Scholar] [CrossRef]
- Gruelich, F.; Yoshihara, T.; Ichihara, A. Coronatine, a bacterial phytotoxin, acts as a stereospecific analog of jasmonate type signals in tomato cells and potato tissue. Plant Physiol. 1995, 147, 359–366. [Google Scholar] [CrossRef]
- Young, S.A.; Park, S.K.; Rodgers, C.; Mitchell, R.E.; Bender, C.L. Physical and functional characterization of the gene cluster encoding the polyketide phytotoxin coronatine in Pseudomonas syringae pv. glycinea. J. Bacteriol. 1992, 174, 1837–1843. [Google Scholar] [CrossRef] [Green Version]
- Weiler, E.W.; Kutchan, T.M.; Gorba, T.; Brodschelm, W.; Niesel, U.; Bublitz, F. The Pseudomonas phytotoxin coronatine mimics octadecanoid signaling molecules of higher plants. FEBS Lett. 1994, 345, 9–13. [Google Scholar] [CrossRef]
- Uppalapati, S.R.; Ayoubi, P.; Weng, H.; Palmer, D.A.; Mitchell, R.E.; Jones, W. The phytotoxin coronatine and methyl jasmonate impact multiple phytohormone pathways in tomato. Plant J. 2005, 42, 201–217. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Li, J.M.; Duan, L.S.; Zhang, M.C.; Li, Z.H. Regulation of bioregulator coronatine on the development of plant morphology in maize seedling. Chin. J. Pestic. Sci. 2015, 17, 401–408. [Google Scholar]
- Ren, Z.B.; Wang, X.; Tao, Q.; Guo, Q.; Zhou, Y.Y.; Yi, F.; Huang, G.M.; Li, Y.X.; Zhang, M.C.; Li, Z.H.; et al. Transcriptome dynamic landscape underlying the improvement of maize lodging resistance under coronatine treatment. BMC Plant Biol. 2021, 21, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Yu, J.P.F.A. Physiological effects of coronatine on maize seedlings under low temperature stress. J. Maize Sci. 2018, 26, 81–88. [Google Scholar]
- Wang, B.Q.; Li, Z.H.; Duan, L.S.; Zhai, Z.X. Effects of Coronatine on Photosynthesis Parameters and Endogenous Hormone Contents in Maize (Zea mays L.) Seedling under Drought Stress. Plant Physiol. Commun. 2008, 43, 269. [Google Scholar]
- Yan, C.; Fan, M.; Yang, M.; Zhao, J.P.; Zhang, W.H.; Su, Y.; Xiao, L.T.; Deng, H.T.; Xie, D.X. Injury activates Ca2þ/calmodulin-dependent phosphorylation of JAV1-JAZ8-WRKY51 complex for jasmonate biosynthesis. Mol. Cell 2018, 70, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Katsir, L.; Chung, H.S.; Koo, A.J.K.; Howe, G.A. Jasmonate signaling: A conserved mechanism of hormone sensing. Curr. Opin. Plant Biol. 2008, 11, 428–435. [Google Scholar] [CrossRef] [Green Version]
- He, R.; Su, H.; Wang, X.; Ren, Z.; Zhang, K.; Feng, T.; Zhang, M.; Li, Z.; Li, L.; Zhuang, J.; et al. Coronatine promotes maize water uptake by directly binding to the aquaporin ZmPIP2;5 and enhancing its activity. J. Integr. Plant Biol. 2022, 65, 611–875. [Google Scholar] [CrossRef]
- Baoqing, W.; Zhaohu, L.; Liusheng, D.; Zhixi, Z. Effect of Exogenous Coronatine on Contents of Endogenous Hormones in Maize Seedling under Drout Stress. Crops 2008, 126, 35–38. [Google Scholar]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Ghosh, S.; Chan, C.-K.K. Analysis of RNA-Seq data using TopHat and cufflinks. Plant Bioinform. 2016, 1374, 339–361. [Google Scholar]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi Chang, T. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Jiao, Y.; Peluso, P.; Shi, J.; Liang, T.; Stitzer, M.C.; Wang, B.; Campbell, M.S.; Stein, J.C.; Wei, X.; Chin, C.S.; et al. Improved maize reference genome with single-molecule technologies. Nature 2017, 546, 524–527. [Google Scholar] [CrossRef]
- Ku, Y.S.; Sintaha, M.; Cheung, M.Y.; Lam, H.M. Plant Hormone Signaling Crosstalks between Biotic and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [Green Version]
- Makowska, B.; Bakera, B.; Rakoczy-Trojanowska, M. the genetic background of benzoxazinoids. Acta Physiol. Plant. 2015, 37, 176. [Google Scholar] [CrossRef] [Green Version]
- Wouters, F.C.; Blanchette, B.; Gershenzon, J.; Vassao, D.G. Plant defense and herbivore counter defense: Benzoxazinoids and insect herbivores. Phytochem. Rev. 2016, 15, 1127–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.F.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.B.; Manzo, D.; Chervet, N.; Steinger, T.; van der Heijden, M.G.A.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, D.P.; Sellars, J.D.; Kenwright, A.M.; Steel, P.G. The maize benzoxazinone DIMBOA reacts with glutathione and other thiols to form spirocyclic adducts. Phytochemistry 2012, 77, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, F.; Maeli, M.; Yao, J.; He, S.Y. Jasmonate signaling and manipulation by pathogens and insects. J. Exp. Bot. 2017, 68, 1371–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling Network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [Green Version]
- Gen-Ichiro, A. Making Sense of the Way Plants Sense Herbivores. Trends Plant Sci. 2021, 26, 288–298. [Google Scholar]
- Zheng, X.Y.; Spivey, N.W.; Zeng, W.; Liu, P.P.; Fu, Z.Q.; Klessig, D.F.; He, S.Y.; Dong, X. Coronatine promotes Pseudomonas syringae virulence in plants by activating a signaling cascade that inhibits salicylic acid accumulation. Cell Host Microbe 2012, 11, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Zhang, L.; Zhou, C.; Yu, C.W.; Chaikam, V. HDA6 is required for jasmonate response, senescence and flowering in Arabidopsis. J. Exp. Bot. 2008, 59, 225–234. [Google Scholar] [CrossRef]
- Zhou, C.H.; Zhang, L.; Duan, J.; Miki, B.; Wu, K.Q. Histone deacetylase 19 is involved in jasmonic acid and ethylene signaling of pathogen response in Arabidopsis. Plant Cell 2005, 17, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.Q.; An, F.Y.; Feng, Y.; Li, P.P.; Xue, L.; Mu, A.; Jiang, Z.Q.; Kim, J.M.; To, T.K.; Li, W. Derepression of ethylene-stabilized transcription factors (EIN3/EIL1) mediates jasmonate and ethylene signaling synergy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 12539–12544. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, Q.; Zhai, H.; Gao, S.P.; Yang, L.; Wang, Z.; Xu, Y.T.; Huo, J.X.; Ren, Z.T.; Zhao, N.; et al. IbBBX24 Promotes the Jasmonic Acid Pathway and Enhances Fusarium Wilt Resistance in Sweet Potato. Plant Cell 2020, 32, 1102–1123. [Google Scholar] [CrossRef]
- Shyu, C.; Figueroa, P.; DePew, C.L.; Cooke, T.F.; Sheard, L.B.; Moreno, J.E.; Katsir, L.; Zheng, N.; Browse, J.; Howe, G.A. JAZ8 lacks a canonical degron and has an EAR motif that mediates transcriptional repression of jasmonate responses in Arabidopsis. Plant Cell 2012, 24, 536–550. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, A.; Ishihara, A.; Hasegawa, M.; Kodama, O.; Iwamura, H. Induced accumulation of 2-hydroxy-4,7-dimethoxy-1,4-benzoxazin-3-one glucoside (HDMBOA-Glc) in maize leaves. Phytochemistry 2001, 56, 669–675. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mapped Readings | Unique Mapped Readings | ||||
|---|---|---|---|---|---|
| Sample | Raw Readings | Number | % Mapped | Number | % Mapped |
| control1 | 46,020,064 | 37,748,120 | 82.03 | 25,691,854 | 55.83 |
| control2 | 46,188,984 | 37,766,492 | 81.77 | 25,679,736 | 55.68 |
| COR1 | 51,153,190 | 41,618,084 | 81.36 | 27,964,204 | 54.67 |
| COR2 | 45,283,680 | 36,566,492 | 80.75 | 24,400,408 | 53.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lou, Y.; Jin, X.; Jia, Z.; Sun, Y.; Xu, Y.; Liu, Z.; Tan, S.; Yi, F.; Duan, L. Coronatine-Based Gene Expression Changes Impart Partial Resistance to Fall Armyworm (Spodoptera frugiperda) in Seedling Maize. Genes 2023, 14, 735. https://doi.org/10.3390/genes14030735
Lou Y, Jin X, Jia Z, Sun Y, Xu Y, Liu Z, Tan S, Yi F, Duan L. Coronatine-Based Gene Expression Changes Impart Partial Resistance to Fall Armyworm (Spodoptera frugiperda) in Seedling Maize. Genes. 2023; 14(3):735. https://doi.org/10.3390/genes14030735
Chicago/Turabian StyleLou, Yuxuan, Xiaoxiao Jin, Zhiguo Jia, Yuqi Sun, Yiming Xu, Zihan Liu, Shuqian Tan, Fei Yi, and Liusheng Duan. 2023. "Coronatine-Based Gene Expression Changes Impart Partial Resistance to Fall Armyworm (Spodoptera frugiperda) in Seedling Maize" Genes 14, no. 3: 735. https://doi.org/10.3390/genes14030735