
Loss of Rose Fragrance under Chilling Stress Is Associated with Changes in DNA Methylation and Volatile Biosynthesis
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Chilling-Induced Influences on the Volatile Components of R. ‘Crimson Glory’
2.3. DNA Isolation, Library Construction, and WGBS-Seq
2.4. RNA-Seq and Differentially Expressed Gene (DEG) Identification
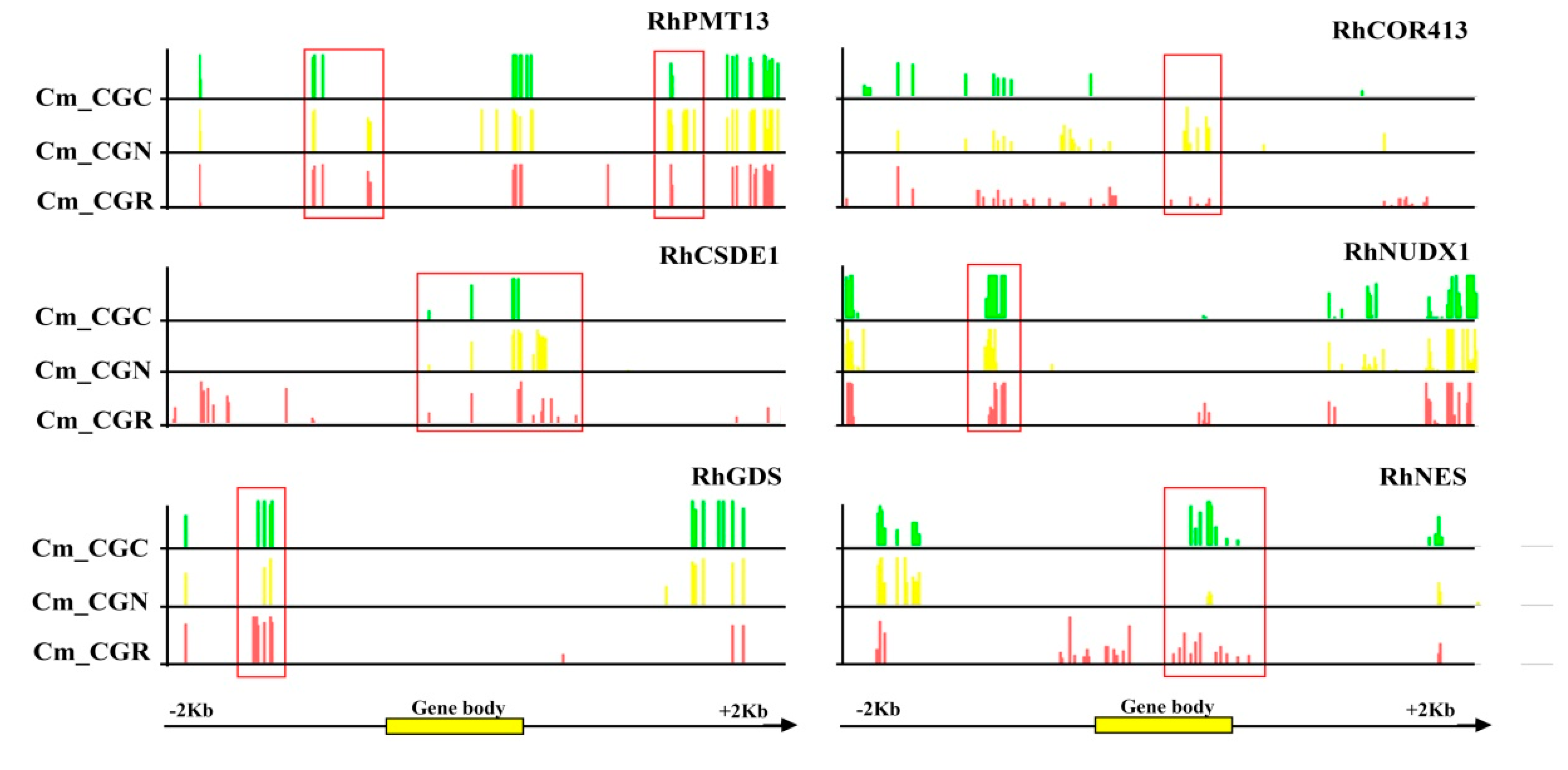
2.5. Identification of Differentially Methylated Regions (DMRs) and DMR-Related Genes
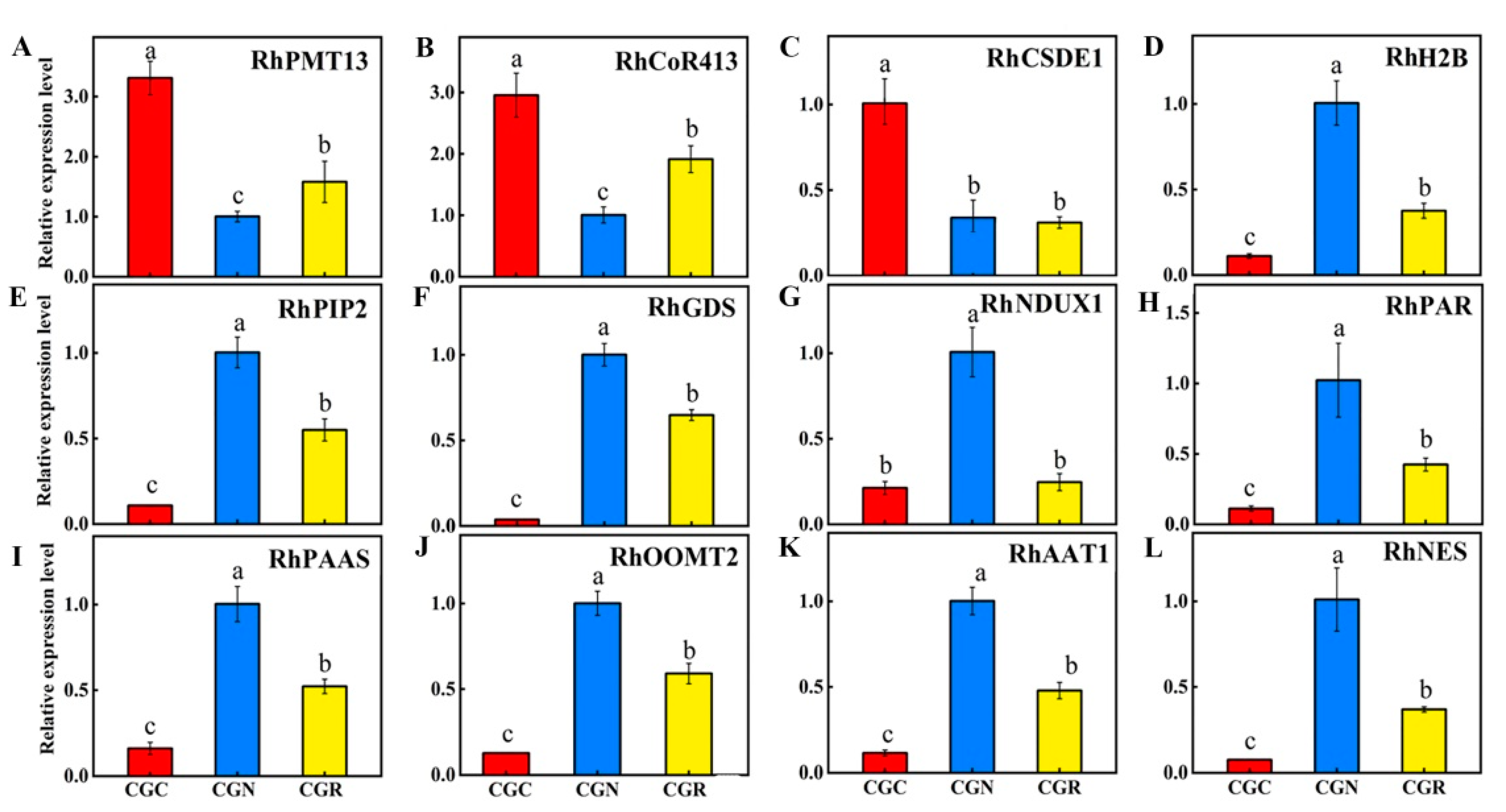
2.6. Gene Validation and Expression Analyses
3. Results
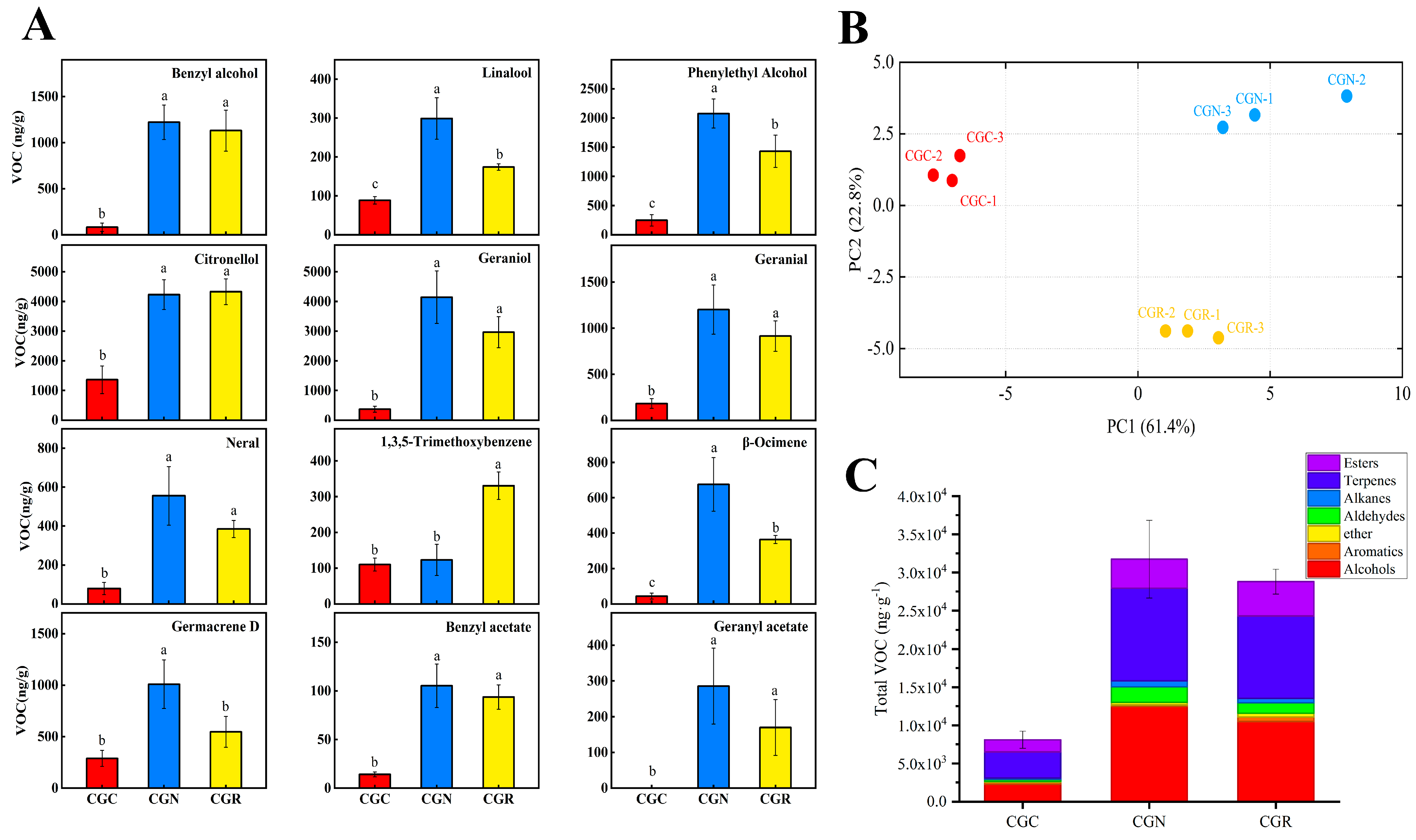
3.1. Chilling-Induced Influences on VOCs Emissions
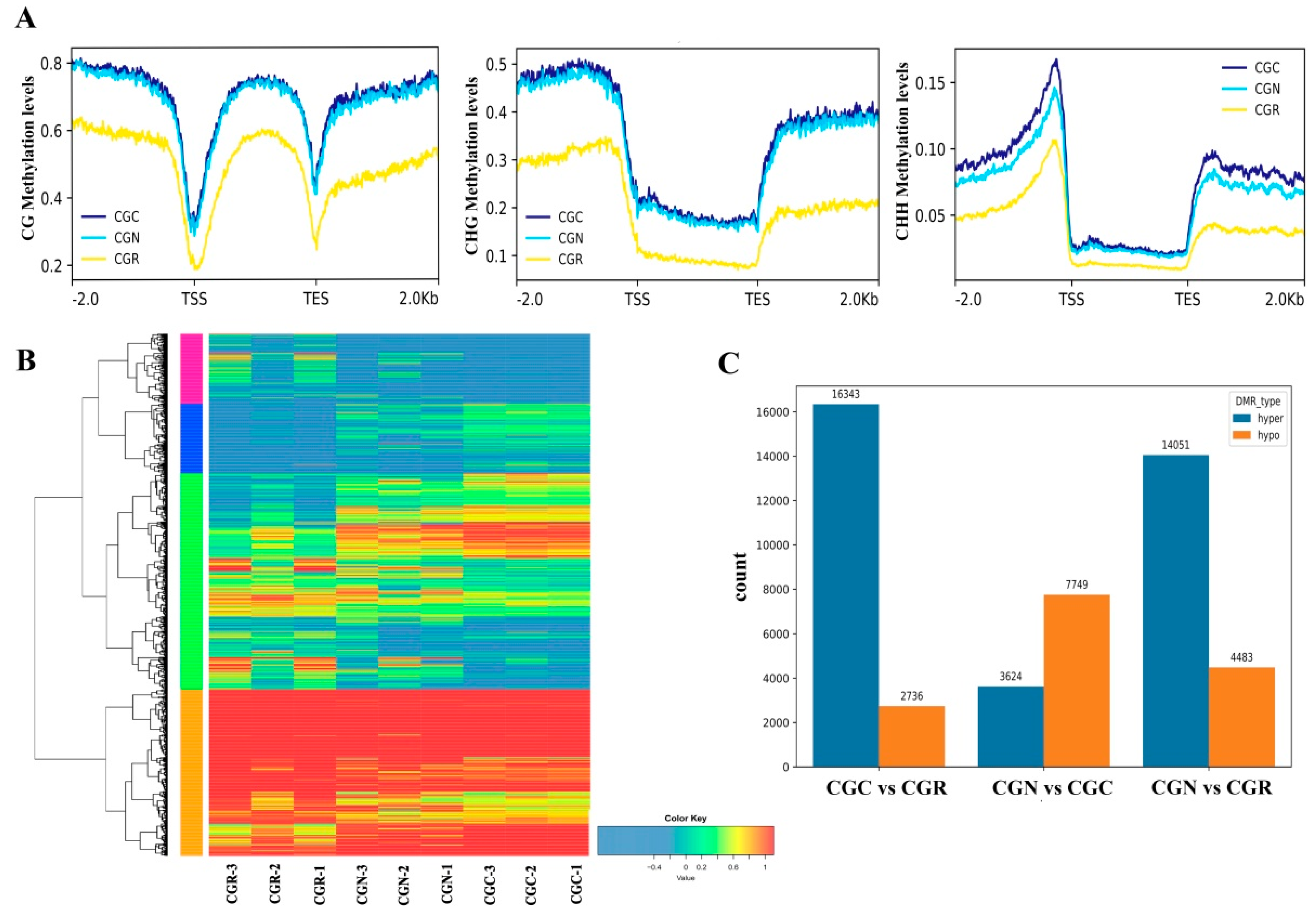
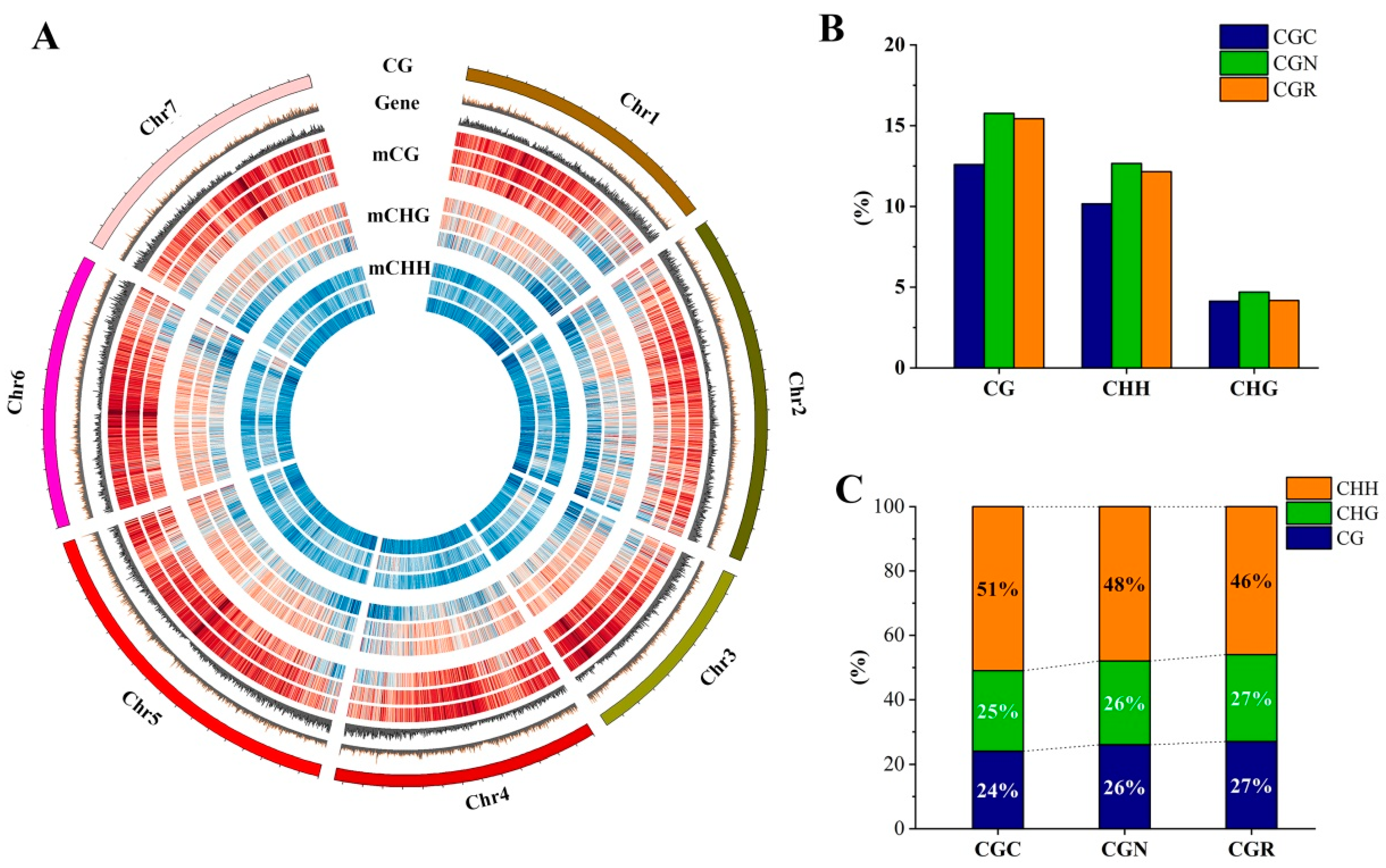
3.2. Characteristics of DNA Methylation in R. ‘Crimson Glory’
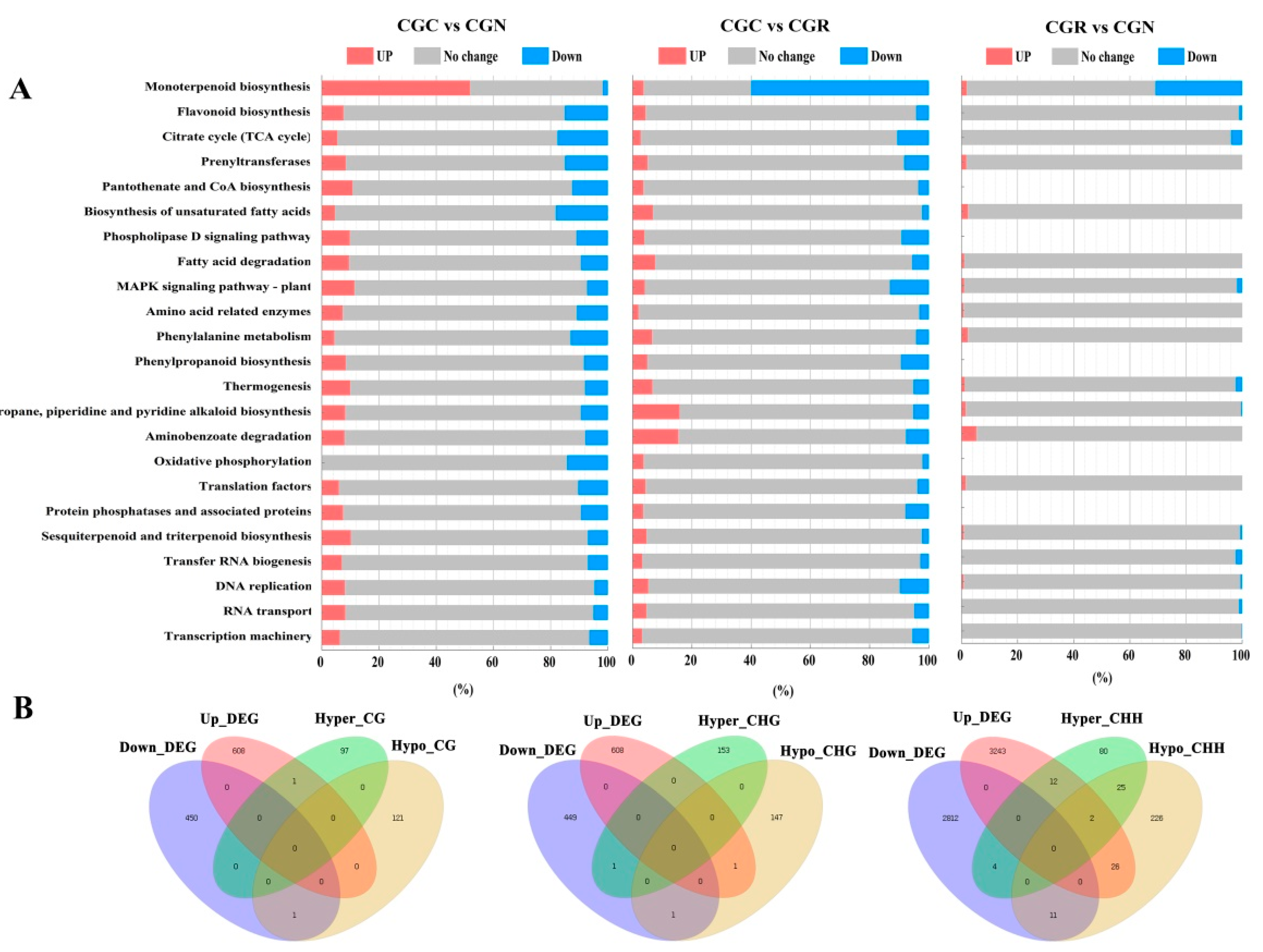
3.3. Identification of DNA Methylation and DEGs
3.4. Identification of DEGs and DMRs Associated with Volatile Biosynthesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shalit, M.; Guterman, I.; Volpin, H.; Bar, E.; Tamari, T.; Menda, N.; Adam, Z.; Zamir, D.; Vainstein, A.; Weiss, D.; et al. Volatile ester formation in roses. Identification of an Acetyl-Coenzyme A. Geraniol/Citronellol Acetyltransferase in Developing Rose Petals. Plant Physiol. 2003, 131, 1868–1876. [Google Scholar] [CrossRef] [Green Version]
- Roccia, A.; Hibrand-Saint, O.L.; Cavel, E.; Caissard, J.C.; Machenaud, J.; Thouroude, T.; Jeauffre, J.; Bony, A.; Dubois, A.; Vergne, P.; et al. Biosynthesis of 2-Phenylethanol in Rose Petals Is Linked to the Expression of One Allele of RhPAAS. Plant Physiol. 2019, 179, 1064–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.J.; Zhang, H.; Chen, M.; Jian, H.Y.; Baudino, S.; Caissard, J.C.; Bendahmane, M.; Li, S.B.; Zhang, T.; Zhou, N.N.; et al. Transcriptome and gene expression analysis during flower blooming in Rosa chinensis ‘Pallida’. Gene 2014, 540, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.H.; Du, X.Y.; Zhong, M.C.; Fang, W.; Suo, Z.Q.; Wang, D.; Dong, X.; Jiang, X.D.; Hu, J.Y. Complex and reticulate origin of edible roses (Rosa, Rosaceae) in China. Hortic. Res. 2022, 9, uhab051. [Google Scholar] [CrossRef]
- Ikeura, H.; Kobayashi, F.; Kai, T.; Tsuchiya, Y.; Tamaki, M. Effects of different storage conditions on the colour, antioxidant activity, and volatile components of edible roses. Sci. Hortic. 2023, 310, 111707. [Google Scholar] [CrossRef]
- Zhang, B.; Tieman, D.M.; Jiao, C.; Xu, Y.M.; Chen, K.S.; Fei, Z.J.; Giovannoni, J.J.; Klee, H.J. Chilling-induced tomato flavor loss is associated with altered volatile synthesis and transient changes in DNA methylation. Proc. Natl. Acad. Sci. USA 2016, 113, 12580–12585. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.Y. The Research of Chemical Components and Functions of Essential Oil of Rosa damascene; Zhejiang University: Hangzhou, China, 2012; pp. 4–10. [Google Scholar]
- Qiu, L.Q.; Zhang, M.; Mujumdar, A.S.; Chang, L. Convenient use of near-infrared spectroscopy to indirectly predict the antioxidant activitiy of edible rose (Rose chinensis Jacq “Crimsin Glory” H.T.) petals during infrared drying. Food Chem. 2022, 369, 130951. [Google Scholar] [CrossRef]
- Zhou, L.J.; Yu, C.; Cheng, B.X.; Wan, H.H.; Lou, L.; Pan, H.T.; Zhang, Q.X. Volatile compound analysis and aroma evaluation of tea-scented roses in China. Ind. Crops Prod. 2020, 155, 112735. [Google Scholar] [CrossRef]
- Shepherd, G.M. Smell images and the flavour system in the human brain. Nature 2006, 444, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Vivaldo, G.; Masi, E.; Taiti, C.; Caldarelli, G.; Mancuso, S. The network of plants volatile organic compounds. Sci. Rep. 2017, 7, 11050. [Google Scholar] [CrossRef] [Green Version]
- Sagae, M.; Oyama-Okubo, N.; Ando, T.; Marchesi, E.; Nakayama, M. Effect of temperature on the floral scent emission and endogenous volatile profile of Petunia axillaris. Biosci. Biotechnol. Biochem. 2008, 72, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Hansen, U.; Seufert, G. Temperature and light dependence of β-caryophyllene emission rates. J. Geophys. Res. Atmos. 2003, 108, 24. [Google Scholar] [CrossRef]
- Niinemets, U. Mild versus severe stress and BVOCs: Thresholds, priming and consequences. Trends Plant Sci. 2010, 15, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D.; Chen, X.; Yeh, S. Isoprene increases thermotolerance of fosmidomycin-fed leaves. Plant Physiol. 2001, 125, 2001–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, H.B.; Olsen, C.E. Influence of climatic factors on emission of flower volatiles in situ. Planta 1994, 192, 365–371. [Google Scholar] [CrossRef]
- Hu, Z.H.; Zhang, H.X.; Leng, P.S.; Zhao, J.; Wang, W.H.; Wang, S.D. The emission of floral scent from Lilium ‘siberia’ in response to light intensity and temperature. Acta Physiol. Plant. 2013, 35, 1691–1700. [Google Scholar] [CrossRef]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Niinemets, Ü.; Peñuelas, J. Changes in floral bouquets from compound-specific responses to increasing temperatures. Glob. Change Biol. 2014, 20, 3660–3669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picazo-Aragonés, J.; Terrab, A.; Balao, F. Plant volatile organic compounds evolution: Transcriptional regulation, epigenetics and polyploidy. Int. J. Mol. Sci. 2020, 21, 8956. [Google Scholar] [CrossRef] [PubMed]
- Gutschker, S.; Corral, J.M.; Schmiedl, A.; Ludewig, F.; Koch, W.; Fiedler-Wiechers, K.; Czarnecki, O.; Harms, K.; Keller, I.; Rodrigues, C.M.; et al. Multi-omics data integration reveals link between epigenetic modifications and gene expression in sugar beet (Beta vulgaris subsp. vulgaris) in response to cold. BMC Genomics 2022, 23, 144. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.; Li, R.P.; Huang, J.; Zhao, H.J.; Ge, R.H.; Wu, Q.; Mallano, A.I.; Wang, Y.L.; Li, F.D.; Deng, W.W.; et al. Divergent DNA methylation contributes to duplicated gene evolution and chilling response in tea plants. Plant J. 2021, 106, 1312–1327. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Chen, W.; Fan, T.G.; Tian, Y.R.; Zhang, S.; Zeng, D.X.; Li, Y.H. Low temperature-induced DNA hypermethylation attenuates expression of RhAG, an AGAMOUS homolog, and increases petal number in rose (Rosa hybrida). BMC Plant Biol. 2015, 15, 237. [Google Scholar] [CrossRef] [Green Version]
- Aslamarz, A.A.; Vahdati, K.; Rahemi, M.; Hasani, D. Cold-hardiness evaluation of Persian walnut by thermal analysis and freezing technique. Acta Hortic. 2010, 861, 269–272. [Google Scholar] [CrossRef]
- Gholizadeh, J.; Sadeghipour, H.; Abdolzadeh, A.; Hemmati, K.; Hassani, D.; Vahdati, K. Redox rather than carbohydrate metabolism differentiates endodormant lateral buds in walnut cultivars with contrasting chilling requirements. Sci. Hortic. 2017, 225, 29–37. [Google Scholar] [CrossRef]
- Newell-Price, J.; Clark, A.J.L.; King, P. DNA methylation and silencing of gene expression. Trends Endocrinol. Metab. 2000, 11, 142–148. [Google Scholar] [CrossRef]
- Urano, K.; Kurihara, Y.; Seki, M.; Shinozaki, K. ‘Omics’ analyses of regulatory networks in plant abiotic stress responses. Curr. Opin. Plant Biol. 2010, 13, 132–138. [Google Scholar] [CrossRef]
- Feng, D.D.; Wang, Q.G.; Jian, H.Y.; Zhang, H.; Qiu, X.Q.; Wang, Z.Z.; Du, W.W.; Xie, L.M.; Zhou, N.N.; Wang, H.C.; et al. Comparison of Volatile Compounds between Wild and Cultivated Roses. HortScience 2022, 57, 657–663. [Google Scholar] [CrossRef]
- Shi, S.; Zhang, S.; Wu, J.; Liu, X.; Zhang, Z. Identification of long non-coding RNAs involved in floral scent of Rosa hybrida. Front. Plant Sci. 2022, 13, 996474. [Google Scholar] [CrossRef]
- Raymond, O.; Gouzy, J.; Just, J.; Badouin, H.; Verdenaud, M.; Lemainque, A.; Vergne, P.; Moja, S.; Choisne, N.; Pont, C.; et al. The Rosa genome provides new insights into the domestication of modern roses. Nat. Genet. 2018, 50, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Akalin, A.; Kormaksson, M.; Li, S.; Garrett-Bakelman, F.E.; Figueroa, M.E.; Melnick, A.; Mason, C.E. methylKit: A comprehensive R package for the analysis of genome-wide DNA methylation profiles. Genome Biol. 2012, 3, R87. [Google Scholar] [CrossRef] [Green Version]
- Rajkumar, M.S.; Gupta, K.; Khemka, N.K.; Garg, R.; Jain, M. DNA ethylation reprogramming during seed development and its functional elevance in seed size/weight determination in chickpea. Commun. Biol. 2020, 3, 340. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Decato, B.; Hong, E.E.; Zhou, M.; Fang, F.; Qu, J.; Garvin, T.; Kessler, M.; Zhou, J.; Smith, A.D. A reference methylome database and analysis pipeline to facilitate integrative and comparative epigenomics. PLoS ONE 2013, 8, e81148–e81156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Appl. Biosyst. 2001, 25, 402–408. [Google Scholar]
- Kim, J.M.; Sasaki, T.; Ueda, M.; Sako, K.; Seki, M. Chromatin changes in response to drought, salinity, heat, and cold stresses in plants. Front. Plant Sci. 2015, 6, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbanczyk-Wochniak, E.; Usadel, B.; Thimm, O.; Nunes-Nesi, A.; Carrari, F.; Davy, M.; Bläsing, O.; Kowalczyk, M.; Weicht, D.; Polinceusz, A.; et al. Conversion of MapMan to allow the analysis of transcript data from Solanaceous species: Effects of genetic and environmental alterations in energy metabolism in the leaf. Plant Mol. Biol. 2006, 60, 773–792. [Google Scholar] [CrossRef] [PubMed]
- Viggiano, L.; DePinto, M.C. Dynamic DNA methylation patterns in stress response. In Plant Epigenetics. RNA Technologies; Rajewsky, N., Jurga, S., Barciszewski, J., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 281–302. [Google Scholar]
- Hu, X.F.; Liu, J.H.; Liu, E.H.; Qiao, K.; Gong, S.F.; Wang, J.G.; Zhou, A.M.; Zhang, J.Z. Arabidopsis cold-regulated plasma membrane protein Cor413pm1 is a regulator of ABA response. Biochem. Biophys. Res. Commun. 2021, 561, 88–92. [Google Scholar] [CrossRef]
- Dubois, A.; Carrere, S.; Raymond, O.; Pouvreau, B.; Cottret, L.; Roccia, A.; Onesto, J.P.; Sakr, S.; Atanassova, R.; Baudino, S.; et al. Transcriptome database resource and gene expression atlas for the rose. BMC Genomics 2012, 13, 638. [Google Scholar] [CrossRef] [Green Version]
- Magnard, J.L.; Roccia, A.; Caissard, J.C.; Vergne, P.; Sun, P.; Hecquet, R.; Dubois, A.; Hibrand-Saint, O.L.; Jullien, F.; Nicolè, F.; et al. Biosynthesis of monoterpene scent compounds in roses. Science 2015, 349, 81–83. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.M.; Kobayashi, H.; Sakai, M.; Hirata, H.; Asai, T.; Ohnishi, T.; Baldermann, S.; Watanabe, N. Functional characterization of rose phenylacetaldehyde reductase (PAR), an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol. J. Plant Physiol. 2011, 168, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Magnard, J.L.; Bony, A.R.; Bettini, F.; Campanaro, A.; Blerot, B.; Baudino, S.; Jullien, F. Linalool and linalool nerolidol synthases in roses, several genes for little scent. Plant Physiol. Biochem. 2018, 127, 74–87. [Google Scholar] [CrossRef]
- Scalliet, G.; Piola, F.; Douady, C.J.; Réty, S.; Raymond, O.; Baudino, S.; Bordji, K.; Bendahmane, M.; Dumas, C.; Cock, J.M.; et al. Scent evolution in Chinese roses. Proc. Natl. Acad. Sci. USA 2008, 105, 5927–5932. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Llusià, J. The complexity of factors driving volatile organic compound emissions by plants. Biol. Plantarum 2001, 44, 481–487. [Google Scholar] [CrossRef]
- Grote, R.; Monson, R.K.; Niinemets, Ü. Leaf-level models of constitutive and stress-driven volatile organic compound emissions. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 315–355. [Google Scholar]
- Fu, J.X.; Hou, D.; Zhang, C.; Bao, Z.Y.; Zhao, H.B.; Hu, S.Q. The Emission of the Floral Scent of Four Osmanthus fragrans Cultivars in Response to Different Temperatures. Molecules 2017, 22, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renard, C.M.; Ginies, C.; Gouble, B.; Bureau, S.; Causse, M. Home conservation strategies for tomato (Solanum lycopersicum): Storage temperature vs. duration-Is there a compromise for better aroma preservation? Food Chem. 2013, 139, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Pichersky, E. Biochemical and Molecular Genetic Aspects of Floral Scents. Plant Physiol. 2000, 122, 627–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, L.Z.; Xu, W.; Ma, P.F.; Guo, Z.H.; Li, D.Z. Single-base methylome analysis reveals dynamic changes of genome-wide DNA methylation associated with rapid stem growth of woody bamboos. Planta 2022, 256, 53. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, X.; Guo, Y.; Zheng, Y.; Yue, C.; Yang, J.; Ye, N. Identification of CBF transcription factors in tea plants and a survey of potential CBF target genes under low temperature. Int. J. Mol. Sci. 2019, 20, 5137. [Google Scholar] [CrossRef] [Green Version]
- Shan, X.H.; Wang, X.Y.; Yang, G.; Wu, Y.; Su, S.Z.; Li, S.P.; Liu, H.K.; Yuan, Y.P. Analysis of the DNA methylation of maize (Zea mays L.) in response to cold stress based on methylation-sensitive amplified polymorphisms. J. Plant Biol. 2013, 56, 32–38. [Google Scholar] [CrossRef]
- Zhou, A.M.; Sun, H.W.; Feng, S.; Zhou, M.; Gong, S.F.; Wang, J.G.; Zhang, S.Z. A novel cold-regulated gene from Phlox subulata, PsCor413im1, enhances low temperature tolerance in Arabidopsis. Biochem. Biophys. 2018, 495, 1688–1694. [Google Scholar] [CrossRef]
- Huang, D.Q.; Wu, W.R.; Abrams, S.R.; Cutler, A.J. The relationship of drought-related gene expression in Arabidopsis thaliana to hormonal and environmental factors. J. Exp. Bot. 2008, 59, 2991–3007. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.W.; Zhang, X.H.; Gao, J.F.; Wang, P.K.; Xu, X.Y.; Liu, Z.X.; Shen, S.H.; Feng, B.L. A buckwheat (Fagopyrum esculentum) DRE-binding transcription factor gene, FeDREB1, enhances freezing and drought tolerance of transgenic Arabidopsis. Plant Mol. Biol. 2015, 33, 1510–1525. [Google Scholar] [CrossRef]
- Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M.; Maeshima, M. Identification of 33 Rice Aquaporin genes and analysis of their expression and function. Plant Cell Physiol. 2005, 46, 1568–1577. [Google Scholar] [CrossRef] [Green Version]
- Aroca, R.; Amodeo, G.; Fernández-Illescas, S.; Herman, E.M.; Chaumont, F.; Chrispeels, M.J. The role of aquaporins and membrane damage in chilling and hydrogen peroxide induced changes in the hydraulic conductance of maize roots. Plant Physiol. 2005, 137, 341–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.Y.; Kim, D.G.; Kim, Y.O.; Kim, J.S.; Kang, H. An expression analysis of a gene family encoding plasma membrane aquaporins in response to abiotic stresses in Arabidopsis thaliana. Plant Mol. Biol. 2004, 54, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Katsuhara, M.; Chung, G.C.; Sakurai, J. Low temperature and aquaporins, a molecular mechanism of water transport. Cryobiol. Cryotechnol. 2007, 53, 21–32. [Google Scholar]
- Guo, H.; Wu, T.K.; Li, S.X.; He, Q.H.; Yang, Z.L.; Zhang, W.H.; Gan, Y.; Sun, P.Y.; Xiang, G.L.; Zhang, H.Y.; et al. The methylation patterns and transcriptional responses to chilling stress at the seedling stage in rice. Int. J. Mol. Sci. 2019, 20, 5089. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.X.; Li, Z.W.; Chen, X.; Yun, Z.; Li, T.T.; Jiang, Y.M. Comparative metabolites profiling of harvested papaya (Carica paoaya L.) peel in response to chilling stress. J. Sci. Food Agric. 2019, 99, 6868–6881. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, L.; Bai, X.; Zhang, H.; Qiu, X.; Jian, H.; Wang, Q.; Wang, H.; Feng, D.; Tang, K.; Yan, H. Loss of Rose Fragrance under Chilling Stress Is Associated with Changes in DNA Methylation and Volatile Biosynthesis. Genes 2023, 14, 692. https://doi.org/10.3390/genes14030692
Xie L, Bai X, Zhang H, Qiu X, Jian H, Wang Q, Wang H, Feng D, Tang K, Yan H. Loss of Rose Fragrance under Chilling Stress Is Associated with Changes in DNA Methylation and Volatile Biosynthesis. Genes. 2023; 14(3):692. https://doi.org/10.3390/genes14030692
Chicago/Turabian StyleXie, Limei, Xue Bai, Hao Zhang, Xianqin Qiu, Hongying Jian, Qigang Wang, Huichun Wang, Dedang Feng, Kaixue Tang, and Huijun Yan. 2023. "Loss of Rose Fragrance under Chilling Stress Is Associated with Changes in DNA Methylation and Volatile Biosynthesis" Genes 14, no. 3: 692. https://doi.org/10.3390/genes14030692