
Deep Learning-Based Feature Extraction with MRI Data in Neuroimaging Genetics for Alzheimer’s Disease
 ,
on behalf of the Alzheimer’s Disease Neuroimaging Initiative
,
on behalf of the Alzheimer’s Disease Neuroimaging Initiative
Abstract
:1. Introduction
2. Data
Preprocessing and Description
3. Methods
3.1. Classification and Feature Extraction
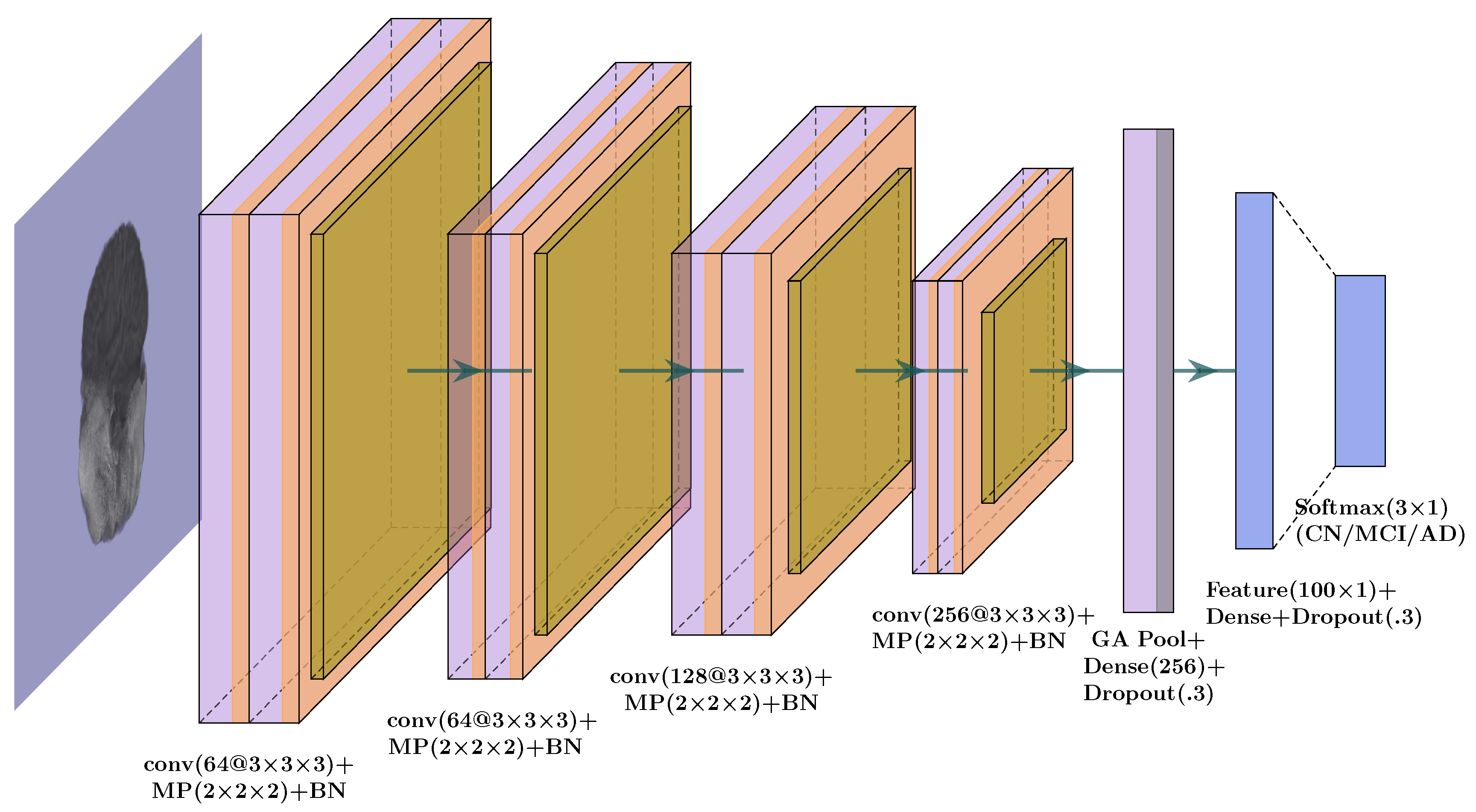
3.1.1. Whole-Brain Structure
3.1.2. Multi-Branch CNN
3.2. Segmentation
3.3. GWAS
3.4. PC vs. Gray Matter Volume of Regions of Interests (ROIs)
3.5. Identifying Genetic Pathways
3.6. Evaluation Criteria
4. Results
4.1. Whole-Brain Structure
4.2. Multi-Branch CNN
4.3. PC vs. Gray Matter Volumes of ROIs
4.4. Genetic Pathway Identification with Enrichment Analysis
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kramarow, E.A.; Tejada-Vera, B. Dementia Mortality in the United States, 2000–2017; Centers for Disease Control and Prevention National Center for Health Statistics National Vital Statistics System: Atlanta, GA, USA, 2019; Volume 68, pp. 1–29. [Google Scholar]
- Alzheimer’s Association. Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2018, 14, 367–429. [Google Scholar] [CrossRef]
- Petersen, R.C.; Aisen, P.S.; Beckett, L.A.; Donohue, M.C.; Gamst, A.C.; Harvey, D.J.; Jack, C.R.; Jagust, W.J.; Shaw, L.M.; Toga, A.W.; et al. Alzheimer’s Disease Neuroimaging Initiative (ADNI): Clinical characterization. Neurology 2010, 74, 201–209. [Google Scholar] [CrossRef]
- Elliott, L.T.; Sharp, K.; Smith, S.M. Genome-wide association studies of brain imaging phenotypes in UK Biobank. Nature 2018, 562, 210–216. [Google Scholar] [CrossRef]
- Gillies, R.J.; Kinahan, P.E.; Hricak, H. Radiomics: Images Are More than Pictures, They Are Data. Radiology 2016, 278, 563–577. [Google Scholar] [CrossRef]
- Pan, Y.; Mai, Q.; Zhang, X. Covariate-Adjusted Tensor Classification in High Dimensions. J. Am. Stat. Assoc. 2019, 114, 1305–1319. [Google Scholar] [CrossRef]
- Miranda, M.F.; Zhu, H.; Ibrahim, J.G. TPRM: Tensor partition regression models with applications in imaging biomarker detection. Ann. Appl. Stat. 2018, 12, 1422–1450. [Google Scholar] [CrossRef]
- Shi, R.; Kang, J. Thresholded Multiscale Gaussian Processes with Application to Bayesian Feature Selection for Massive Neuroimaging Data. arXiv 2015, arXiv:1504.06074. [Google Scholar]
- Feng, X.; Li, T.; Song, X.; Zhu, H. Bayesian Scalar on Image Regression With Nonignorable Nonresponse. J. Am. Stat. Assoc. 2020, 115, 1574–1597. [Google Scholar] [CrossRef]
- Saykin, A.J.; Shen, L.; Foroud, T.M.; Potkin, S.G.; Swaminathan, S.; Kim, S.; Risacher, S.L.; Nho, K.; Huentelman, M.J.; Craig, D.W.; et al. Alzheimer’s Disease Neuroimaging Initiative biomarkers as quantitative phenotypes: Genetics core aims, progress, and plans. Alzheimers Dement. 2010, 6, 265–273. [Google Scholar] [CrossRef]
- Zhao, B.; Li, T.; Yang, Y.; Wang, X.; Luo, T.; Shan, Y.; Zhu, Z.; Xiong, D.; Hauberg, M.E.; Bendl, J.; et al. Common genetic variation influencing human white matter microstructure. Science 2021, 372, eabf3736. [Google Scholar] [CrossRef]
- Pan, D.; Zeng, A.; Jia, L.; Huang, Y.; Frizzell, T.; Song, X. Early Detection of Alzheimer’s Disease Using Magnetic Resonance Imaging: A Novel Approach Combining Convolutional Neural Networks and Ensemble Learning. Front. Neurosci. 2020, 14, 259. [Google Scholar] [CrossRef]
- Dhinagar, N.J.; Thomopoulos, S.I.; Rajagopalan, P.; Stripelis, D.; Ambite, J.L.; Steeg, G.V.; Thompson, P.M. Evaluation of Transfer Learning Methods for Detecting Alzheimer’s Disease with Brain MRI. bioRxiv 2022. [Google Scholar] [CrossRef]
- Lama, R.K.; Gwak, J.; Park, J.S.; Lee, S.W. Diagnosis of Alzheimer’s Disease Based on Structural MRI Images Using a Regularized Extreme Learning Machine and PCA Features. J. Healthc. Eng. 2017, 2017, 5485080. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, X.; Kim, M.; Bao, J.; Shen, L. A Novel Bayesian Semi-parametric Model for Learning Heritable Imaging Traits. In Proceedings of the Medical Image Computing and Computer Assisted Intervention (MICCAI 2021), Virtual, 27 September–1 October 2021; Springer International Publishing: Berlin/Heidelberg, Germany, 2021; pp. 678–687. [Google Scholar] [CrossRef]
- Bao, J.; Wen, Z.; Kim, M.; Zhao, X.; Lee, B.N.; Jung, S.H.; Davatzikos, C.; Saykin, A.J.; Thompson, P.M.; Kim, D.; et al. Identifying highly heritable brain amyloid phenotypes through mining Alzheimer’s imaging and sequencing biobank data. Pac. Symp. Biocomput. 2022, 27, 109–120. [Google Scholar] [CrossRef]
- Grasby, K.L.; Jahanshad, N.; Painter, J.N.; Colodro-Conde, L.; Bralten, J.; Hibar, D.P.; Lind, P.A.; Pizzagalli, F.; Ching, C.R.K.; McMahon, M.A.B.; et al. The genetic architecture of the human cerebral cortex. Science 2020, 367, eaay6690. [Google Scholar] [CrossRef]
- Zhao, B.; Li, T.; Smith, S.M.; Xiong, D.; Wang, X.; Yang, Y.; Luo, T.; Zhu, Z.; Shan, Y.; Matoba, N.; et al. Common variants contribute to intrinsic human brain functional networks. Nat. Genet. 2022, 54, 508–517. [Google Scholar] [CrossRef]
- Zhao, B.; Luo, T.; Li, T.; Li, Y.; Zhang, J.; Shan, Y.; Wang, X.; Yang, L.; Zhou, F.; Zhu, Z.; et al. Genome-wide association analysis of 19,629 individuals identifies variants influencing regional brain volumes and refines their genetic co-architecture with cognitive and mental health traits. Nat. Genet. 2019, 51, 1637–1644. [Google Scholar] [CrossRef]
- Potkin, S.G.; Guffanti, G.; Lakatos, A.; Turner, J.A.; Kruggel, F.; Fallon, J.H.; Saykin, A.J.; Orro, A.; Lupoli, S.; Salvi, E.; et al. Hippocampal Atrophy as a Quantitative Trait in a Genome-Wide Association Study Identifying Novel Susceptibility Genes for Alzheimer’s Disease. PLoS ONE 2009, 4, e6501. [Google Scholar] [CrossRef]
- Shen, L.; Thompson, P.M.; Potkin, S.G.; Bertram, L.; Farrer, L.A.; Foroud, T.M.; Green, R.C.; Hu, X.; Huentelman, M.J.; Kim, S.; et al. Genetic analysis of quantitative phenotypes in AD and MCI: Imaging, cognition and biomarkers. Brain Imaging Behav. 2013, 8, 183–207. [Google Scholar] [CrossRef]
- Tang, X.; Bi, X.; Qu, A. Individualized Multilayer Tensor Learning With an Application in Imaging Analysis. J. Am. Stat. Assoc. 2020, 115, 836–851. [Google Scholar] [CrossRef]
- Wang, J.X.; Li, Y.; Li, X.; Lu, Z.H. Alzheimer’s Disease Classification Through Imaging Genetic Data with IGnet. Front. Neurosci. 2022, 16, 846638. [Google Scholar] [CrossRef]
- Smith, S.M. Fast robust automated brain extraction. Hum. Brain Mapp. 2002, 17, 143–155. [Google Scholar] [CrossRef]
- Woolrich, M.W.; Jbabdi, S.; Patenaude, B.; Chappell, M.; Makni, S.; Behrens, T.; Beckmann, C.; Jenkinson, M.; Smith, S.M. Bayesian analysis of neuroimaging data in FSL. NeuroImage 2009, 45, S173–S186. [Google Scholar] [CrossRef]
- Chakraborty, D.; Roy, A. Time Series Methodology in STORJ Token Prediction. In Proceedings of the 2019 International Conference on Data Mining Workshops (ICDMW), Beijing, China, 8–11 November 2019. [Google Scholar] [CrossRef]
- Kruthika, K.; Rajeswari; Maheshappa, H. CBIR system using Capsule Networks and 3D CNN for Alzheimer’s disease diagnosis. Inform. Med. Unlocked 2019, 14, 59–68. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, R.; Yeh, M.; Qu, A. Dermoscopic Image Classification with Neural Style Transfer. J. Comput. Graph. Stat. 2022, 31, 1318–1331. [Google Scholar] [CrossRef]
- Gao, X.W.; Hui, R.; Tian, Z. Classification of CT brain images based on deep learning networks. Comput. Methods Programs Biomed. 2017, 138, 49–56. [Google Scholar] [CrossRef]
- Lu, B.; Li, H.X.; Chang, Z.K.; Li, L.; Chen, N.X.; Zhu, Z.C.; Zhou, H.X.; Li, X.Y.; Wang, Y.W.; Cui, S.X.; et al. A Practical Alzheimer Disease Classifier via Brain Imaging-Based Deep Learning on 85,721 Samples. J. Big Data 2020, 9, 101. [Google Scholar] [CrossRef]
- Fukushima, K. Neocognitron: A self-organizing neural network model for a mechanism of pattern recognition unaffected by shift in position. Biol. Cybern. 1980, 36, 193–202. [Google Scholar] [CrossRef]
- Nwankpa, C.; Ijomah, W.; Gachagan, A.; Marshall, S. Activation Functions: Comparison of trends in Practice and Research for Deep Learning. arXiv 2018, arXiv:1811.03378. [Google Scholar]
- Lee, W.Y.; Park, S.M.; Sim, K.B. Optimal hyperparameter tuning of convolutional neural networks based on the parameter-setting-free harmony search algorithm. Optik 2018, 172, 359–367. [Google Scholar] [CrossRef]
- Krizhevsky, A.; Sutskever, I.; Hinton, G.E. ImageNet Classification with Deep Convolutional Neural Networks. In Proceedings of the 25th International Conference on Neural Information Processing Systems—NIPS’12, Lake Tahoe, NV, USA, 3–6 December 2012; Curran Associates Inc.: Red Hook, NY, USA, 2012; Volume 1, pp. 1097–1105. [Google Scholar]
- Simonyan, K.; Zisserman, A. Very Deep Convolutional Networks for Large-Scale Image Recognition. arXiv 2015, arXiv:1409.1556. [Google Scholar]
- He, K.; Zhang, X.; Ren, S.; Sun, J. Deep Residual Learning for Image Recognition. In Proceedings of the 2016 IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Las Vegas, NV, USA, 27–30 June 2016; pp. 770–778. [Google Scholar] [CrossRef]
- Zhang, Y.; Brady, M.; Smith, S. Segmentation of brain MR images through a hidden Markov random field model and the expectation-maximization algorithm. IEEE Trans. Med Imaging 2001, 20, 45–57. [Google Scholar] [CrossRef]
- Ribeiro, M.T.; Singh, S.; Guestrin, C. “Why Should I Trust You?”: Explaining the Predictions of Any Classifier. In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, KDD ’16, San Francisco, CA, USA, 13–17 August 2016; Association for Computing Machinery: New York, NY, USA, 2016; pp. 1135–1144. [Google Scholar] [CrossRef]
- Lindberg, O.; Walterfang, M.; Looi, J.C.; Malykhin, N.; Östberg, P.; Zandbelt, B.; Martin, S.; Beatriz, P.; Dennis, V.; Eva, Ö.; et al. Hippocampal Shape Analysis in Alzheimer’s Disease and Frontotemporal Lobar Degeneration Subtypes. J. Alzheimer’s Dis. 2012, 30, 355–365. [Google Scholar] [CrossRef]
- Wang, J.; Syal, C. Biomarker-guided drug therapy: Personalized medicine for treating Alzheimer’s disease. Neural Regen. Res. 2021, 16, 2010. [Google Scholar] [CrossRef]
- Yan, T.; Ding, F.; Zhao, Y. Integrated identification of key genes and pathways in Alzheimer’s disease via comprehensive bioinformatical analyses. Hereditas 2019, 156, 25. [Google Scholar] [CrossRef]
- Watanabe, K.; Taskesen, E.; van Bochoven, A.; Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef]
- Bradley, A.P. The use of the area under the ROC curve in the evaluation of machine learning algorithms. Pattern Recognit. 1997, 30, 1145–1159. [Google Scholar] [CrossRef]
- Denny, J.C.; Bastarache, L.; Ritchie, M.D.; Carroll, R.J.; Zink, R.; Mosley, J.D.; Field, J.R.; Pulley, J.M.; Ramirez, A.H.; Bowton, E.; et al. Systematic comparison of phenome-wide association study of electronic medical record data and genome-wide association study data. Nat. Biotechnol. 2013, 31, 1102–1111. [Google Scholar] [CrossRef]
- Hoogenraad, N.J.; Ward, L.A.; Ryan, M.T. Import and assembly of proteins into mitochondria of mammalian cells. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2002, 1592, 97–105. [Google Scholar] [CrossRef]
- Hu, R.; Yu, Q.; Zhou, S.; Yin, Y.; Hu, R.; Lu, H.; Hu, B. Co-expression Network Analysis Reveals Novel Genes Underlying Alzheimer’s Disease Pathogenesis. Front. Aging Neurosci. 2020, 12, 605961. [Google Scholar] [CrossRef]
- Rouillard, A.D.; Gundersen, G.W.; Fernandez, N.F.; Wang, Z.; Monteiro, C.D.; McDermott, M.G.; Ma’ayan, A. The harmonizome: A collection of processed datasets gathered to serve and mine knowledge about genes and proteins. Database 2016, 2016, baw100. [Google Scholar] [CrossRef]
- Shen, L.; Kim, S.; Risacher, S.L.; Nho, K.; Swaminathan, S.; West, J.D.; Foroud, T.; Pankratz, N.; Moore, J.H.; Sloan, C.D.; et al. Whole genome association study of brain-wide imaging phenotypes for identifying quantitative trait loci in MCI and AD: A study of the ADNI cohort. NeuroImage 2010, 53, 1051–1063. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Z.; Shen, X.; Pan, W. Testing for association with multiple traits in generalized estimation equations, with application to neuroimaging data. NeuroImage 2014, 96, 309–325. [Google Scholar] [CrossRef]
- Nazarian, A.; Yashin, A.I.; Kulminski, A.M. Genome-wide analysis of genetic predisposition to Alzheimer’s disease and related sex disparities. Alzheimer’s Res. Ther. 2019, 11, 5. [Google Scholar] [CrossRef]
- Furney, S.J.; Simmons, A.; Breen, G.; Pedroso, I.; Lunnon, K.; Proitsi, P.; Hodges, A.; Powell, J.; Wahlund, L.O.; Kloszewska, I.; et al. Genome-wide association with MRI atrophy measures as a quantitative trait locus for Alzheimer’s disease. Mol. Psychiatry 2010, 16, 1130–1138. [Google Scholar] [CrossRef]
- Boison, D. Adenosine Kinase: Exploitation for Therapeutic Gain. Pharmacol. Rev. 2013, 65, 906–943. [Google Scholar] [CrossRef]
- Sandau, U.S.; Diogenes, M.C.O.M.J.; Sebastiao, A.M.; Boison, D. Adenosine Kinase Deficiency in the Brain Results in Maladaptive Synaptic Plasticity. J. Neurosci. 2016, 36, 12117–12128. [Google Scholar] [CrossRef]
- Sieg, K. Neurodevelopmental Disorders Associated with Chromosome 15. Jefferson J. Psychiatry 1990, 8. [Google Scholar] [CrossRef]
- Kuang, X.L.; Zhao, X.M.; Xu, H.F.; Shi, Y.Y.; Deng, J.B.; Sun, G.T. Spatio-temporal expression of a novel neuron-derived neurotrophic factor (NDNF) in mouse brains during development. BMC Neurosci. 2010, 11, 137. [Google Scholar] [CrossRef]
- Sze, C.I. Role of WWOX WOX1 in Alzheimer s disease pathology and in cell death signaling. Front. Biosci. 2012, 4, 1951–1965. [Google Scholar] [CrossRef]
- Setti, S.E.; Hunsberger, H.C.; Reed, M.N. Alterations in hippocampal activity and Alzheimer’s disease. Transl. Issues Psychol. Sci. 2017, 3, 348–356. [Google Scholar] [CrossRef]
- Jacobs, H.I.L.; Hopkins, D.A.; Mayrhofer, H.C.; Bruner, E.; van Leeuwen, F.W.; Raaijmakers, W.; Schmahmann, J.D. The cerebellum in Alzheimer’s disease: Evaluating its role in cognitive decline. Brain 2017, 141, 37–47. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, Y.; Liu, S.; Song, X.; Yang, X.Z.; Fan, Y.; Chen, W.; Akdemir, Z.C.; Yan, Z.; Zou, Y.; et al. The coexistence of copy number variations (CNVs) and single nucleotide polymorphisms (SNPs) at a locus can result in distorted calculations of the significance in associating SNPs to disease. Hum. Genet. 2018, 137, 553–567. [Google Scholar] [CrossRef]
- Wang, H.; Yang, J.; Schneider, J.A.; Jager, P.L.D.; Bennett, D.A.; Zhang, H.Y. Genome-wide interaction analysis of pathological hallmarks in Alzheimer’s disease. Neurobiol. Aging 2020, 93, 61–68. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Test AUC (CI) | PC | SNP (N/K) | Chr | p-Value |
|---|---|---|---|---|---|
| ine Augmentation with respect to whole brain | 0.75 (0.717, 0.782) | 3 | (K) [46] | 1 | 2.06 |
| 2 | (K) ([44]) | 19 | 4.58 | ||
| 4 | (N) | 3 | |||
| ine AD vs. CN binary classification | 0.90 (0.885, 0.914) | 4 | (K) [47] | 8 | 4.21 |
| 7 | (N) | 15 | 4.33 |
| Model | Test AUC (CI) | PC | SNP (N/K) | Chr | p-Value |
|---|---|---|---|---|---|
| ine Multi-branch | 0.76 (0.729, 0.790) | 4 | (K) [55] | 8 | 9.40 |
| 7 | (K) [56] | 16 | 2.67 | ||
| 9 | (N) | 7 | 9.59 | ||
| ine Multi-branch on GM images | 0.74 (0.707, 0.772) | 2 | (N) | 6 | |
| 3 | (N) | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakraborty, D.; Zhuang, Z.; Xue, H.; Fiecas, M.B.; Shen, X.; Pan, W., on behalf of the Alzheimer’s Disease Neuroimaging Initiative. Deep Learning-Based Feature Extraction with MRI Data in Neuroimaging Genetics for Alzheimer’s Disease. Genes 2023, 14, 626. https://doi.org/10.3390/genes14030626
Chakraborty D, Zhuang Z, Xue H, Fiecas MB, Shen X, Pan W on behalf of the Alzheimer’s Disease Neuroimaging Initiative. Deep Learning-Based Feature Extraction with MRI Data in Neuroimaging Genetics for Alzheimer’s Disease. Genes. 2023; 14(3):626. https://doi.org/10.3390/genes14030626
Chicago/Turabian StyleChakraborty, Dipnil, Zhong Zhuang, Haoran Xue, Mark B. Fiecas, Xiatong Shen, and Wei Pan on behalf of the Alzheimer’s Disease Neuroimaging Initiative. 2023. "Deep Learning-Based Feature Extraction with MRI Data in Neuroimaging Genetics for Alzheimer’s Disease" Genes 14, no. 3: 626. https://doi.org/10.3390/genes14030626