
miR-33a Inhibits the Differentiation of Bovine Preadipocytes through the IRS2–Akt Pathway

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Ethics Statement
2.2. Sample Preparation, Cell Culture, and Transfection
2.3. RNA Preparation and Quantitative Real-Time PCR
2.4. Oil Red O Staining and BODIPY Staining
2.5. Western Blotting
2.6. Vector Construction
2.7. Luciferase Activity Assay
2.8. Statistical Analysis
3. Results
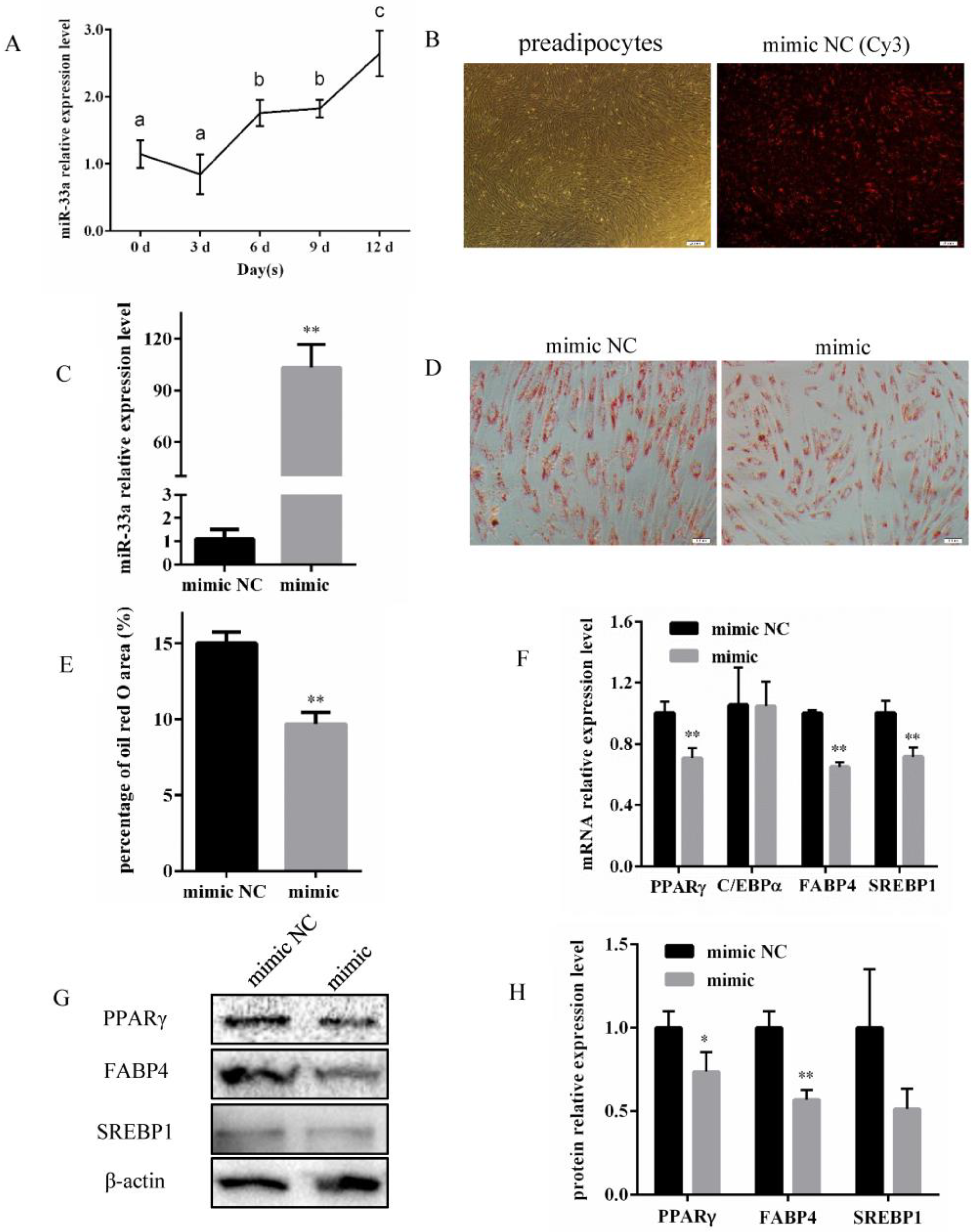
3.1. miR-33a Overexpression Inhibits Differentiation of Bovine Preadipocytes
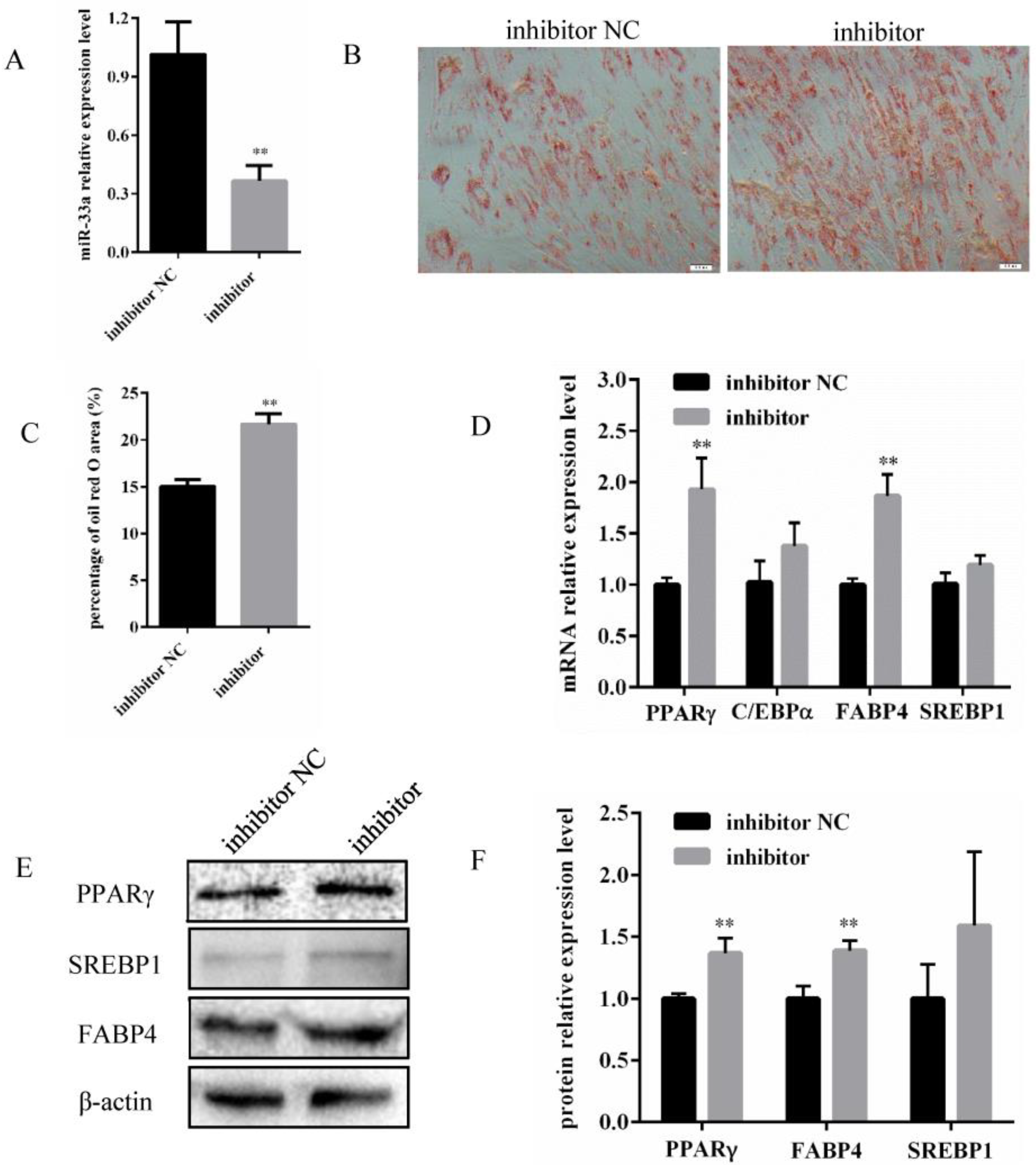
3.2. Inhibition of miR-33a Expression Promotes Differentiation of Bovine Preadipocytes
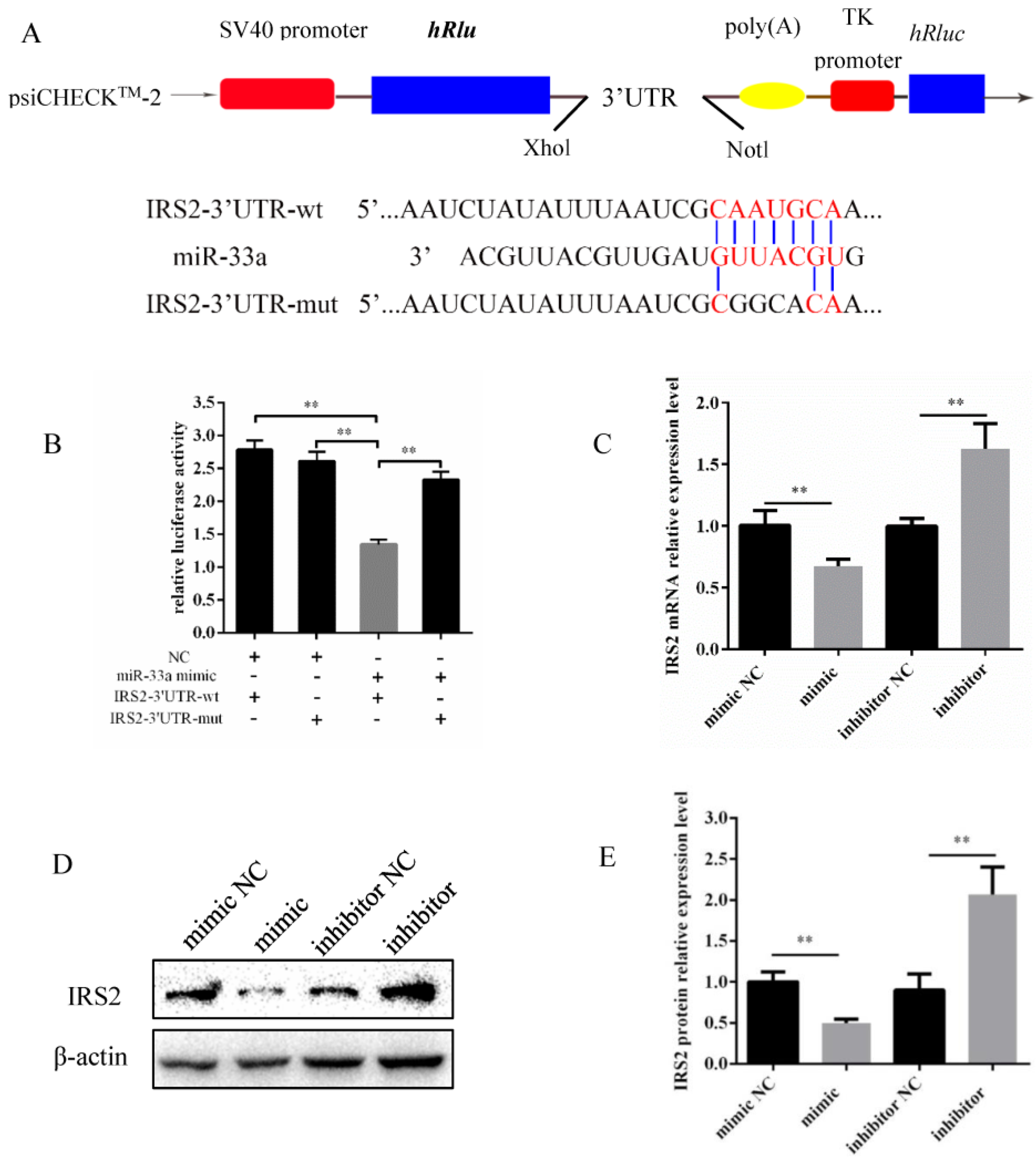
3.3. miR-33a Inhibits IRS2 Expression by Directly Targeting 3′-UTR
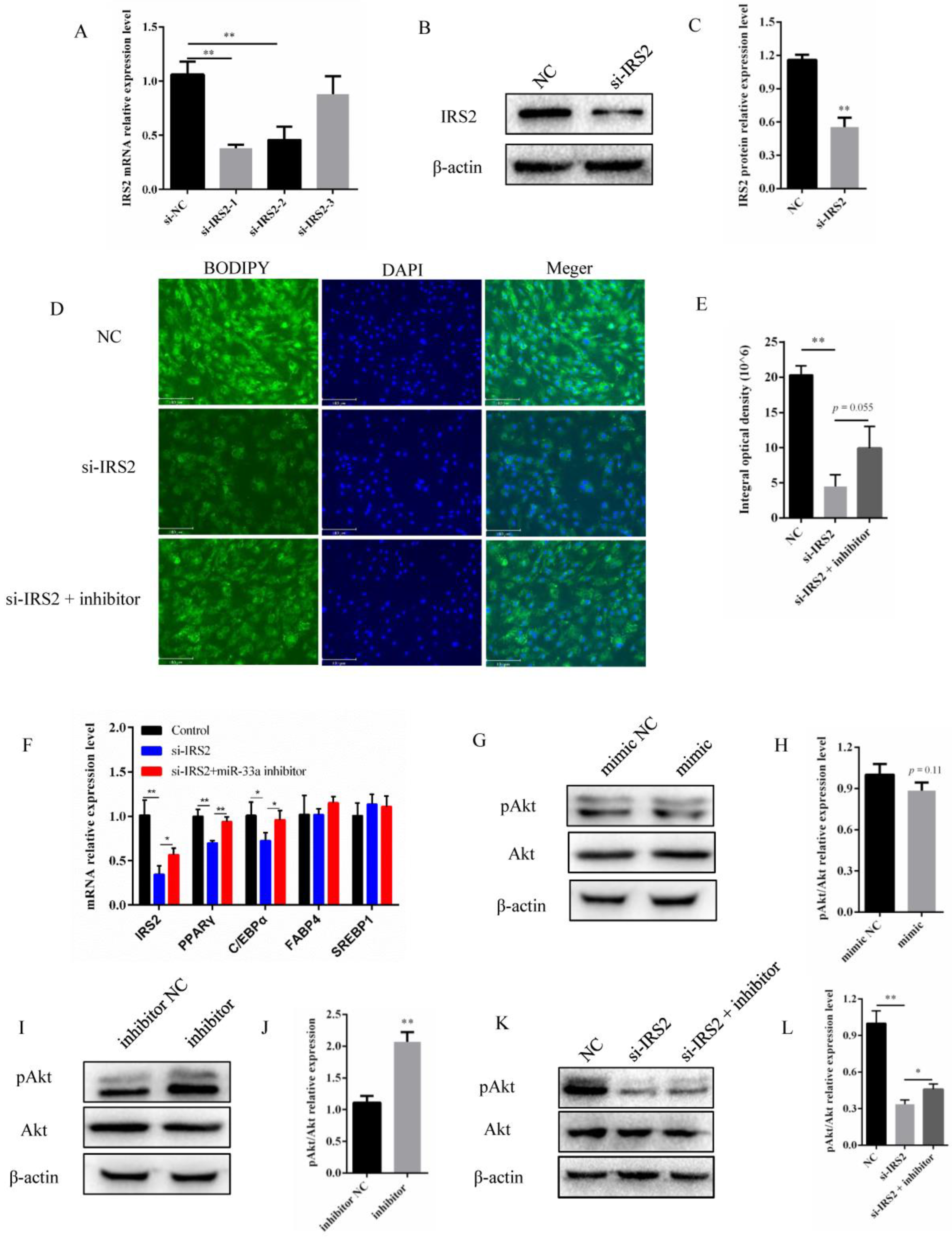
3.4. miR-33a Regulated the Activity of the IRS2–Akt Pathway in Preadipocyte Differentiation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, Y.H.; Mottillo, E.P.; Granneman, J.G. Adipose tissue plasticity from WAT to BAT and in between. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 358–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Gurmaches, J.; Guertin, D.A. Adipocyte lineages: Tracing back the origins of fat. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 340–351. [Google Scholar] [CrossRef] [Green Version]
- Harvey, I.; Boudreau, A.; Stephens, J.M. Adipose tissue in health and disease. Open Biol. 2020, 10, 200291. [Google Scholar] [CrossRef]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Obesity and cancer risk: Emerging biological mechanisms and perspectives. Metab. Clin. Exp. 2019, 92, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and nonalcoholic fatty liver disease: From pathophysiology to therapeutics. Metab. Clin. Exp. 2019, 92, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Ortega, F.B.; Lavie, C.J.; Blair, S.N. Obesity and Cardiovascular Disease. Circ. Res. 2016, 118, 1752–1770. [Google Scholar] [CrossRef] [Green Version]
- Bhupathiraju, S.N.; Hu, F.B. Epidemiology of Obesity and Diabetes and Their Cardiovascular Complications. Circ. Res. 2016, 118, 1723–1735. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.X.; Xie, Y.H.; Chen, W.; Zhang, Y.; Zeng, Y.Q. The role of long noncoding RNAs in livestock adipose tissue deposition—A review. Anim. Biosci. 2021, 34, 1089–1099. [Google Scholar] [CrossRef]
- Gesta, S.; Tseng, Y.H.; Kahn, C.R. Developmental origin of fat: Tracking obesity to its source. Cell 2007, 131, 242–256. [Google Scholar] [CrossRef] [Green Version]
- Hausman, D.B.; DiGirolamo, M.; Bartness, T.J.; Hausman, G.J.; Martin, R.J. The biology of white adipocyte proliferation. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2001, 2, 239–254. [Google Scholar] [CrossRef]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPAR γ is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Lecoutre, S.; Deracinois, B.; Laborie, C.; Eberle, D.; Guinez, C.; Panchenko, P.E.; Lesage, J.; Vieau, D.; Junien, C.; Gabory, A.; et al. Depot- and sex-specific effects of maternal obesity in offspring’s adipose tissue. J. Endocrinol. 2016, 230, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, Y.; Lu, S.; Yin, L.; Zong, C.; Cui, S.; Qin, D.; Yang, Y.; Guan, Q.; Li, X.; et al. The role and possible mechanism of lncRNA U90926 in modulating 3T3-L1 preadipocyte differentiation. Int. J. Obes. 2017, 41, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Engin, A.B. MicroRNA and Adipogenesis. Adv. Exp. Med. Biol. 2017, 960, 489–509. [Google Scholar] [PubMed]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esau, C.; Kang, X.L.; Peralta, E.; Hanson, E.; Marcusson, E.G.; Ravichandran, L.V.; Sun, Y.Q.; Koo, S.; Perera, R.J.; Jain, R.; et al. MicroRNA-143 regulates adipocyte differentiation. J. Biol. Chem. 2004, 279, 52361–52365. [Google Scholar] [CrossRef] [Green Version]
- Kajimoto, K.; Naraba, H.; Iwai, N. MicroRNA and 3T3-L1 pre-adipocyte differentiation. RNA 2006, 12, 1626–1632. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, Y.C.; Wang, J.H.; Kong, J.; Qi, Y.C.; Quigg, R.J.; Li, X.M. miR-17-92 cluster accelerates adipocyte differentiation by negatively regulating. tumor-suppressor Rb2/p130. Proc. Natl. Acad. Sci. USA 2008, 105, 2889–2894. [Google Scholar] [CrossRef] [Green Version]
- Arner, P.; Kulyte, A. MicroRNA regulatory networks in human adipose tissue and obesity. Nat. Rev. Endocrinol. 2015, 11, 276–288. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, S.; Zhang, W.Z.; Cheng, G.; Khan, R.; Junjvlieke, Z.; Li, S.J.; Zan, L.S. miR-424 Promotes Bovine Adipogenesis Through an Unconventional Post-Transcriptional Regulation of STK11. Front. Genet. 2020, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, S.; Cheng, G.; Mei, C.G.; Li, S.J.; Zhang, W.Z.; Junjvlieke, Z.; Zan, L.S. MiR-145 reduces the activity of PI3K/Akt and MAPK signaling pathways and inhibits adipogenesis in bovine preadipocytes. Genomics 2020, 112, 2688–2694. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Wang, H.B.; Wang, Y.N.; Wang, H.C.; Zhang, S.; Hong, J.Y.; Guo, H.F.; Chen, D.; Yang, Y.; Zan, L.S. Transcriptome analysis of mRNA and microRNAs in intramuscular fat tissues of castrated and intact male Chinese Qinchuan cattle. PLoS ONE 2017, 12, e0185961. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Z.; Wang, L.; Raza, S.H.A.; Wang, X.Y.; Wang, G.H.; Liang, C.C.; Cheng, G.; Li, B.Z.; Zan, L.S. MiR-33a plays a crucial role in the proliferation of bovine preadipocytes. Adipocyte 2021, 10, 189–200. [Google Scholar] [CrossRef]
- Keppler-Noreuil, K.M.; Parker, V.E.R.; Darling, T.N.; Martinez-Agosto, J.A. Somatic overgrowth disorders of the PI3K/AKT/mTOR pathway & therapeutic strategies. Am. J. Med. Genet. C 2016, 172, 402–421. [Google Scholar]
- Li, T.; Zhang, L.; Jin, C.; Xiong, Y.; Cheng, Y.Y.; Chen, K. Pomegranate flower extract bidirectionally regulates the proliferation, differentiation and apoptosis of 3T3-L1 cells through regulation of PPARγ expression mediated by PI3K-AKT signaling pathway. Biomed. Pharmacother. 2020, 131, 110769. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Dong, J.C.; Bian, Q. Small molecules for mesenchymal stem cell fate determination. World J. Stem Cells 2019, 11, 1084–1103. [Google Scholar] [CrossRef] [PubMed]
- White, M.F. Regulating insulin signaling and β-cell function through IRS proteins. Can. J. Physiol. Pharmacol. 2006, 84, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Wei, Q.; Zhai, X.C.; Qin, L.; Li, H.B.; Meng, R.; Chen, S.C. MiRNA-7b-5p attenuates the progression of osteoporosis by inhibiting adipose differentiation of hMSCs via regulating IRS2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9207–9214. [Google Scholar]
- Groeneveld, M.P.; Brierley, G.V.; Rocha, N.M.; Siddle, K.; Semple, R.K. Acute knockdown of the insulin receptor or its substrates Irs1 and 2 in 3T3-L1 adipocytes suppresses adiponectin production. Sci. Rep. 2016, 6, 21105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Ning, Y.; Li, P.; Guo, H.; Zan, L. Tissue Expression Analysis and Characterization of Smad3 Promoter in Bovine Myoblasts and Preadipocytes. DNA Cell Biol. 2018, 37, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.Z.; Sun, B.; Zhao, Y.Q.; Raza, S.H.A.; Li, Y.S.; Wang, J.F.; Ma, X.H.; Almohaimeed, H.M.; Shaheen, S.; Al-Sarraj, F.; et al. Proliferation of bovine myoblast by LncPRRX1 via regulation of the miR-137/CDC42 axis. Int. J. Biol. Macromol. 2022, 220, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.N.; Yang, W.C.; Li, P.W.; Wang, H.B.; Zhang, Y.Y.; Zan, L.S. Myocyte enhancer factor 2A promotes proliferation and its inhibition attenuates myogenic differentiation via myozenin 2 in bovine skeletal muscle myoblast. PLoS ONE 2018, 13, e0196255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercado-Matos, J.; Clark, J.L.; Piper, A.J.; Janusis, J.; Shaw, L.M. Differential involvement of the microtubule cytoskeleton in insulin receptor substrate 1 (IRS-1) and IRS-2 signaling to AKT determines the response to microtubule disruption in breast carcinoma cells. J. Biol. Chem. 2017, 292, 7806–7816. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.H.; Lim, D.S. Insulin receptor substrate 2: A bridge between Hippo and AKT pathways. BMB Rep. 2018, 51, 209–210. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.J.; Han, F.; Cui, Q.Y.; Liao, W.F.; Liu, H.; Guan, G.P.; Yang, L. IRS2 depletion inhibits cell proliferation and decreases hormone secretion in mouse granulosa cells. J. Reprod. Dev. 2018, 64, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Price, N.L.; Fernandez-Hernando, C. miRNA regulation of white and brown adipose tissue differentiation and function. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids. 2016, 1861, 2104–2110. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Pan, Y.; Zhao, B.; Qiao, L.; Liu, J.; Liang, Y.; Liu, W. MiR-33a inhibits the adipogenic differentiation of ovine adipose-derived stromal vascular fraction cells by targeting SIRT6. Domest. Anim. Endocrinol. 2021, 74, 106513. [Google Scholar] [CrossRef]
- Wu, Y.N.; Zhang, W.; Wang, Y.H.; Lu, Q.Y.; Zhou, J.P.; Chen, Z.; Yang, Z.P.; Loor, J.J. Molecular mechanisms in the miR-33a/LPPR4 pathway regulating unsaturated fatty acid synthesis in bovine mammary epithelial cells. Anim. Biotechnol. 2022, 1–13. [Google Scholar] [CrossRef]
- Fan, G.; Xu, Z.; Hu, X.; Li, M.; Zhou, J.; Zeng, Y.; Xie, Y. miR-33a hinders the differentiation of adipose mesenchymal stem cells towards urothelial cells in an inductive condition by targeting β-catenin and TGFR. Mol. Med. Rep. 2018, 17, 2341–2348. [Google Scholar] [CrossRef] [Green Version]
- Hanouskova, B.; Neprasova, B.; Skalova, L.; Maletinska, L.; Zemanova, K.; Ambroz, M.; Matouskova, P. High-fructose drinks affect microRNAs expression differently in lean and obese mice. J. Nutr. Biochem. 2019, 68, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Baik, M.; Kang, H.J.; Park, S.J.; Na, S.W.; Piao, M.; Kim, S.Y.; Fassah, D.M.; Moon, Y.S. Triennial growth and development symposium: Molecular mechanisms related to bovine intramuscular fat deposition in the longissimus muscle. J. Anim. Sci. 2017, 95, 2284–2303. [Google Scholar] [CrossRef] [PubMed]
- Salone, V.; Rederstorff, M. Stem-loop RT-PCR based quantification of small non-coding RNAs. Methods Mol. Biol. 2015, 1296, 103–108. [Google Scholar] [PubMed]
- Tang, C.Y.; Man, X.F.; Guo, Y.; Tang, H.N.; Tang, J.; Zhou, C.L.; Tan, S.W.; Wang, M.; Zhou, H.D. IRS-2 Partially Compensates for the Insulin Signal Defects in IRS-1(-/-) Mice Mediated by miR-33. Mol. Cells 2017, 40, 123–132. [Google Scholar] [CrossRef]
- Chen, Z.; Shi, H.P.; Sun, S.; Xu, H.F.; Cao, D.Y.; Luo, J. MicroRNA-181b suppresses TAG via target IRS2 and regulating multiple genes in the Hippo pathway. Exp. Cell Res. 2016, 348, 66–74. [Google Scholar] [CrossRef]
- Prockop, D.J. Marrow stromal cells as steam cells for nonhematopoietic tissues. Science 1997, 276, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.L.; Yang, L.; Liu, X.F.; Hong, T.; Wang, T.; Dong, A.W.; Li, J.X.; Xu, X.Y.; Cao, L.L. miR-431 inhibits adipogenic differentiation of human bone marrow-derived mesenchymal stem cells via targeting insulin receptor substance 2. Stem Cell Res. Ther. 2018, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Wirth, D.; Schmitz, I.; Meyer-Hermann, M. A mathematical model of the impact of insulin secretion dynamics on selective hepatic insulin resistance. Nat. Commun. 2017, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Calejman, C.M.; Trefely, S.; Entwisle, S.W.; Luciano, A.; Jung, S.M.; Hsiao, W.; Torres, A.; Hung, C.M.; Li, H.; Snyder, N.W.; et al. mTORC2-AKT signaling to ATP-citrate lyase drives brown adipogenesis and de novo lipogenesis. Nat. Commun. 2020, 11, 16. [Google Scholar] [CrossRef]
- Vissenberg, R.; Manders, V.D.; Mastenbroek, S.; Fliers, E.; Afink, G.B.; Ris-Stalpers, C.; Goddijn, M.; Bisschop, P.H. Pathophysiological aspects of thyroid hormone disorders/thyroid peroxidase autoantibodies and reproduction. Hum. Reprod. Update 2015, 21, 378–387. [Google Scholar] [CrossRef] [Green Version]
- Burgos-Ramos, E.; Chowen, J.A.; Arilla-Ferreiro, E.; Canelles, S.; Argente, J.; Barrios, V. Chronic central leptin infusion modifies the response to acute central insulin injection by reducing the interaction of the insulin receptor with IRS2 and increasing its association with SOCS3. J. Neurochem. 2011, 117, 175–185. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Raza, S.H.A.; Li, B.; Sun, B.; Wang, S.; Pant, S.D.; Al-Abbas, N.S.; Shaer, N.A.; Zan, L. miR-33a Inhibits the Differentiation of Bovine Preadipocytes through the IRS2–Akt Pathway. Genes 2023, 14, 529. https://doi.org/10.3390/genes14020529
Zhang W, Raza SHA, Li B, Sun B, Wang S, Pant SD, Al-Abbas NS, Shaer NA, Zan L. miR-33a Inhibits the Differentiation of Bovine Preadipocytes through the IRS2–Akt Pathway. Genes. 2023; 14(2):529. https://doi.org/10.3390/genes14020529
Chicago/Turabian StyleZhang, Wenzhen, Sayed Haidar Abbas Raza, Bingzhi Li, Bing Sun, Sihu Wang, Sameer D. Pant, Nouf S. Al-Abbas, Nehad A. Shaer, and Linsen Zan. 2023. "miR-33a Inhibits the Differentiation of Bovine Preadipocytes through the IRS2–Akt Pathway" Genes 14, no. 2: 529. https://doi.org/10.3390/genes14020529