
CeRNA Network Reveals the Circular RNA Characterization in Goat Ear Fibroblasts Reprogramming into Mammary Epithelial Cells
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of circRNA
2.2. Prediction of circRNA Targeting Relationship
2.3. Differential Expression Analysis of circRNA
2.4. Functional Enrichment and Analysis of Differentially Expressed circRNA-Derived Genes Construction of PPI Network
3. Results
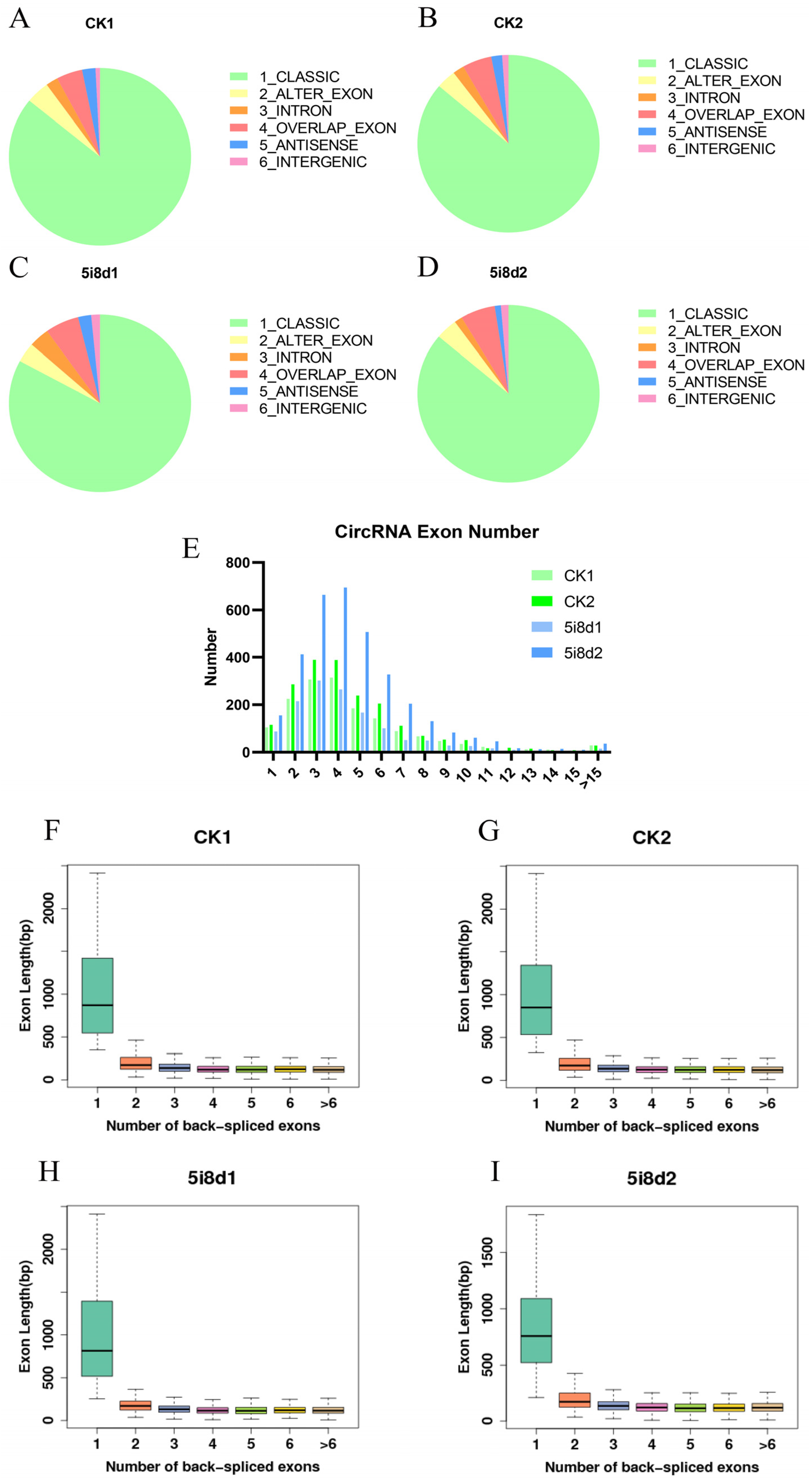
3.1. Sequencing and Characterization of circRNAs
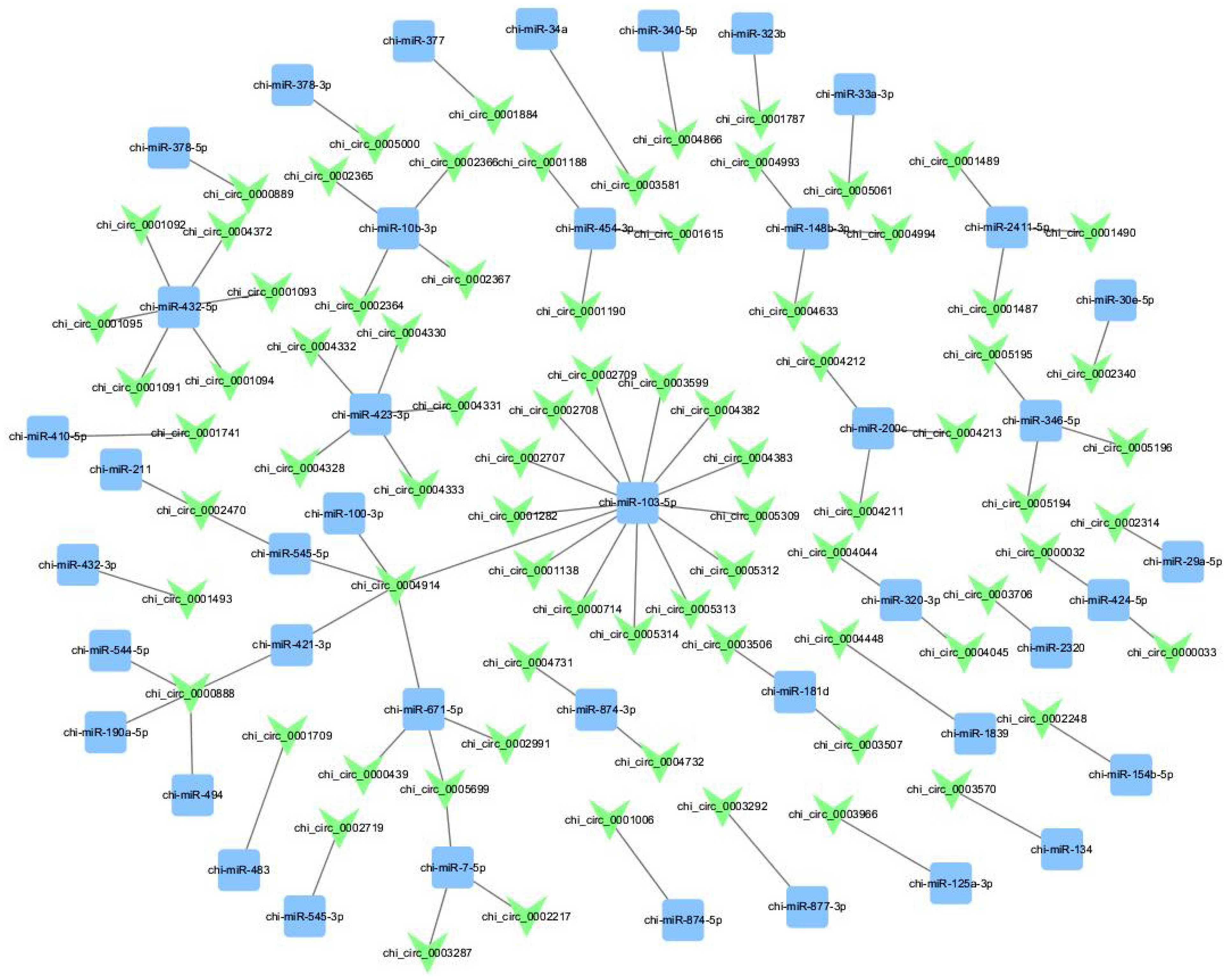
3.2. The Potential Functions of the Detected circRNA’s Sponge with miRNA
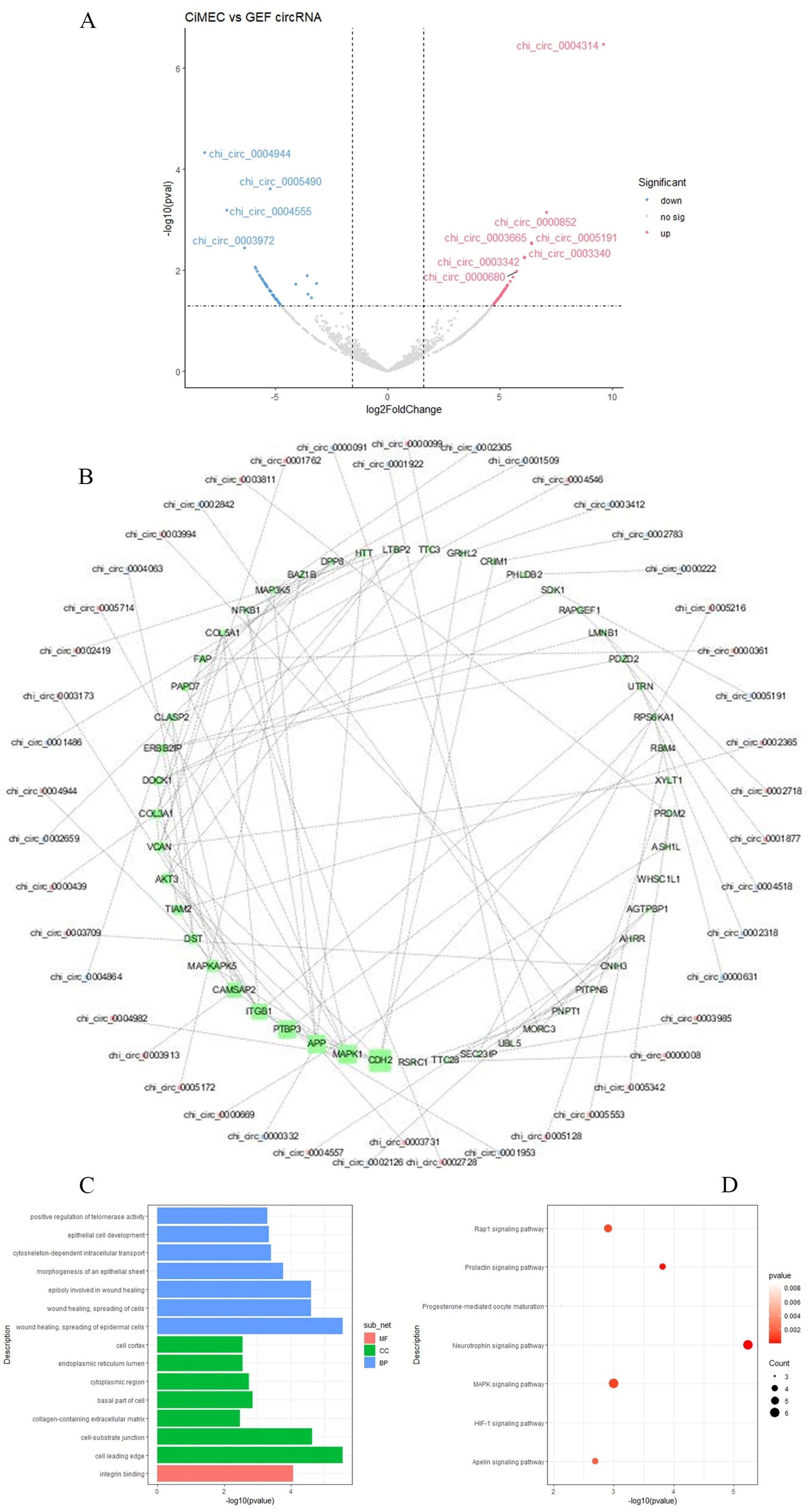
3.3. Differentially Expressed circRNA
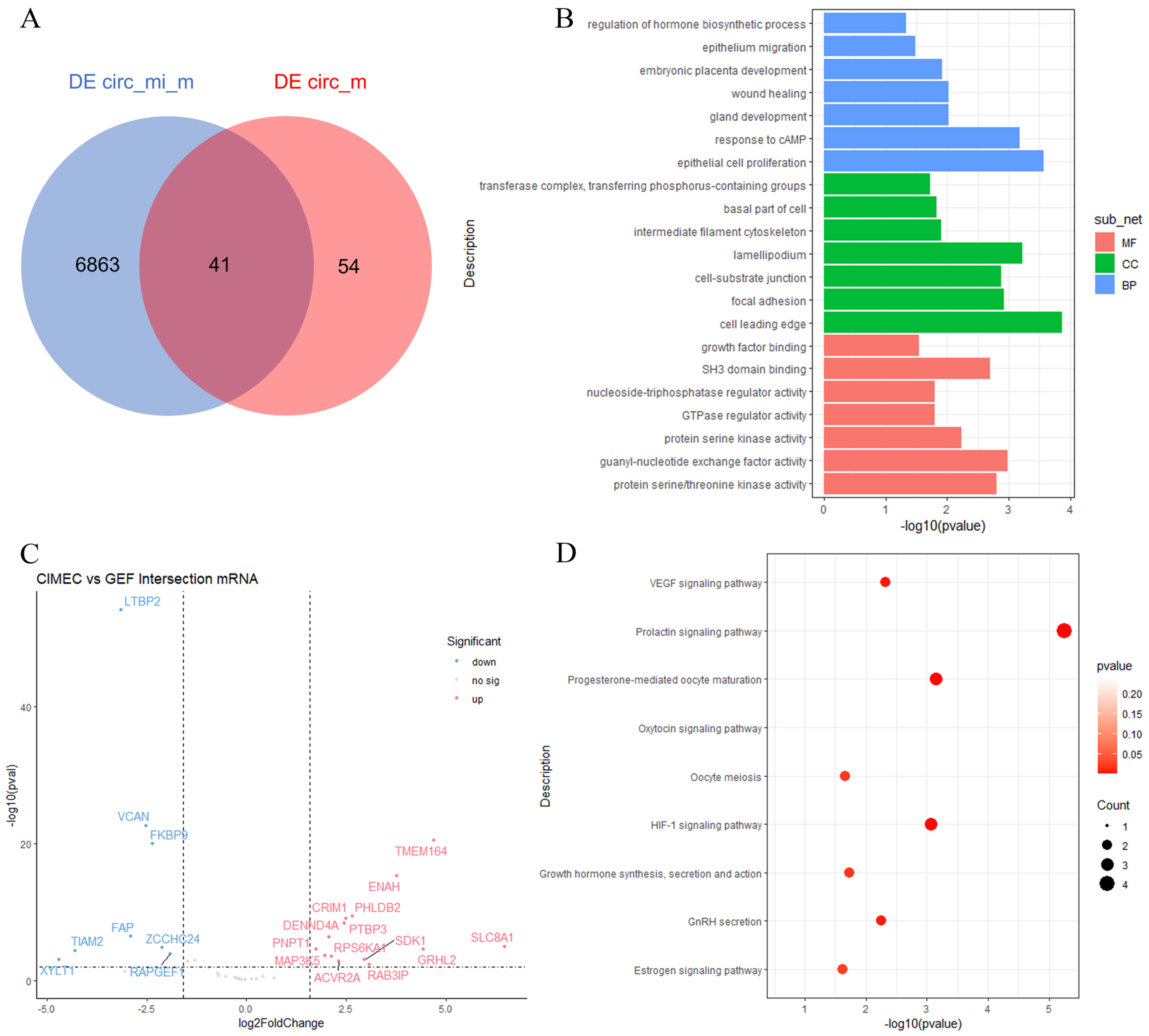
3.4. Prediction of Differentially Expressed circRNA–miRNA–mRNA
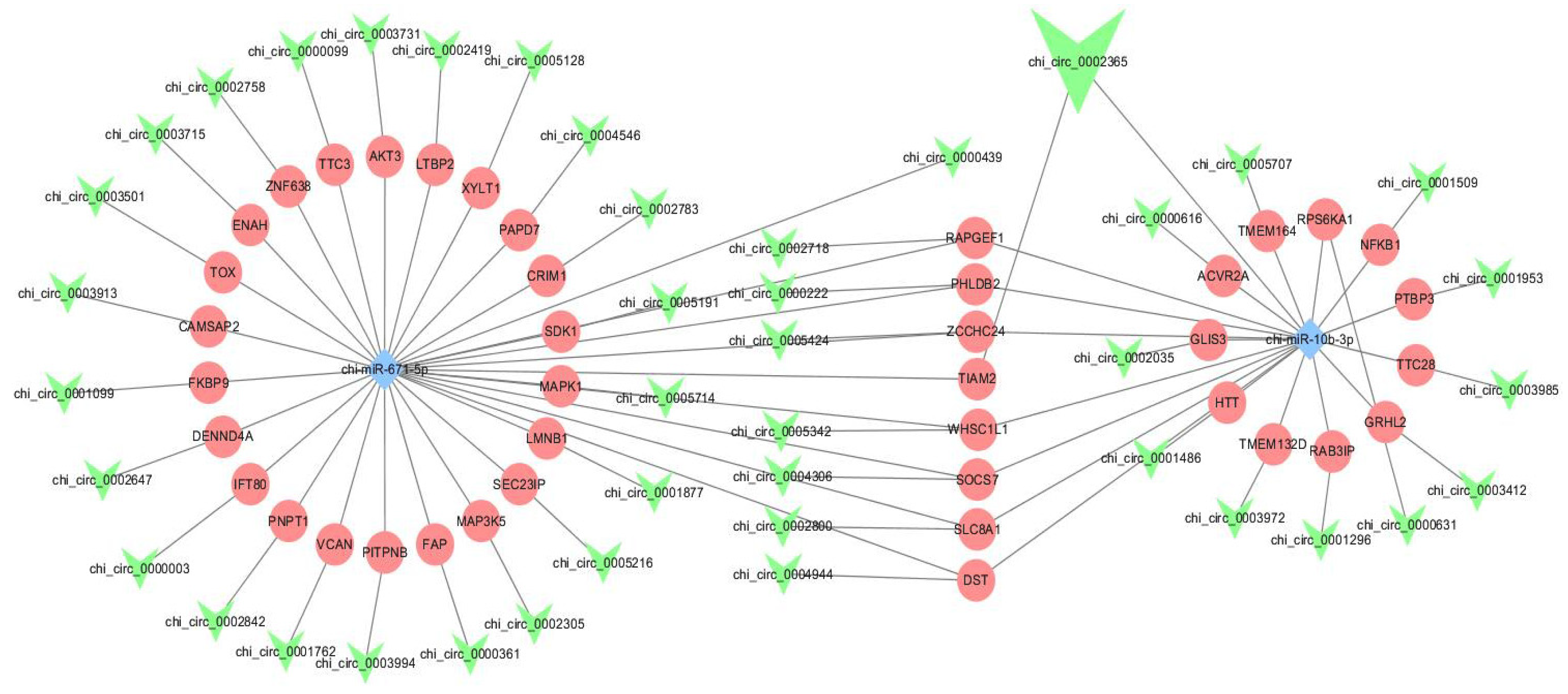
3.5. Establishment of ceRNA Network
4. Discussion
4.1. Establishment of ceRNA Networks for Candidate circRNA’s
4.2. Identification and Detection of Key Candidate circRNA’s
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cherry, A.B.; Daley, G.Q. Reprogramming cellular identity for regenerative medicine. Cell 2012, 148, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Du, Y.; Deng, H. Direct lineage reprogramming: Strategies, mechanisms, and applications. Cell Stem Cell 2015, 16, 119–134. [Google Scholar] [CrossRef]
- Yoshimoto, R.; Rahimi, K.; Hansen, T.B.; Kjems, J.; Mayeda, A. Biosynthesis of circular RNA ciRS-7/CDR1as is mediated by mammalian-wide interspersed repeats. Iscience 2020, 23, 101345. [Google Scholar] [CrossRef]
- Chen, Z.; He, Q.; Lu, T.; Wu, J.; Shi, G.; He, L.; Zong, H.; Liu, B.; Zhu, P. mcPGK1-dependent mitochondrial import of PGK1 promotes metabolic reprogramming and self-renewal of liver TICs. Nat. Commun. 2023, 14, 1121. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hao, Z.; Hu, J.; Liu, X.; Li, S.; Wang, J.; Shen, J.; Song, Y.; Ke, N.; Luo, Y. Small RNA deep sequencing reveals the expressions of microRNAs in ovine mammary gland development at peak-lactation and during the non-lactating period. Genomics 2021, 113, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wu, H.; Wang, Y.; Zhu, S.; Liu, J.; Fang, X.; Chen, H. Circular RNA of cattle casein genes are highly expressed in bovine mammary gland. J. Dairy Sci. 2016, 99, 4750–4760. [Google Scholar] [CrossRef]
- Ma, D.; Zhao, Y.; Yu, S.; Zhang, H.; Cheng, M.; Cao, H.; Li, Q.; Min, L. CircRNA as CeRNA mediated by microRNA may be involved in goat lactation. Small Rumin. Res. 2019, 171, 63–72. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, H.; Wang, Y.; Zhao, Y.; Fang, X.; Chen, C.; Chen, H. Expression patterns of circular RNAs from primary kinase transcripts in the mammary glands of lactating rats. J. Breast Cancer 2015, 18, 235–241. [Google Scholar] [CrossRef]
- Christoforou, N.; Chakraborty, S.; Kirkton, R.D.; Adler, A.F.; Addis, R.C.; Leong, K.W. Core transcription factors, microRNAs, and small molecules drive transdifferentiation of human fibroblasts towards the cardiac cell lineage. Sci. Rep. 2017, 7, 40285. [Google Scholar] [CrossRef]
- Subramanyam, D.; Lamouille, S.; Judson, R.L.; Liu, J.Y.; Bucay, N.; Derynck, R.; Blelloch, R. Multiple targets of miR-302 and miR-372 promote reprogramming of human fibroblasts to induced pluripotent stem cells. Nat. Biotechnol. 2011, 29, 443–448. [Google Scholar] [CrossRef]
- Lee, K.; Yu, P.; Lingampalli, N.; Kim, H.J.; Tang, R.; Murthy, N. Peptide-enhanced mRNA transfection in cultured mouse cardiac fibroblasts and direct reprogramming towards cardiomyocyte-like cells. Int. J. Nanomed. 2015, 10, 1841. [Google Scholar]
- Wang, Y.; Xu, Z.; Jiang, J.; Xu, C.; Kang, J.; Xiao, L.; Wu, M.; Xiong, J.; Guo, X.; Liu, H. Endogenous miRNA sponge lincRNA-RoR regulates Oct4, Nanog, and Sox2 in human embryonic stem cell self-renewal. Dev. Cell 2013, 25, 69–80. [Google Scholar] [CrossRef]
- Zheng, Q.; Bao, C.; Guo, W.; Li, S.; Chen, J.; Chen, B.; Luo, Y.; Lyu, D.; Li, Y.; Shi, G. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat. Commun. 2016, 7, 11215. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, J.; Zheng, K.; Zhang, H.; Pei, X.; Yin, Z.; Wen, D.; Kong, Q. Long noncoding RNAs sustain high expression levels of exogenous octamer-binding protein 4 by sponging regulatory microRNAs during cellular reprogramming. J. Biol. Chem. 2019, 294, 17863–17874. [Google Scholar] [CrossRef]
- Liu, Y.; Xue, M.; Du, S.; Feng, W.; Zhang, K.; Zhang, L.; Liu, H.; Jia, G.; Wu, L.; Hu, X. Competitive endogenous RNA is an intrinsic component of EMT regulatory circuits and modulates EMT. Nat. Commun. 2019, 10, 1637. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, G.; Qin, L.; Liu, Q.; Zhu, S.; Ye, S.; Li, X.; Wu, Y.; Hu, Y.; Liu, S. Restoring mammary gland structures and functions with autogenous cell therapy. Biomaterials 2021, 277, 121075. [Google Scholar] [CrossRef] [PubMed]
- Kulcheski, F.R.; Christoff, A.P.; Margis, R. Circular RNAs are miRNA sponges and can be used as a new class of biomarker. J. Biotechnol. 2016, 238, 42–51. [Google Scholar] [CrossRef]
- Danan, M.; Schwartz, S.; Edelheit, S.; Sorek, R. Transcriptome-wide discovery of circular RNAs in Archaea. Nucleic Acids Res. 2012, 40, 3131–3142. [Google Scholar] [CrossRef]
- Tong, C.; Chen, Q.; Zhao, L.; Ma, J.; Ibeagha-Awemu, E.M.; Zhao, X. Identification and characterization of long intergenic noncoding RNAs in bovine mammary glands. BMC Genom. 2017, 18, 468. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhou, J.; Wang, M.; Liu, J.; Zhang, L.; Loor, J.J.; Liang, Y.; Wu, H.; Yang, Z. Circ09863 regulates unsaturated fatty acid metabolism by adsorbing miR-27a-3p in bovine mammary epithelial cells. J. Agric. Food Chem. 2020, 68, 8589–8601. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Li, Y.; Zhang, A.; Gao, L. Circular RNA circ-BANP regulates oxidized low-density lipoprotein-induced endothelial cell injury through targeting the miR-370/Thioredoxin-interacting protein Axis. J. Cardiovasc. Pharmacol. 2021, 77, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Yettram, A.; Grewal, B.; Gibson, D.; Dawson, J. Relation between intraventricular pressure and volume in diastole. Heart 1990, 64, 304–308. [Google Scholar] [CrossRef]
- Nakaya, Y.; Sukowati, E.W.; Sheng, G. Epiblast integrity requires CLASP and Dystroglycan-mediated microtubule anchoring to the basal cortex. J. Cell Biol. 2013, 202, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Elias, S.; Thion, M.S.; Yu, H.; Sousa, C.M.; Lasgi, C.; Morin, X.; Humbert, S. Huntingtin regulates mammary stem cell division and differentiation. Stem Cell Rep. 2014, 2, 491–506. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Luo, J.; Ma, L.; Wang, H.; Cao, W.; Xu, H.; Zhu, J.; Sun, Y.; Li, J.; Yao, D. MiR130b-Regulation of PPARγ coactivator-1α suppresses fat metabolism in goat mammary epithelial cells. PLoS ONE 2015, 10, e0142809. [Google Scholar] [CrossRef]
- Elias, S.; McGuire, J.R.; Yu, H.; Humbert, S. Huntingtin is required for epithelial polarity through RAB11A-mediated apical trafficking of PAR3-aPKC. PLoS Biol. 2015, 13, e1002142. [Google Scholar] [CrossRef]
- Brantley, D.M.; Yull, F.E.; Muraoka, R.S.; Hicks, D.J.; Cook, C.M.; Kerr, L.D. Dynamic expression and activity of NF-κB during post-natal mammary gland morphogenesis. Mech. Dev. 2000, 97, 149–155. [Google Scholar] [CrossRef]
- Huang, X.; Zang, Y.; Zhang, M.; Yuan, X.; Li, M.; Gao, X. Nuclear factor of κB1 is a key regulator for the transcriptional activation of milk synthesis in bovine mammary epithelial cells. DNA Cell Biol. 2017, 36, 295–302. [Google Scholar] [CrossRef]
- Lu, L.M.; Li, Q.Z.; Huang, J.G.; Gao, X.J. Proteomic and functional analyses reveal MAPK1 regulates milk protein synthesis. Molecules 2012, 18, 263–275. [Google Scholar] [CrossRef]
- Laitinen, L.; Miettinen, A.; Tikkanen, I.; TÖrnroth, T.; Nordling, S. Glomerular sialic acid in Heymann nephritis and diacetylbenzidine induced nephropathy in rats. Clin. Sci. 1985, 69, 57–62. [Google Scholar] [CrossRef]
- Green, K.; Streuli, C. Apoptosis regulation in the mammary gland. Cell. Mol. Life Sci. 2004, 61, 1867–1883. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.M.; Liang, A.; Hua, G.; Talpur, H.S.; Salim, M.; Ahmad, S.; Abulaiti, A.; Khan, M.; Safdar, M.; Kakar, I.U. Adaptive molecular evolution of AKT3 gene for positive diversifying selection in mammals. BioMed Res. Int. 2020, 2020, 2584627. [Google Scholar]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine mammary protein synthesis during the lactation cycle. Bioinform. Biol. Insights 2011, 5. [Google Scholar] [CrossRef]
- Jaiswal, S.; Jagannadham, J.; Kumari, J.; Iquebal, M.A.; Gurjar, A.K.S.; Nayan, V.; Angadi, U.B.; Kumar, S.; Kumar, R.; Datta, T.K. Genome wide prediction, mapping and development of genomic resources of mastitis associated genes in water buffalo. Front. Vet. Sci. 2021, 8, 593871. [Google Scholar] [CrossRef]
- Matsuoka, S.; Suzuki, H.; Kato, C.; Kamikawa-Tokai, M.; Kamikawa, A.; Okamatsu-Ogura, Y.; Kimura, K. Expression of Grainyhead-like 2 in the Process of Ductal Development of Mouse Mammary Gland. J. Histochem. Cytochem. 2021, 69, 373–388. [Google Scholar] [CrossRef]
- Do, D.; Bissonnette, N.; Lacasse, P.; Miglior, F.; Sargolzaei, M.; Zhao, X.; Ibeagha-Awemu, E. Genome-wide association analysis and pathways enrichment for lactation persistency in Canadian Holstein cattle. J. Dairy Sci. 2017, 100, 1955–1970. [Google Scholar] [CrossRef]
- Le Guillou, S.; Marthey, S.; Laloë, D.; Laubier, J.; Mobuchon, L.; Leroux, C.; Le Provost, F. Characterisation and comparison of lactating mouse and bovine mammary gland miRNomes. PLoS ONE 2014, 9, e91938. [Google Scholar] [CrossRef]
- Lin, X.; Luo, J.; Zhang, L.; Wang, W.; Gou, D. MiR-103 controls milk fat accumulation in goat (Capra hircus) mammary gland during lactation. PLoS ONE 2013, 8, e79258. [Google Scholar] [CrossRef]
- Huang, L.; Yin, Z.; Feng, Y.; Zhang, X.; Wu, T.; Ding, Y.; Ye, P.; Fu, K.; Zhang, M. Identification and differential expression of micro RNA s in the ovaries of pigs (Sus scrofa) with high and low litter sizes. Anim. Genet. 2016, 47, 543–551. [Google Scholar] [CrossRef]
- Chen, Z.; Qiu, H.; Ma, L.; Luo, J.; Sun, S.; Kang, K.; Gou, D.; Loor, J.J. miR-30e-5p and miR-15a synergistically regulate fatty acid metabolism in goat mammary epithelial cells via LRP6 and YAP1. Int. J. Mol. Sci. 2016, 17, 1909. [Google Scholar] [CrossRef]
- Hartmann, D.L.; Tank, A.M.G.K.; Rusticucci, M.; Alexander, L.V.; Brönnimann, S.; Charabi, Y.A.R.; Dentener, F.J.; Dlugokencky, E.J.; Easterling, D.R.; Kaplan, A. Observations: Atmosphere and surface. In Climate Change 2013 the Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013; pp. 159–254. [Google Scholar]
- Capra, E.; Lazzari, B.; Frattini, S.; Chessa, S.; Coizet, B.; Talenti, A.; Castiglioni, B.; Marsan, P.A.; Crepaldi, P.; Pagnacco, G. Distribution of ncRNAs expression across hypothalamic-pituitary-gonadal axis in Capra hircus. BMC Genom. 2018, 19, 417. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Zhao, F.Q. Emerging evidence of the physiological role of hypoxia in mammary development and lactation. J. Anim. Sci. Biotechnol. 2014, 5, 9. [Google Scholar] [CrossRef]
- Benmoussa, A.; Laugier, J.; Beauparlant, C.J.; Lambert, M.; Droit, A.; Provost, P. Complexity of the microRNA transcriptome of cow milk and milk-derived extracellular vesicles isolated via differential ultracentrifugation. J. Dairy Sci. 2020, 103, 16–29. [Google Scholar] [CrossRef]
- Baran, N.; Kelly, P.A.; Binart, N. Characterization of a Prolactin-Regulated Gene in Reproductive Tissues Usingthe Prolactin Receptor Knockout Mouse Model. Biol. Reprod. 2002, 66, 1210–1218. [Google Scholar] [CrossRef]
- Eyal, O.; Jomain, J.B.; Kessler, C.; Goffin, V.; Handwerger, S. Autocrine prolactin inhibits human uterine decidualization: A novel role for prolactin. Biol. Reprod. 2007, 76, 777–783. [Google Scholar] [CrossRef]
- Cordero, A.; Pellegrini, P.; Sanz-Moreno, A.; Trinidad, E.M.; Serra-Musach, J.; Deshpande, C.; Dougall, W.C.; Pujana, M.A.; González-Suárez, E. Rankl impairs lactogenic differentiation through inhibition of the prolactin/Stat5 pathway at midgestation. Stem Cells 2016, 34, 1027–1039. [Google Scholar] [CrossRef]
- Macias, H.; Hinck, L. Mammary gland development. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 533–557. [Google Scholar] [CrossRef]
- Barrera, D.; Avila, E.; Díaz, L. Immunological role of progesterone in the maintenance of pregnancy. Rev. De Investig. Clin. 2007, 59, 139–145. [Google Scholar]
- Kashimata, M.; Sayeed, S.; Ka, A.; Onetti-Muda, A.; Sakagami, H.; Faraggiana, T.; Gresik, E.W. The ERK-1/2 signaling pathway is involved in the stimulation of branching morphogenesis of fetal mouse submandibular glands by EGF. Dev. Biol. 2000, 220, 183–196. [Google Scholar] [CrossRef]
- Fata, J.E.; Mori, H.; Ewald, A.J.; Zhang, H.; Yao, E.; Werb, Z.; Bissell, M.J. The MAPKERK-1, 2 pathway integrates distinct and antagonistic signals from TGFα and FGF7 in morphogenesis of mouse mammary epithelium. Dev. Biol. 2007, 306, 193–207. [Google Scholar] [CrossRef]
- Russo, J.; Ao, X.; Grill, C.; Russo, I. Pattern of distribution of cells positive for estrogen receptor α and progesterone receptor in relation to proliferating cells in the mammary gland. Breast Cancer Res. Treat. 1999, 53, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Wilde, C.J.; Knight, C.H. Milk yield and mammary function in goats during and after once-daily milking. J. Dairy Res. 1990, 57, 441–447. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahito, J.Z.A.; Deng, S.; Qin, L.; Xiao, L.; Zhang, D.; Huang, B. CeRNA Network Reveals the Circular RNA Characterization in Goat Ear Fibroblasts Reprogramming into Mammary Epithelial Cells. Genes 2023, 14, 1831. https://doi.org/10.3390/genes14101831
Sahito JZA, Deng S, Qin L, Xiao L, Zhang D, Huang B. CeRNA Network Reveals the Circular RNA Characterization in Goat Ear Fibroblasts Reprogramming into Mammary Epithelial Cells. Genes. 2023; 14(10):1831. https://doi.org/10.3390/genes14101831
Chicago/Turabian StyleSahito, Jam Zaheer Ahmed, Shan Deng, Liangshan Qin, Lianggui Xiao, Dandan Zhang, and Ben Huang. 2023. "CeRNA Network Reveals the Circular RNA Characterization in Goat Ear Fibroblasts Reprogramming into Mammary Epithelial Cells" Genes 14, no. 10: 1831. https://doi.org/10.3390/genes14101831