
Comparative Transcriptomic Assessment of Chemosensory Genes in Adult and Larval Olfactory Organs of Cnaphalocrocis medinalis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Specimen Preparation
2.2. Illumina Sequencing, Sequence Assembly, Function Annotation, and Expression Abundance Analysis
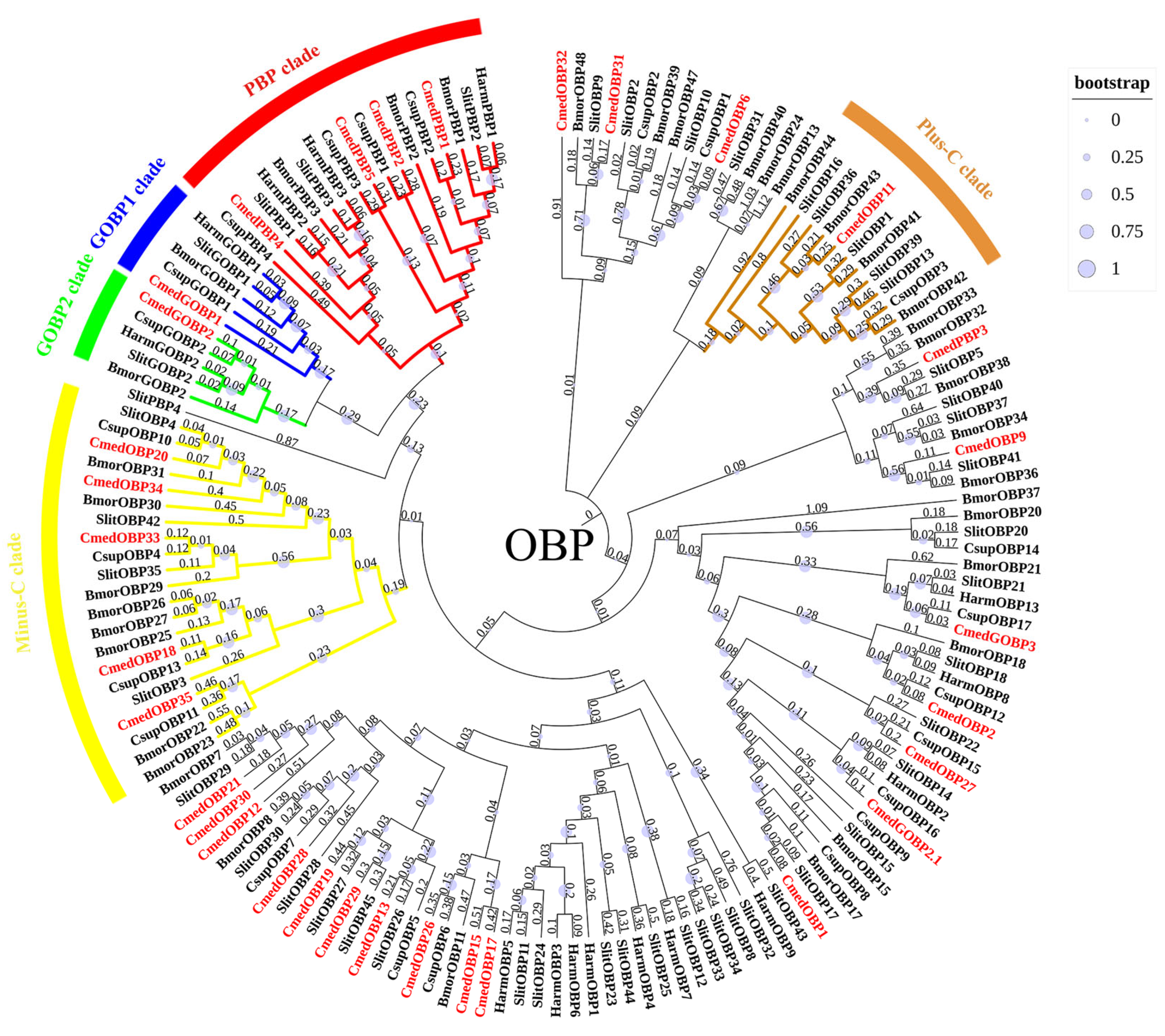
2.3. Identification and Phylogenetic Analyses of Chemosensory Genes
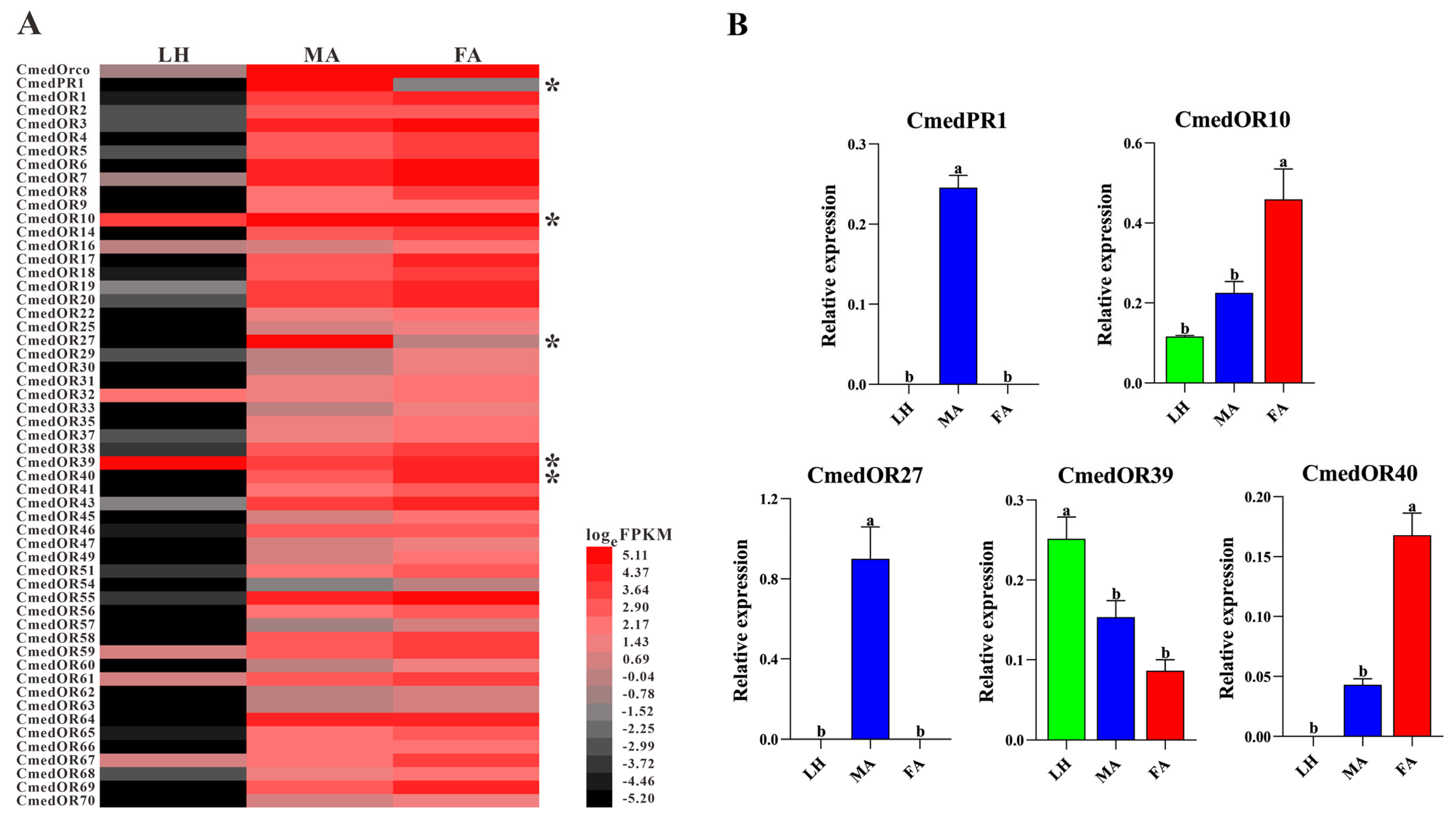
2.4. Quantitative Real-Time Polymerase Chain Reaction Validation
3. Results
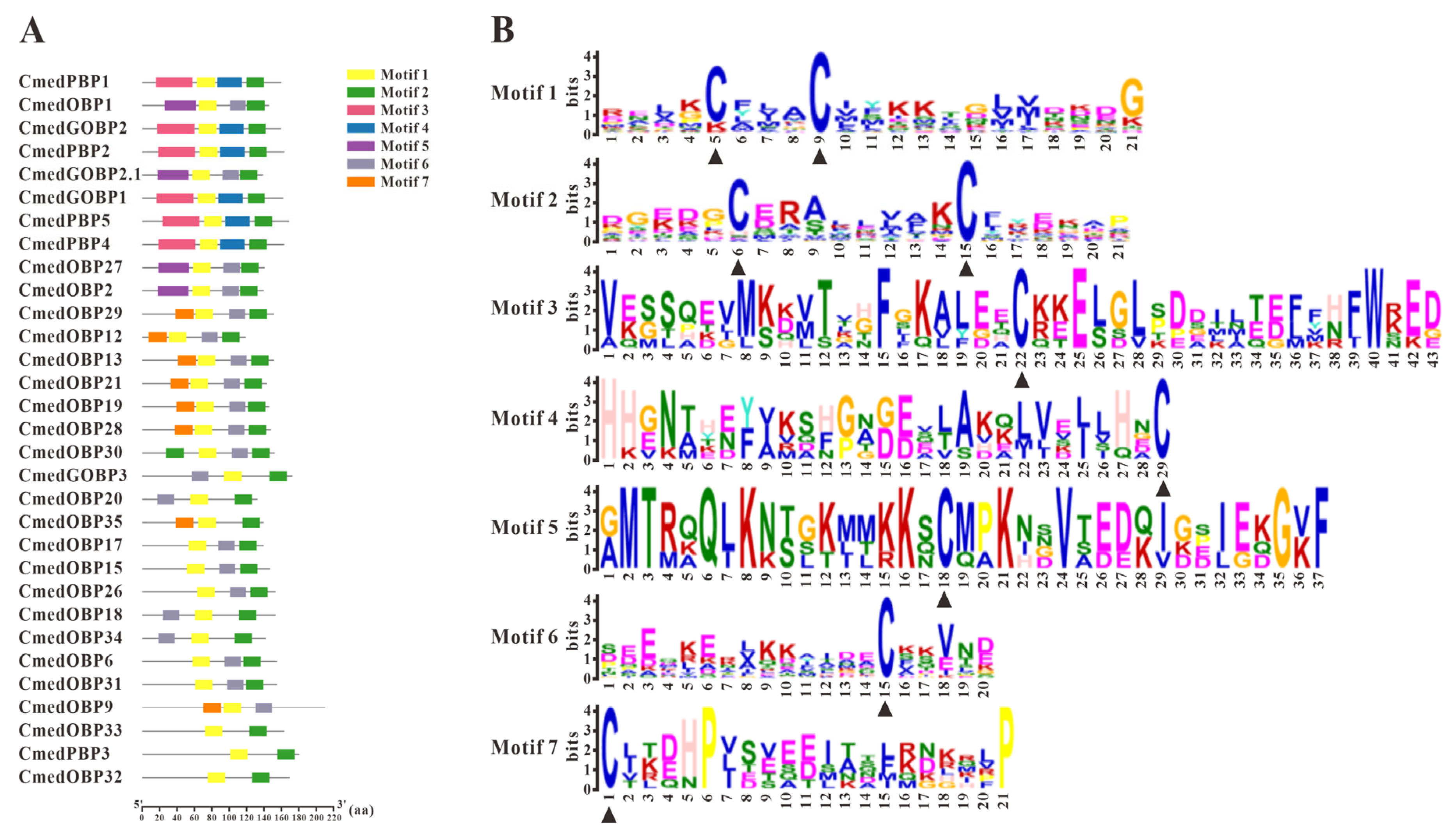
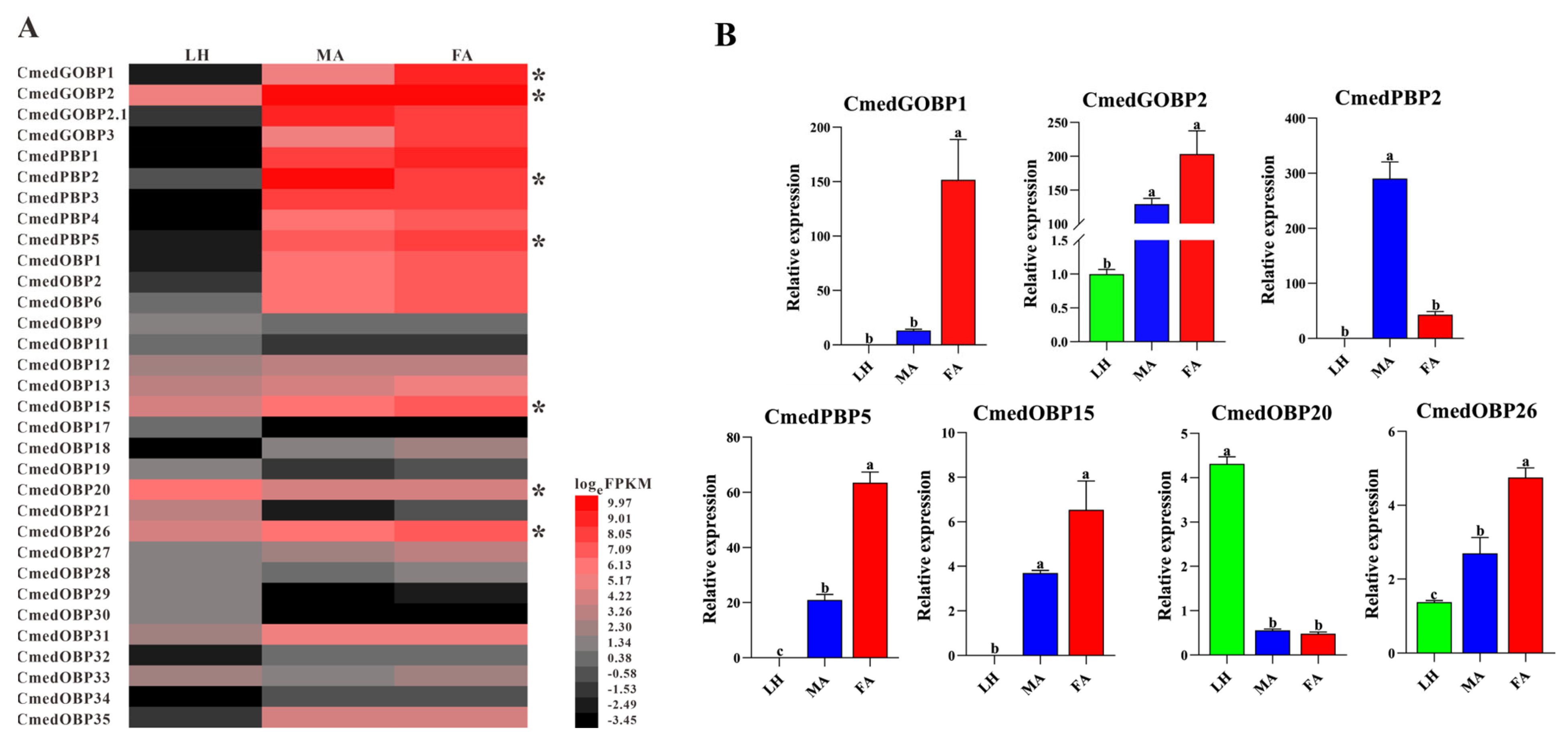
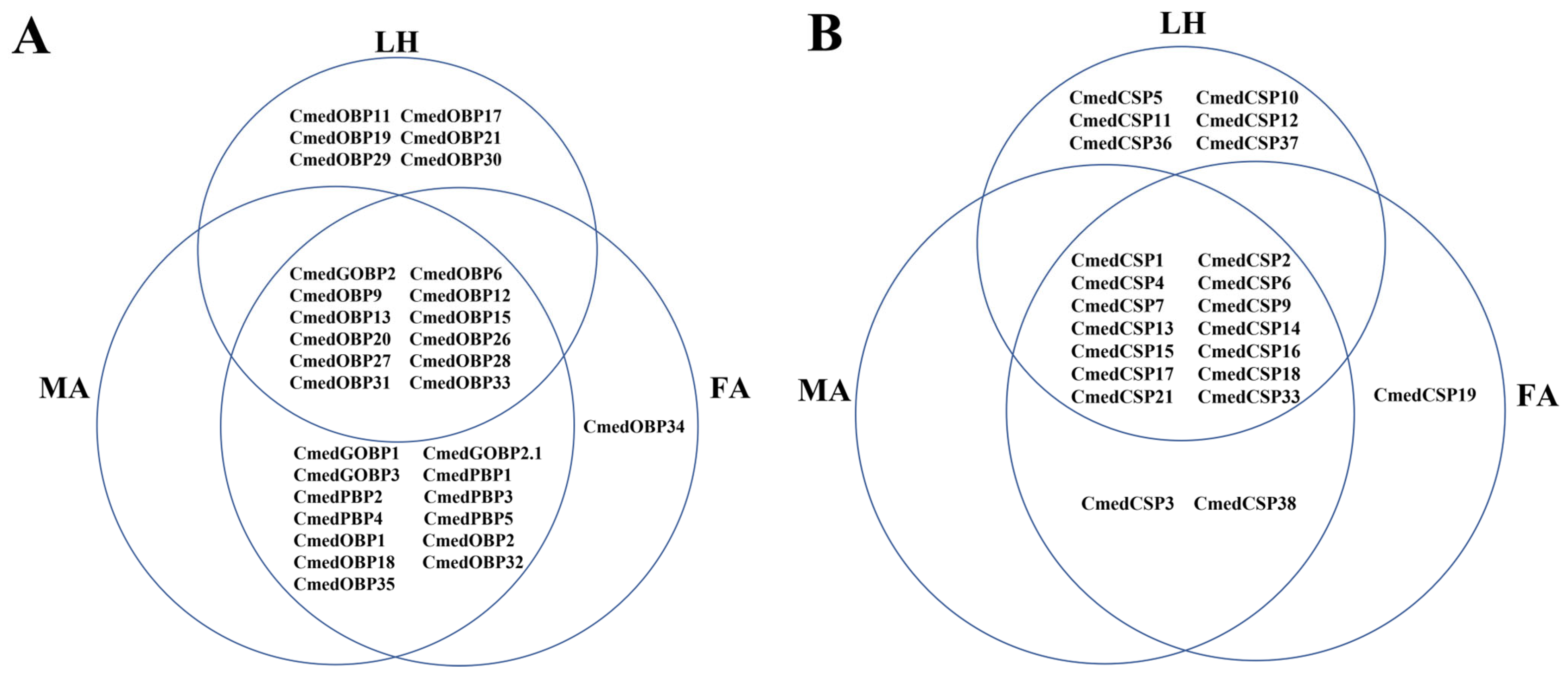
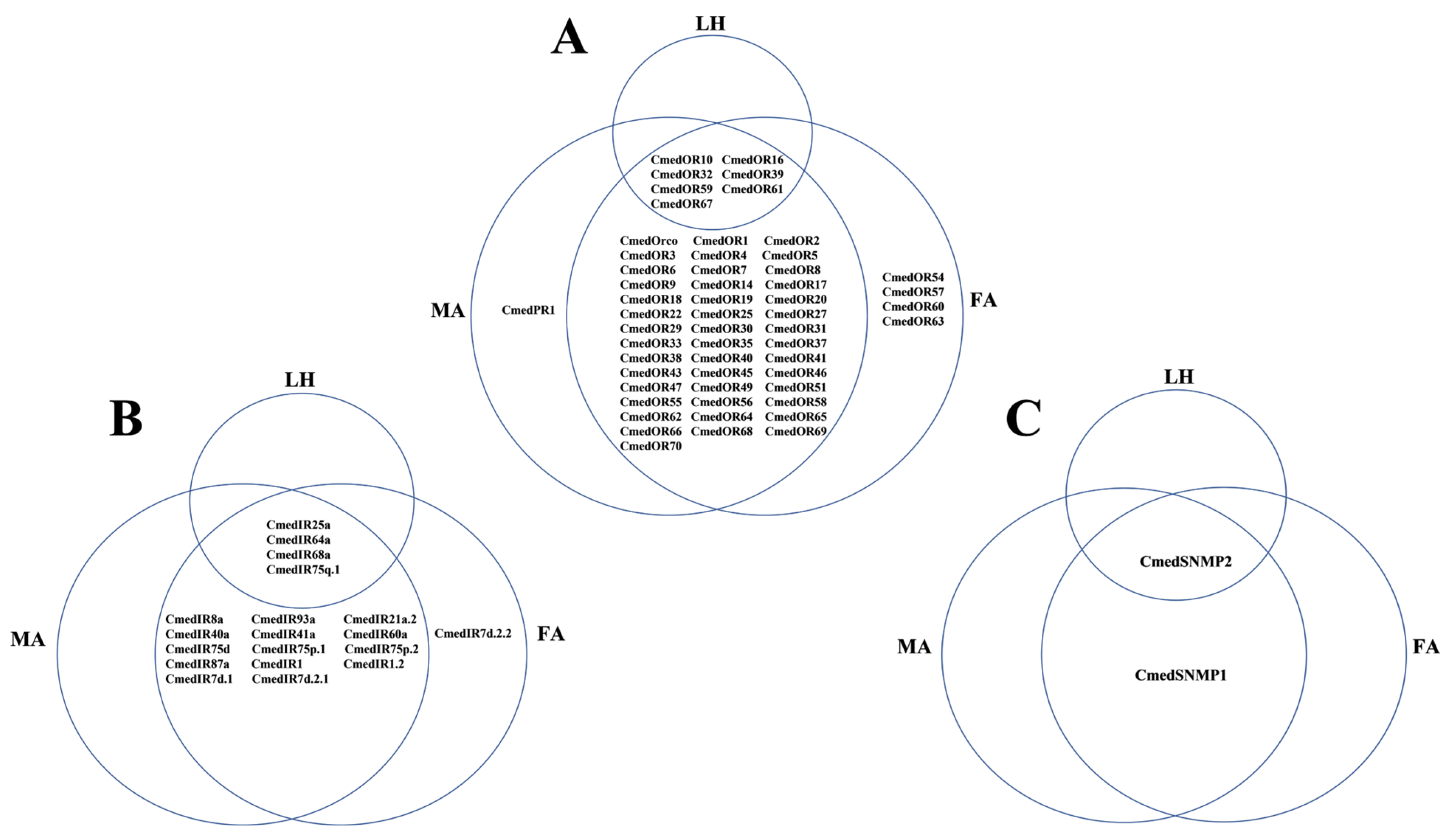
3.1. Identification and Comparison of Odorant-Binding Proteins in C. medinalis Larval and Adult Olfactory Organs
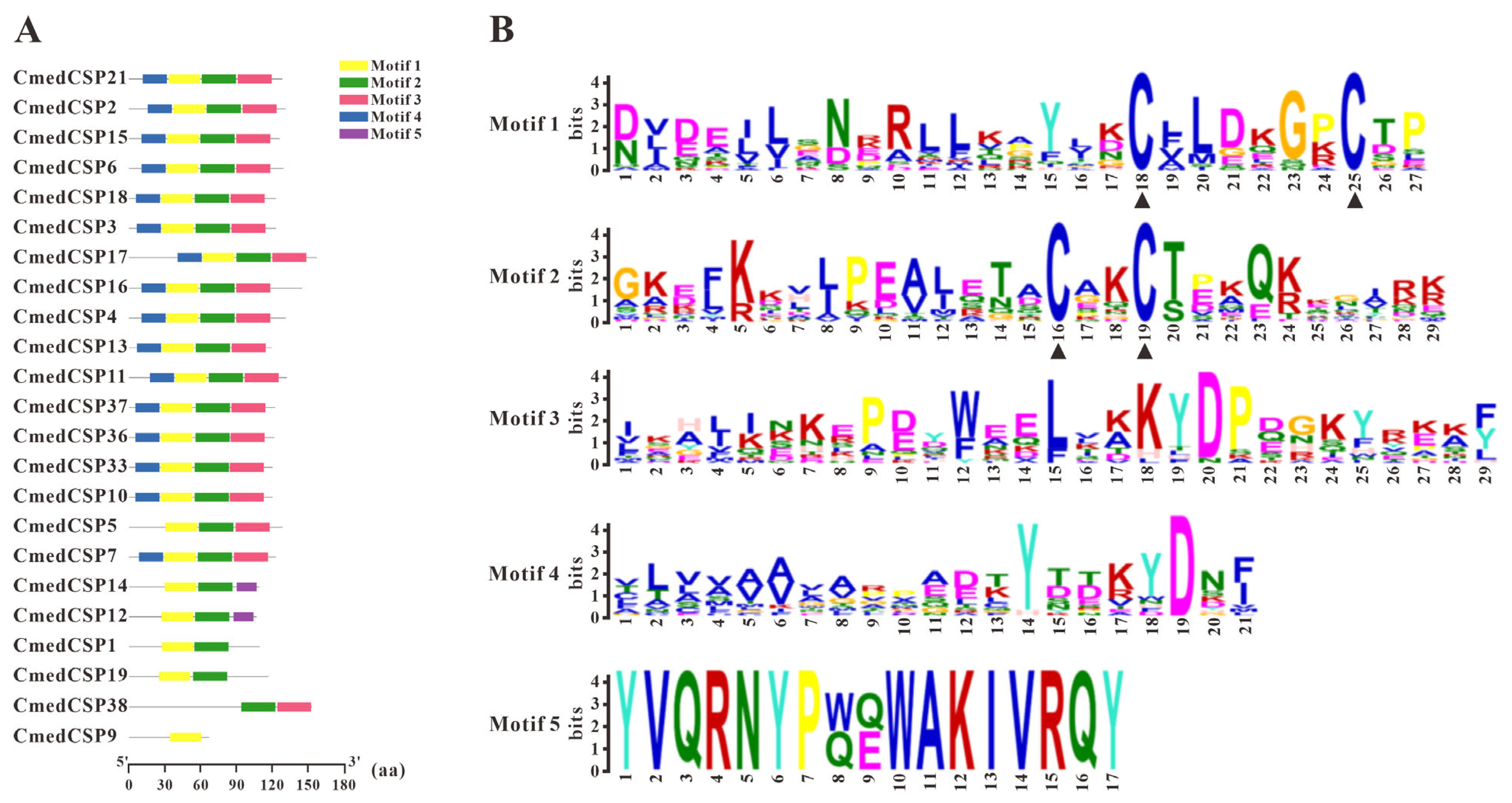
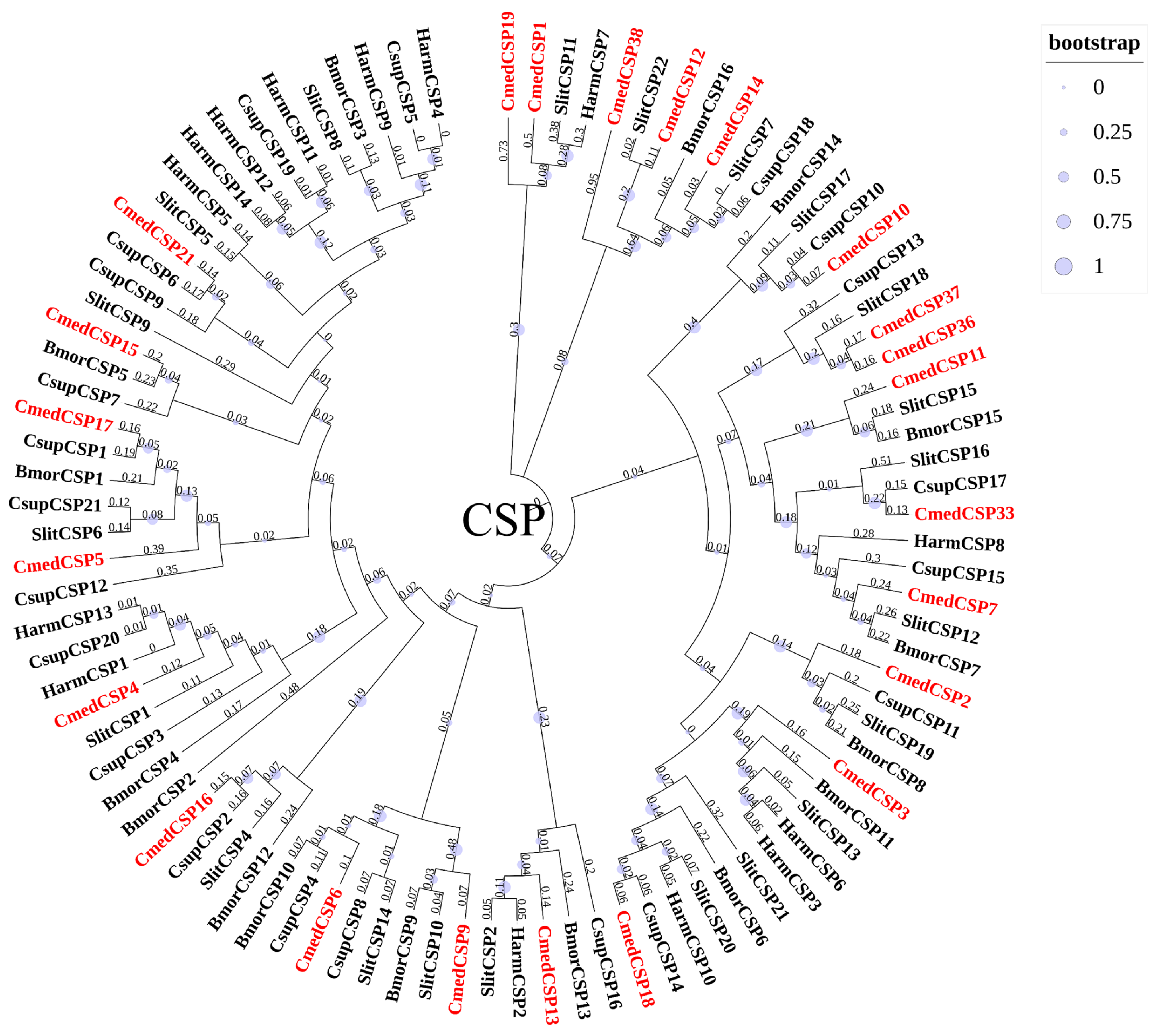
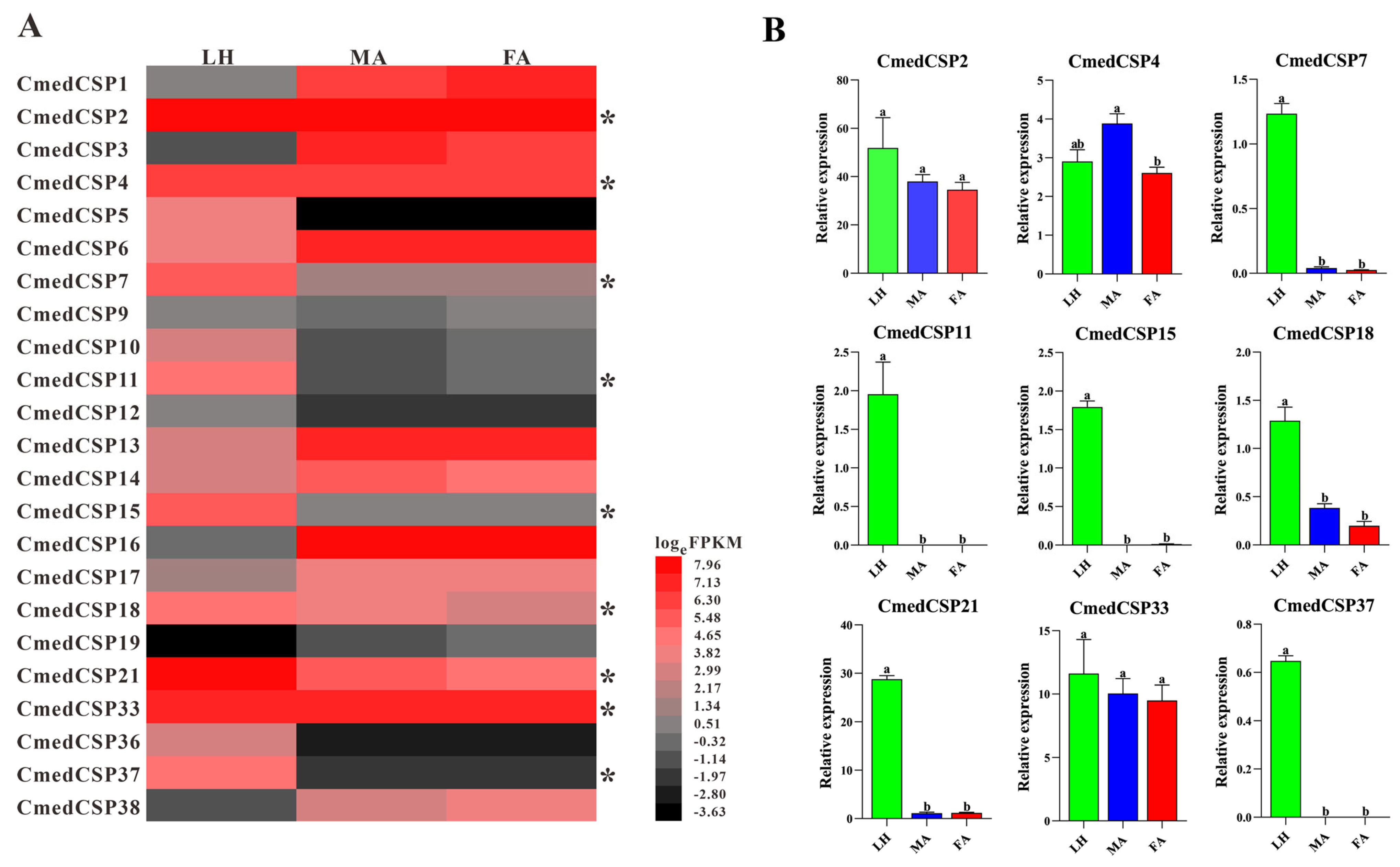
3.2. Identification and Comparison of Chemosensory Proteins in C. medinalis Larval and Adult Olfactory Organs
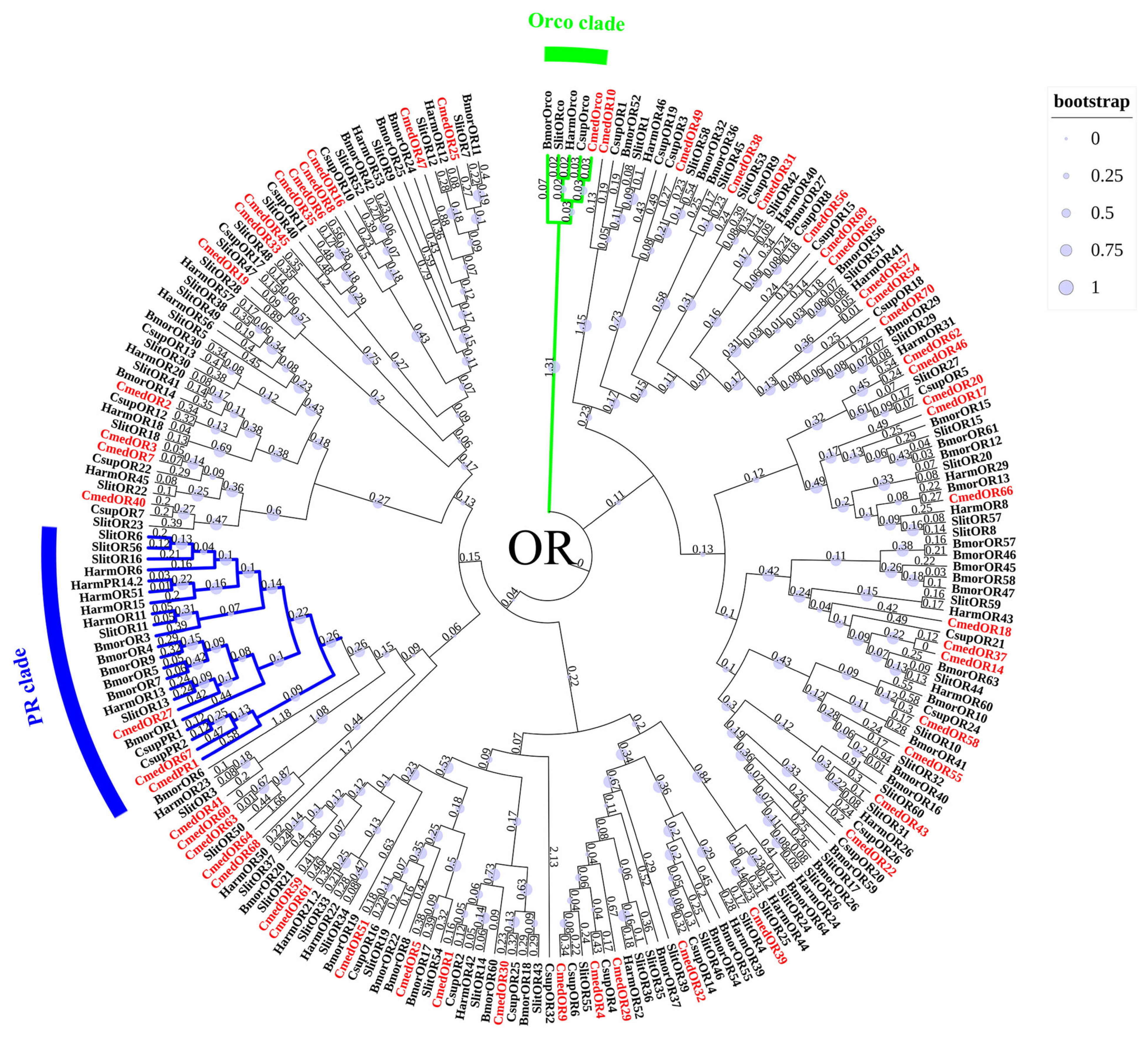
3.3. Identification and Comparison of Odorant Receptors in C. medinalis Larval and Adult Olfactory Organs
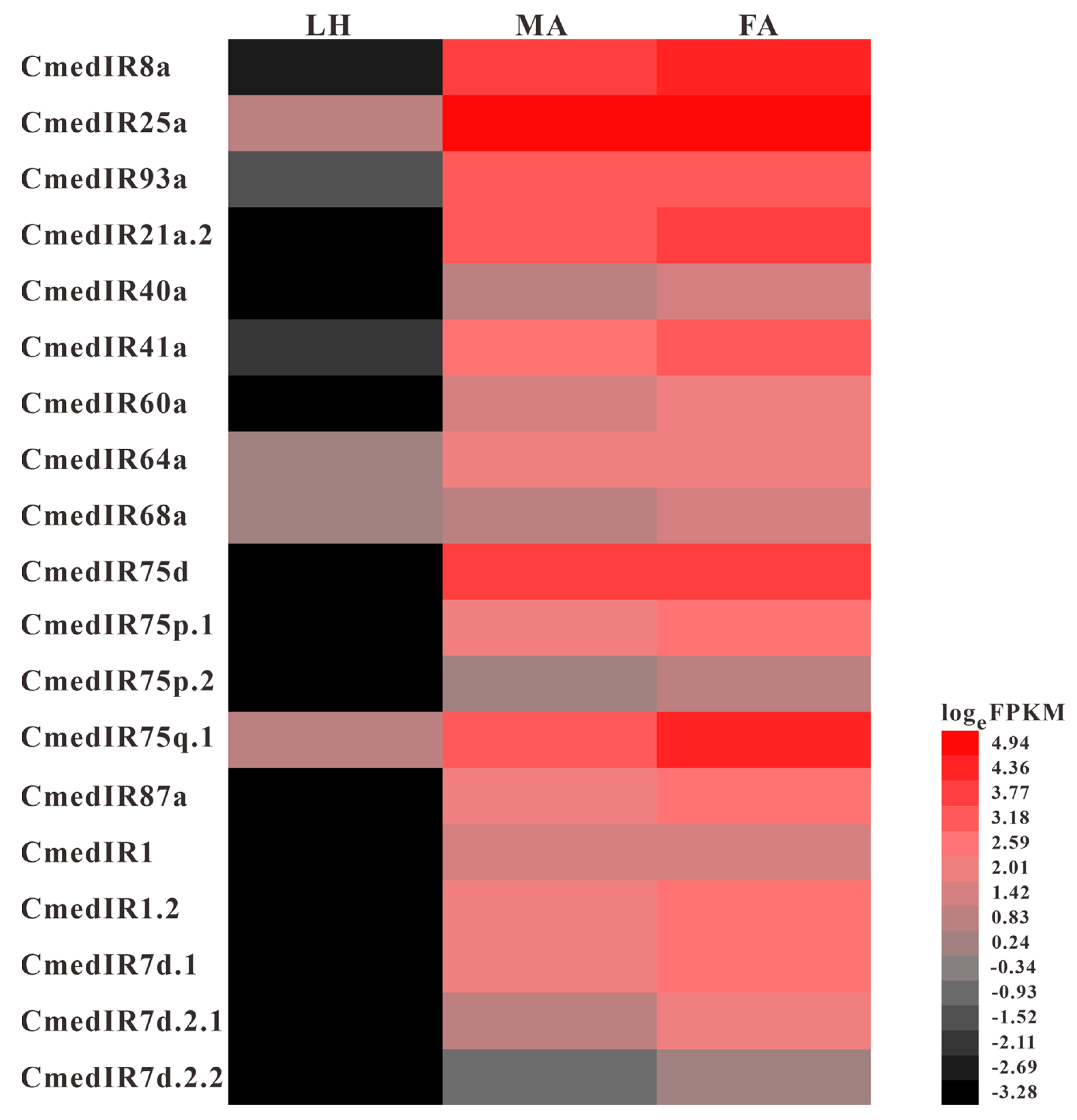
3.4. Identification and Comparison of Ionotropic Receptors and Sensory Neuron Membrane Proteins in C. medinalis Larval and Adult Olfactory Organs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Renou, M.; Anton, S. Insect olfactory communication in a complex and changing world. Curr. Opin. Insect Sci. 2020, 42, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Keil, T.A. Morphology and development of the peripheral olfactory organs. In Insect Olfaction; Springer: Berlin, Germany, 1999; pp. 5–47. [Google Scholar]
- Pelosi, P.; Zhou, J.J.; Ban, L.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef] [PubMed]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Scaloni, A.; Monti, M.; Angeli, S.; Pelosi, P. Structural analysis and disulfide-bridge pairing of two odorant-binding proteins from Bombyx mori. Biochem. Biophys. Res. Commun. 1999, 266, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Iovinella, I.; Felicioli, A.; Dani, F.R. Soluble proteins of chemical communication: An overview across arthropods. Front. Physiol. 2014, 5, 320. [Google Scholar] [CrossRef]
- Mombaerts, P. Seven-transmembrane proteins as odorant and chemosensory receptors. Science 1999, 286, 707–711. [Google Scholar] [CrossRef]
- Rytz, R.; Croset, V.; Benton, R. Ionotropic receptors (IRs): Chemosensory ionotropic glutamate receptors in Drosophila and beyond. Insect Biochem. Mol. Biol. 2013, 43, 888–897. [Google Scholar] [CrossRef]
- Rogers, M.E.; Krieger, J.; Vogt, R.G. Antennal SNMPs (sensory neuron membrane proteins) of Lepidoptera define a unique family of invertebrate CD36-like proteins. J. Neurobiol. 2001, 49, 47–61. [Google Scholar] [CrossRef]
- Nakagawa, T.; Vosshall, L.B. Controversy and consensus: Noncanonical signaling mechanisms in the insect olfactory system. Curr. Opin. Neurobiol. 2009, 19, 284–292. [Google Scholar] [CrossRef]
- Skiri, H.T.; Rø, H.; Berg, B.G.; Mustaparta, H. Consistent organization of glomeruli in the antennal lobes of related species of heliothine moths. J. Comp. Neurol. 2005, 491, 367–380. [Google Scholar] [CrossRef]
- Hansson, B.S.; Stensmyr, M.C. Evolution of insect olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef] [PubMed]
- Knolhoff, L.M.; Heckel, D.G. Behavioral assays for studies of host plant choice and adaptation in herbivorous insects. Annu. Rev. Entomol. 2014, 59, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Dethier, V.; Schoonhoven, L. Olfactory coding by lepidopterous larvae. Entomol. Exp. Appl. 1969, 12, 535–543. [Google Scholar] [CrossRef]
- Ljungberg, H.; Anderson, P.; Hansson, B. Physiology and morphology of pheromone-specific sensilla on the antennae of male and female Spodoptera littoralis (Lepidoptera: Noctuidae). J. Insect Physiol. 1993, 39, 253–260. [Google Scholar] [CrossRef]
- Walker III, W.B.; Mori, B.A.; Cattaneo, A.M.; Gonzalez, F.; Witzgall, P.; Becher, P.G. Comparative transcriptomic assessment of the chemosensory receptor repertoire of Drosophila suzukii adult and larval olfactory organs. Comp. Biochem. Physiol. Part D Genom. Proteom. 2023, 45, 101049. [Google Scholar] [CrossRef] [PubMed]
- Bohbot, J.; Pitts, R.; Kwon, H.W.; Rützler, M.; Robertson, H.; Zwiebel, L. Molecular characterization of the Aedes aegypti odorant receptor gene family. Insect Mol. Biol. 2007, 16, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Glaser, N.; Gallot, A.; Legeai, F.; Montagné, N.; Poivet, E.; Harry, M.; Calatayud, P.A.; Jacquin-Joly, E. Candidate chemosensory genes in the stemborer Sesamia nonagrioides. Int. J. Biol. Sci 2013, 9, 481–495. [Google Scholar] [CrossRef]
- Walker, W.B.; Gonzalez, F.; Garczynski, S.F.; Witzgall, P. The chemosensory receptors of codling moth Cydia pomonella–expression in larvae and adults. Sci. Rep. 2016, 6, 23518. [Google Scholar] [CrossRef]
- Wei, H.S.; Li, K.B.; Zhang, S.; Cao, Y.Z.; Yin, J. Identification of candidate chemosensory genes by transcriptome analysis in Loxostege sticticalis Linnaeus. PLoS ONE 2017, 12, e0174036. [Google Scholar] [CrossRef]
- Poivet, E.; Gallot, A.; Montagné, N.; Glaser, N.; Legeai, F.; Jacquin-Joly, E. A comparison of the olfactory gene repertoires of adults and larvae in the noctuid moth Spodoptera littoralis. PLoS ONE 2013, 8, e60263. [Google Scholar] [CrossRef]
- Tanaka, K.; Uda, Y.; Ono, Y.; Nakagawa, T.; Suwa, M.; Yamaoka, R.; Touhara, K. Highly Selective Tuning of a Silkworm Olfactory Receptor to a Key Mulberry Leaf Volatile. Curr. Biol. 2009, 19, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Barrion, A.; Litsinger, J.; Castilla, N.; Joshi, R. A bibliography of rice leaffolders (Lepidoptera: Pyralidae). Int. J. Trop. Insect Sc. 1988, 9, 129–174. [Google Scholar] [CrossRef]
- Padmavathi, C.; Katti, G.; Padmakumari, A.; Voleti, S.; Subba Rao, L. The effect of leaffolder Cnaphalocrocis medinalis (Guenee) [Lepidoptera: Pyralidae] injury on the plant physiology and yield loss in rice. J. Appl. Entomol. 2013, 137, 249–256. [Google Scholar] [CrossRef]
- Zeng, F.F.; Zhao, Z.F.; Yan, M.J.; Zhou, W.; Zhang, Z.; Zhang, A.; Lu, Z.X.; Wang, M.Q. Identification and comparative expression profiles of chemoreception genes revealed from major chemoreception organs of the rice leaf folder, Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). PLoS ONE 2015, 10, e0144267. [Google Scholar] [CrossRef]
- Liu, S.; Wang, W.L.; Zhang, Y.X.; Zhang, B.X.; Rao, X.J.; Liu, X.M.; Wang, D.M.; Li, S.G. Transcriptome sequencing reveals abundant olfactory genes in the antennae of the rice leaffolder, Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). Entomol. Sci. 2017, 20, 177–188. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Consortium, G.O. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar] [CrossRef]
- Bateman, A.; Coin, L.; Durbin, R.; Finn, R.D.; Hollich, V.; Griffiths-Jones, S.; Khanna, A.; Marshall, M.; Moxon, S.; Sonnhammer, E.L. The Pfam protein families database. Nucleic Acids Res. 2004, 32, D138–D141. [Google Scholar] [CrossRef]
- Wilson, D.; Madera, M.; Vogel, C.; Chothia, C.; Gough, J. The SUPERFAMILY database in 2007: Families and functions. Nucleic Acids Res. 2007, 35, D308–D313. [Google Scholar] [CrossRef]
- Roberts, A.; Pimentel, H.; Trapnell, C.; Pachter, L. Identification of novel transcripts in annotated genomes using RNA-Seq. Bioinformatics 2011, 27, 2325–2329. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, J.; Xiang, Y.; Zhang, Y.; Chen, D.; Liu, F. Identification and comparative expression profiles of chemosensory genes in major chemoreception organs of a notorious pests, Laodelphax striatellus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 33, 100646. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.B.; Roy, A.; Anderson, P.; Schlyter, F.; Hansson, B.S.; Larsson, M.C. Transcriptome Analysis of Gene Families Involved in Chemosensory Function in Spodoptera littoralis (Lepidoptera: Noctuidae). BMC Genom. 2019, 20, 428. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.P.; Liu, Y.; Wei, J.J.; Liao, X.Y.; Walker, W.B.; Li, J.H.; Wang, G.R. Identification of candidate olfactory genes in Chilo suppressalis by antennal transcriptome analysis. Int. J. Biol. Sci. 2014, 10, 846–860. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, B.; Dong, S.L.; Cao, D.P.; Dong, J.F.; Walker, W.B.; Liu, Y.; Wang, G.R. Antennal transcriptome analysis and comparison of chemosensory gene families in two closely related noctuidae moths, Helicoverpa armigera and H. assulta. PLoS ONE 2015, 10, e0117054. [Google Scholar] [CrossRef]
- Chang, H.; Ai, D.; Zhang, J.; Dong, S.; Wang, G. Candidate odorant binding proteins and chemosensory proteins in the larval chemosensory tissues of two closely related noctuidae moths, Helicoverpa armigera and H. assulta. PLoS ONE 2017, 12, e0179243. [Google Scholar] [CrossRef]
- Liu, N.Y.; Zhang, T.; Ye, Z.F.; Li, F.; Dong, S.L. Identification and Characterization of Candidate Chemosensory Gene Families from Spodoptera exigua Developmental Transcriptomes. Int. J. Biol. Sci. 2015, 11, 1036–1048. [Google Scholar] [CrossRef]
- Llopis-Giménez, A.; Carrasco-Oltra, T.; Jacquin-Joly, E.; Herrero, S.; Crava, C.M. Coupling transcriptomics and behaviour to unveil the olfactory system of Spodoptera exigua larvae. J. Chem. Ecol. 2020, 46, 1017–1031. [Google Scholar] [CrossRef]
- Gong, D.P.; Zhang, H.j.; Zhao, P.; Lin, Y.; Xia, Q.Y.; Xiang, Z.H. Identification and expression pattern of the chemosensory protein gene family in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2007, 37, 266–277. [Google Scholar] [CrossRef]
- Maleszka, J.; Forêt, S.; Saint, R.; Maleszka, R. RNAi-induced phenotypes suggest a novel role for a chemosensory protein CSP5 in the development of embryonic integument in the honeybee (Apis mellifera). Dev. Genes Evol. 2007, 217, 189–196. [Google Scholar] [CrossRef] [PubMed]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, H.-T.; Lu, J.-Q.; Ji, K.; Wang, C.-C.; Yao, Z.-C.; Liu, F.; Li, Y. Comparative Transcriptomic Assessment of Chemosensory Genes in Adult and Larval Olfactory Organs of Cnaphalocrocis medinalis. Genes 2023, 14, 2165. https://doi.org/10.3390/genes14122165
Du H-T, Lu J-Q, Ji K, Wang C-C, Yao Z-C, Liu F, Li Y. Comparative Transcriptomic Assessment of Chemosensory Genes in Adult and Larval Olfactory Organs of Cnaphalocrocis medinalis. Genes. 2023; 14(12):2165. https://doi.org/10.3390/genes14122165
Chicago/Turabian StyleDu, Hai-Tao, Jia-Qi Lu, Kun Ji, Chu-Chu Wang, Zhi-Chao Yao, Fang Liu, and Yao Li. 2023. "Comparative Transcriptomic Assessment of Chemosensory Genes in Adult and Larval Olfactory Organs of Cnaphalocrocis medinalis" Genes 14, no. 12: 2165. https://doi.org/10.3390/genes14122165