
Variants of IFNL4 Gene in Amazonian and Northern Brazilian Populations
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Variants
2.2. Data Analysis
2.3. Study Populations
2.4. DNA Extraction and Exome Library Preparation
2.5. Bioinformatics Analysis
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Syedbasha, M.; Egli, A. Interferon Lambda: Modulating Immunity in Infectious Diseases. Front. Immunol. 2017, 8, 119. [Google Scholar]
- Hemann, E.A.; Gale, M.; Savan, R. Interferon Lambda Genetics and Biology in Regulation of Viral Control. Front. Immunol. 2017, 8, 1707. [Google Scholar] [CrossRef]
- Ye, L.; Schnepf, D.; Staeheli, P. Interferon-λ orchestrates innate and adaptive mucosal immune responses. Nat. Rev. Immunol. 2019, 19, 614–625. [Google Scholar] [CrossRef]
- Bruening, J.; Weigel, B.; Gerold, G. The Role of Type III Interferons in Hepatitis C Virus Infection and Therapy. J. Immunol. Res. 2017, 2017, 7232361. [Google Scholar] [CrossRef]
- O’Brien, T.R.; Prokunina-Olsson, L.; Donnelly, R.P. IFN-λ4: The Paradoxical New Member of the Interferon Lambda Family. J. Interferon Cytokine Res. 2014, 34, 829–838. [Google Scholar] [CrossRef]
- Fang, M.Z.; Jackson, S.S.; O’Brien, T.R. IFNL4: Notable variants and associated phenotypes. Gene 2020, 730, 144289. [Google Scholar] [CrossRef]
- Rugwizangoga, B.; Andersson, M.E.; Kabayiza, J.C.; Nilsson, M.S.; Ármannsdóttir, B.; Aurelius, J.; Nilsson, S.; Hellstrand, K.; Lindh, M.; Martner, A. IFNL4 Genotypes Predict Clearance of RNA Viruses in Rwandan Children with Upper Respiratory Tract Infections. Front. Cell. Infect. Microbiol. 2019, 9, 340. [Google Scholar] [CrossRef]
- Bhushan, A.; Chinnaswamy, S. Identifying causal variants at the interferon lambda locus in case-control studies: Utilizing non-synonymous variant rs117648444 to probe the role of IFN-λ4. Gene 2018, 664, 168–180. [Google Scholar] [CrossRef]
- Prokunina-Olsson, L. Genetics of the Human Interferon Lambda Region. J. Interferon Cytokine Res. 2019, 39, 599–608. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. ClinVar; [VCV000225949.4]. Available online: https://www.ncbi.nlm.nih.gov/clinvar/variation/VCV000225949.4 (accessed on 3 May 2023).
- Pinto, P.; Salgado, C.; Santos, N.P.C.; Santos, S.; Ribeiro-dos-Santos, Â. Influence of Genetic Ancestry on INDEL Markers of NFKβ1, CASP8, PAR1, IL4 and CYP19A1 Genes in Leprosy Patients. PLOS Neglected Trop. Dis. 2015, 9, e0004050. [Google Scholar] [CrossRef]
- Leal, D.F.d.V.B.; da Silva, M.N.S.; Fernandes, D.C.R.d.O.; Rodrigues, J.C.G.; Barros, M.C.d.C.; Pinto, P.D.d.o.C.; Pastana, L.F.; Silva, C.A.; Fernandes, M.R.; Assumpção, P.P.; et al. Amerindian genetic ancestry as a risk factor for tuberculosis in an amazonian population. PLoS ONE 2020, 15, e0236033. [Google Scholar]
- Carvalho, D.C.; Wanderley, A.V.; dos Santos, A.M.R.; Moreira, F.C.; de Sá, R.B.A.; Fernandes, M.R.; Modesto, A.A.; de Souza, T.P.; Cohen-Paes, A.; Leitão, L.P.; et al. Characterization of pharmacogenetic markers related to Acute Lymphoblastic Leukemia toxicity in Amazonian native Americans population. Sci. Rep. 2020, 10, 10292. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Hepatitis. Health Topics. Available online: https://www.who.int/health-topics/hepatitis#tab=tab_1 (accessed on 4 May 2023).
- Cuautle-Rodríguez, P.; Llerena, A.; Molina-Guarneros, J. Present status, and perspective of pharmacogenetics in Mexico. Drug Metab. Drug Interact. 2014, 29, 37–45. [Google Scholar]
- Nastri, A.C.S.S.; Malta, F.M.; Diniz, M.A.; Yoshino, A.; Abe-Sandes, K.; dos Santos, S.E.B.; de Castro Lyra, A.; Carrilho, F.J.; Pinho, J.R. Association of IFNL3 and IFNL4 polymorphisms with hepatitis C virus infection in a population from southeastern Brazil. Arch. Virol. 2016, 161, 1477–1484. [Google Scholar] [CrossRef]
- Ma, N.; Zhang, X.; Yang, L.; Zhou, J.; Liu, W.; Gao, X.; Yu, F.; Zheng, W.; Ding, S.; Gao, P.; et al. Role of Functional IFNL4, IFNLR1, IFNA, IFNAR2Polymorphisms in Hepatitis B virus-related liver disease in Han Chinese population. J. Viral Hepat. 2017, 25, 306–313. [Google Scholar] [CrossRef]
- Gonzalez-Aldaco, K.; Rebello Pinho, J.R.; Roman, S.; Gleyzer, K.; Fierro, N.A.; Oyakawa, L.; Ramos-Lopez, O.; Ferraz Santana, R.A.; Sitnik, R.; Panduro, A.; et al. Association with Spontaneous Hepatitis C Viral Clearance and Genetic Differentiation of IL28B/IFNL4 Haplotypes in Populations from Mexico. PLoS ONE 2016, 11, e0146258. [Google Scholar]
- Jefferies, M.; Rauff, B.; Rashid, H.; Lam, T.; Rafiq, S. Update on global epidemiology of viral hepatitis and preventive strategies. World J. Clin. Cases 2018, 6, 589–599. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6232563/ (accessed on 4 April 2023).
- Razavi, H. Global Epidemiology of Viral Hepatitis. Gastroenterol. Clin. N. Am. 2020, 49, 179–189. [Google Scholar]
- Sant’Anna, T.B.; Araujo, N.M. Hepatitis B Virus Genotype D: An Overview of Molecular Epidemiology, Evolutionary History, and Clinical Characteristics. Microorganisms 2023, 11, 1101. [Google Scholar]
- Pedergnana, V.; Irving, W.L.; Barnes, E.; McLauchlan, J.; Spencer, C.C.A. Impact of IFNL4 Genetic Variants on Sustained Virologic Response and Viremia in Hepatitis C Virus Genotype 3 Patients. J. Interferon Cytokine Res. 2019, 39, 642–649. [Google Scholar] [CrossRef]
- Galmozzi, E.; Facchetti, F.; Grossi, G.; Loglio, A.; Viganò, M.; Lunghi, G.; Colombo, M.; Lampertico, P. IFNL4 rs368234815 and rs117648444 variants predict off-treatment HBsAg seroclearance in IFN-treated HBeAg-negative chronic hepatitis B patients. Liver Int. 2017, 38, 417–423. [Google Scholar] [CrossRef] [PubMed]
- De Re, V.; Tornesello, M.L.; De Zorzi, M.; Caggiari, L.; Pezzuto, F.; Leone, P.; Racanelli, V.; Lauletta, G.; Zanussi, S.; Repetto, O.; et al. PDCD1 and IFNL4 genetic variants and risk of developing hepatitis C virus-related diseases. Liver Int. 2020, 41, 133–149. [Google Scholar]
- Luo, Y.; Jin, C.; Ling, Z.; Mou, X.; Zhang, Q.; Xiang, C. Association study of IL28B: rs12979860 and rs8099917 polymorphisms with SVR in patients infected with chronic HCV genotype 1 to PEG-INF/RBV therapy using systematic meta-analysis. Gene 2013, 513, 292–296. [Google Scholar] [CrossRef]
- Ragheb, M.M.; Nemr, N.A.; Kishk, R.M.; Mandour, M.F.; Abdou, M.M.; Matsuura, K.; Watanabe, T.; Tanaka, Y. Strong prediction of virological response to combination therapy by IL28B gene variants rs12979860 and rs8099917 in chronic hepatitis C genotype 4. Liver Int. 2014, 34, 890–895. [Google Scholar] [CrossRef]
- Prokunina-Olsson, L.; Muchmore, B.; Tang, W.; Pfeiffer, R.M.; Park, H.; Dickensheets, H.; Hergott, D.; Porter-Gill, P.; Mumy, A.; Kohaar, I.; et al. A variant upstream of IFNL3 (IL28B) creating a new interferon gene IFNL4 is associated with impaired clearance of hepatitis C virus. Nat. Genet. 2013, 45, 164–171. [Google Scholar] [CrossRef]
- Cohen-Paes, A.d.N.; de Carvalho, D.C.; Pastana, L.F.; Dobbin, E.A.F.; Moreira, F.C.; de Souza, T.P.; Fernandes, M.R.; Leal, D.F.; de Sá, R.B.; de Alcântara, A.L.; et al. Characterization of PCLO Gene in Amazonian Native American Populations. Genes 2022, 13, 499. [Google Scholar] [CrossRef]
- Garcia, U. Crônicas de caça e Criação; Hedra: São Paulo, Brazil, 2022. [Google Scholar]
- Sambrook, J.; Green, M.R. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Faust, G.G.; Hall, I.M. SAMBLASTER: Fast duplicate marking and structural variant read extraction. Bioinformatics 2014, 30, 2503–2505. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, A.; Vilella, A.J.; Cuppen, E.; Nijman, I.J.; Prins, P. Sambamba: Fast processing of NGS alignment formats. Bioinformatics 2015, 31, 2032–2034. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Machiela, M.J.; Chanock, S.J. LDlink: A web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants: Fig. 1. Bioinformatics 2015, 31, 3555–3557. [Google Scholar] [CrossRef]
- Salzano, F.M. Genetic diversity of South American human populations at the DNA and protein levels. J. Exp. Zool. 1998, 282, 157–163. [Google Scholar] [CrossRef]
- Rodrigues, J.C.G.; Fernandes, M.R.; Guerreiro, J.F.; da Silva, A.L.D.C.; Ribeiro-Dos-Santos, Â.; Santos, S.; Santos, N.P. Polymorphisms of ADME-related genes and their implications for drug safety and efficacy in Amazonian Amerindians. Sci. Rep. 2019, 9, 7201. [Google Scholar] [CrossRef]
- Rodrigues, J.C.G.; de Souza, T.P.; Pastana, L.F.; dos Santos, A.M.R.; Fernandes, M.R.; Pinto, P.; Wanderley, A.V.; de Souza, S.J.; Kroll, J.E.; Pereira, A.L.; et al. Identification of NUDT15 gene variants in Amazonian Amerindians and admixed individuals from northern Brazil. PLoS ONE 2020, 15, e0231651. [Google Scholar] [CrossRef]
- Dobbin, E.A.F.; Medeiros, J.A.G.; Costa, M.S.C.R.; Rodrigues, J.C.G.; Guerreiro, J.F.; Kroll, J.E.; Souza, S.J.; de Assumpção, P.P.; Ribeiro-dos-Santos, Â.; Santos, S.E.; et al. Identification of Variants (rs11571707, rs144848, and rs11571769) in the BRCA2 Gene Associated with Hereditary Breast Cancer in Indigenous Populations of the Brazilian Amazon. Genes 2021, 12, 142. [Google Scholar] [CrossRef] [PubMed]
- Vergara, C.; Thio, C.L.; Johnson, E.; Kral, A.H.; O’brien, T.R.; Goedert, J.J.; Mangia, A.; Piazzolla, V.; Mehta, S.H.; Kirk, G.D.; et al. Multi-Ancestry Genome-Wide Association Study of Spontaneous Clearance of Hepatitis C Virus. Gastroenterology 2019, 156, 1496–1507. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.P.; Ribeiro-Rodrigues, E.M.; Ribeiro-Dos-Santos, K.; Pereira, R.; Gusmão, L.; Amorim, A.; Guerreiro, J.F.; Zago, M.A.; Matte, C.; Hutz, M.H.; et al. Assessing individual interethnic admixture and population substructure using a 48-insertion-deletion (INSEL) ancestry-informative marker (AIM) panel. Hum. Mutat. 2010, 31, 184–190. [Google Scholar] [CrossRef]
- Key, F.M.; Peter, B.; Dennis, M.Y.; Huerta-Sánchez, E.; Tang, W.; Prokunina-Olsson, L.; Nielsen, R.; Andrés, A.M. Selection on a Variant Associated with Improved Viral Clearance Drives Local, Adaptive Pseudogenization of Interferon Lambda 4 (IFNL4). PLoS Genet. 2014, 10, e1004681. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lewis, C.M.; Jakobsson, M.; Ramachandran, S.; Ray, N.; Bedoya, G.; Rojas, W.; Parra, M.V.; A Molina, J.; Gallo, C.; et al. Genetic Variation and Population Structure in Native Americans. PLoS Genet. 2007, 3, e185. [Google Scholar] [PubMed]
- Amador, M.A.; Cavalcante, G.C.; Santos, N.P.; Gusmão, L.; Guerreiro, J.F.; Ribeiro-dos-Santos, Â.; Santos, S. Distribution of allelic and genotypic frequencies of IL1A, IL4, NFKB1 and PAR1 variants in Native American, African, European and Brazilian populations. BMC Res. Notes 2016, 9, 101. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.R.; Suppiah, V.; O’Connor, K.; Berg, T.; Weltman, M.; Abate, M.L.; Spengler, U.; Bassendine, M.; Matthews, G.; Irving, W.L.; et al. Identification of improved IL28B SNPs and haplotypes for prediction of drug response in treatment of hepatitis C using massively parallel sequencing in a cross-sectional European cohort. Genome Med. 2011, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Booth, D.R.; Ahlenstiel, G.; George, J. Pharmacogenomics of hepatitis C infections: Personalizing therapy. Genome Med. 2012, 4, 99. [Google Scholar] [CrossRef]
- Skoglund, P.; Reich, D. A genomic view of the peopling of the Americas. Curr. Opin. Genet. Dev. 2016, 41, 27–35. [Google Scholar] [CrossRef]


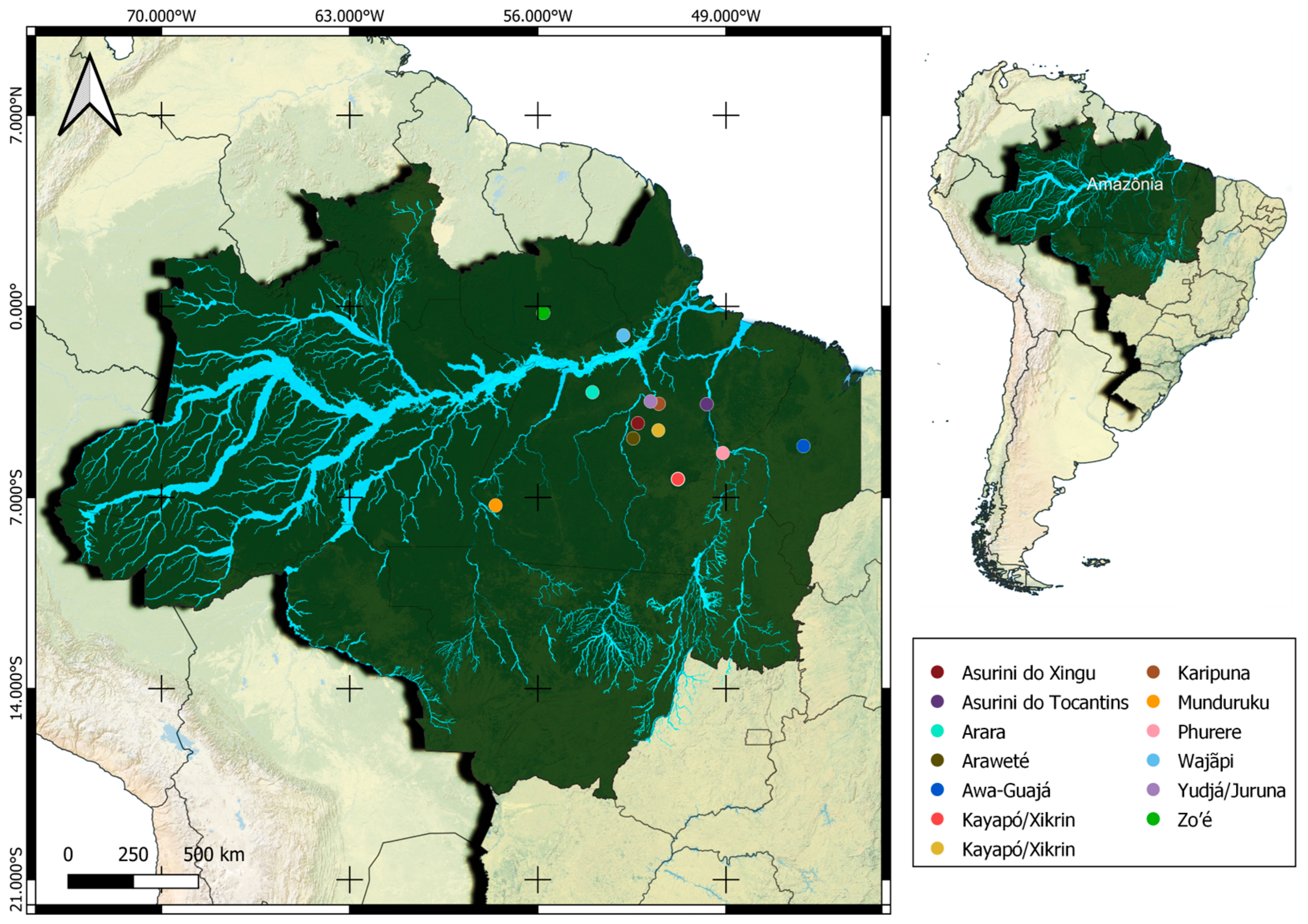{kind=link}
| Tribe | Location | State | Population (N) | Source |
|---|---|---|---|---|
| Asurini do Xingu | Altamira, Senador José Porfírio | Pará | 182 | SIASI |
| Arara/Arara do Iriri | Altamira, Brasil novo, Medicilância, Uruará | Pará | 298 | SIASI |
| Araweté | Altamira, São Félix do Xingu e Senador José Porfírio | Pará | 467 | SIASI |
| Asurini do Tocantins | Baião e Tucuruí | Pará | 565 | SIASI |
| Awa-Guajá | Arariboia | Maranhão | 520 | [29] |
| Kayapó/Xikrin | IL Bacajá (Altamira, Anapu, São Félix do Xingu, Senador José Porfírio | Pará | 746 | FUNAI |
| IL Xikrin do Cateté (Água Azu do Norte, Marabá, Parauapebas) | Pará | 1183 | SIASI | |
| Zo’é | Óbidos | Pará | 330 | IEPE |
| Wajãpi | Laranjal do Jari, Mazagão, Pedra Branca do Amapari | Pará | 1454 | DSEI/AP |
| Karipuna | TI Galibe (Oiapoque) | Amapá | 151 | SIASI |
| TI Juminá (Oiapoque) | Amapá | 291 | IEPE | |
| Uaçá I e II (Oiapoque) | Amapá | 4462 | FUNAI | |
| Phurere | Marabá | Pará | ||
| Munduruku | Itaituba e Jacareacanga | Pará | 6518 | SIASI |
| Yudjá/Juruna | Vitória do Xingu | Pará | 95 | SIASI |
| Genomic Position | SNP Identifier | Nucleotide | Variant | Consequence |
|---|---|---|---|---|
| 19:39246873 | rs12979731 | T > C | SNV | 3′UTR variant |
| 19:39246936 | rs370209610 | T > C | SNV | 3′UTR variant |
| 19:39246970 | rs11882871 | G > A | SNV | 3′UTR variant |
| 19:39247226 | rs12971396 | C > G | SNV | 3′UTR variant |
| 19:39247247 | rs137902769 | A > T | SNV | 3′UTR variant |
| 19:39247389 | rs73555604 | C > T | SNV | missense variant |
| 19:39247677 | rs111531283 | C > A | SNV | intron variant |
| 19:39247938 | rs117648444 | G > A | SNV | missense variant |
| 19:39248147 | rs12979860 | T > C | SNV | intron variant |
| 19:39248489 | rs4803221 | C > G | SNV | missense variant |
| 19:39248514 | rs11322783 | T > TT | INDEL | frameshift variant |
| 19:39248515 | rs74597329 | G > T | SNV | missense variant |
| 19:39248713 | rs4803222 | G > C | SNV | 5′UTR variant |
| 19:39737578 | rs570739705 | C > G | SNV | 3′UTR variant |
| SNP ID | Alleles | AFR | AMR | EAS | EUR | SAS | ABraOm | MISC | NAT |
|---|---|---|---|---|---|---|---|---|---|
| rs74597329 * | T | 0.364 | 0.621 | 0.932 | 0.683 | 0.766 | 0.624 | 0.744 | 0.838 |
| G | 0.636 | 0.379 | 0.068 | 0.317 | 0.234 | 0.376 | 0.256 | 0.162 | |
| rs4803221 * | C | 0.774 | 0.721 | 0.936 | 0.797 | 0.845 | 0.78 | 0.832 | 0.937 |
| G | 0.226 | 0.279 | 0.064 | 0.203 | 0.155 | 0.22 | 0.169 | 0.064 | |
| rs73555604 | C | 0.801 | 0.977 | 0.999 | 0.98 | 0.984 | 0.952 | ||
| T | 0.199 | 0.023 | 0.001 | 0.02 | 0.016 | 0.048 | |||
| rs117648444 | G | 0.932 | 0.937 | 0.995 | 0.906 | 0.95 | 0.921 | ||
| A | 0.068 | 0.063 | 0.005 | 0.094 | 0.05 | 0.079 | |||
| rs11322783 * | TT | 0.364 | 0.621 | 0.932 | 0.683 | 0.766 | 0.624 | 0.744 | 0.838 |
| T- | 0.636 | 0.379 | 0.068 | 0.317 | 0.234 | 0.376 | 0.256 | 0.162 |
| SNP ID | Alleles | AFR | AMR | EAS | EUR | SAS | ABraOm | MISC | NAT |
|---|---|---|---|---|---|---|---|---|---|
| rs74597329 * | T | 0.293 | 0.597 | 0.92 | 0.688 | 0.758 | 0.624 | 0.744 | 0.838 |
| G | 0.707 | 0.403 | 0.08 | 0.312 | 0. 242 | 0.376 | 0.256 | 0.162 | |
| rs4803221 * | C | 0.837 | 0.7 | 0.925 | 0.828 | 0.834 | 0.78 | 0.832 | 0.937 |
| G | 0.163 | 0.3 | 0.075 | 0.172 | 0.166 | 0.22 | 0.169 | 0.064 | |
| rs73555604 | C | 0.737 | 0.976 | 1 | 0.983 | 0.979 | 0.952 | ||
| T | 0.263 | 0.024 | 0.017 | 0.021 | 0.048 | ||||
| rs117648444 | G | 0.925 | 0.935 | 0.995 | 0.882 | 0.956 | 0.921 | ||
| A | 0.075 | 0.065 | 0.005 | 0.118 | 0.044 | 0.079 | |||
| rs11322783 * | T- | 0.707 | 0.403 | 0.081 | 0.311 | 0.239 | 0.376 | 0.256 | 0.162 |
| TT | 0.293 | 0.597 | 0.919 | 0.689 | 0.761 | 0.624 | 0.744 | 0.838 |
| rs74597329 | rs4803221 | rs11322783 | |
|---|---|---|---|
| AFR vs. MISC | <0.0001 | 0.4549 | <0.0001 |
| AMR vs. MISC | 0.1395 | 0.1395 | 0.1395 |
| EAS vs. MISC | 0.0031 | 0.0585 | 0.0031 |
| EUR vs. MISC | 0.4976 | 0.7734 | 0.4976 |
| SAS vs. MISC | 0.7734 | 1 | 0.7734 |
| ABraOm vs. MISC | 0.1395 | 0.5294 | 0.1395 |
| MISC vs. NAT | 0.2281 | 0.1395 | 0.2281 |
| AFR vs. NAT | <0.0001 | 0.0164 | <0.0001 |
| AMR vs. NAT | 0.0097 | 0.0031 | 0.0097 |
| EAS vs. NAT | 0.1659 | 1 | 0.1659 |
| EUR vs. NAT | 0.0638 | 0.0585 | 0.0638 |
| SAS vs. NAT | 0.4131 | 0.1395 | 0.4131 |
| ABraOm vs. NAT | 0.0097 | 0.025 | 0.0097 |
| rs74597329 | rs4803221 | rs11322783 | |
|---|---|---|---|
| AFR vs. MISC | <0.0001 | 0.4579 | <0.0001 |
| AMR vs. MISC | 0.1417 | 0.1417 | 0.1417 |
| EAS vs. MISC | 0.0003 | 0.057 | 0.0003 |
| EUR vs. MISC | 0.4999 | 0.7745 | 0.4999 |
| SAS vs. MISC | 0.7745 | 1 | 0.7745 |
| ABraOm vs. MISC | 0.1417 | 0.5313 | 0.1417 |
| MISC vs. NAT | 0.2303 | 0.1417 | 0.2303 |
| AFR vs. NAT | <0.0001 | 0.016 | <0.0001 |
| AMR vs. NAT | 0.0095 | 0.003 | 0.0095 |
| EAS vs. NAT | 0.1679 | 1 | 0.1679 |
| EUR vs. NAT | 0.0622 | 0.057 | 0.0622 |
| SAS vs. NAT | 0.4162 | 0.1417 | 0.4162 |
| ABraOm vs. NAT | 0.0095 | 0.0244 | 0.0095 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelim, C.C.; Martins, L.D.; Andrade, Á.A.F.; Moreira, F.C.; Guerreiro, J.F.; de Assumpção, P.P.; Santos, S.E.B.d.; Costa, G.d.L.C. Variants of IFNL4 Gene in Amazonian and Northern Brazilian Populations. Genes 2023, 14, 2075. https://doi.org/10.3390/genes14112075
Angelim CC, Martins LD, Andrade ÁAF, Moreira FC, Guerreiro JF, de Assumpção PP, Santos SEBd, Costa GdLC. Variants of IFNL4 Gene in Amazonian and Northern Brazilian Populations. Genes. 2023; 14(11):2075. https://doi.org/10.3390/genes14112075
Chicago/Turabian StyleAngelim, Carolina Cabral, Letícia Dias Martins, Álesson Adam Fonseca Andrade, Fabiano Cordeiro Moreira, João Farias Guerreiro, Paulo Pimentel de Assumpção, Sidney Emanuel Batista dos Santos, and Greice de Lemos Cardoso Costa. 2023. "Variants of IFNL4 Gene in Amazonian and Northern Brazilian Populations" Genes 14, no. 11: 2075. https://doi.org/10.3390/genes14112075