
Association between DNA Methylation in the Core Promoter Region of the CUT-like Homeobox 1 (CUX1) Gene and Lambskin Pattern in Hu Sheep
 , ,
, ,  , and
, and 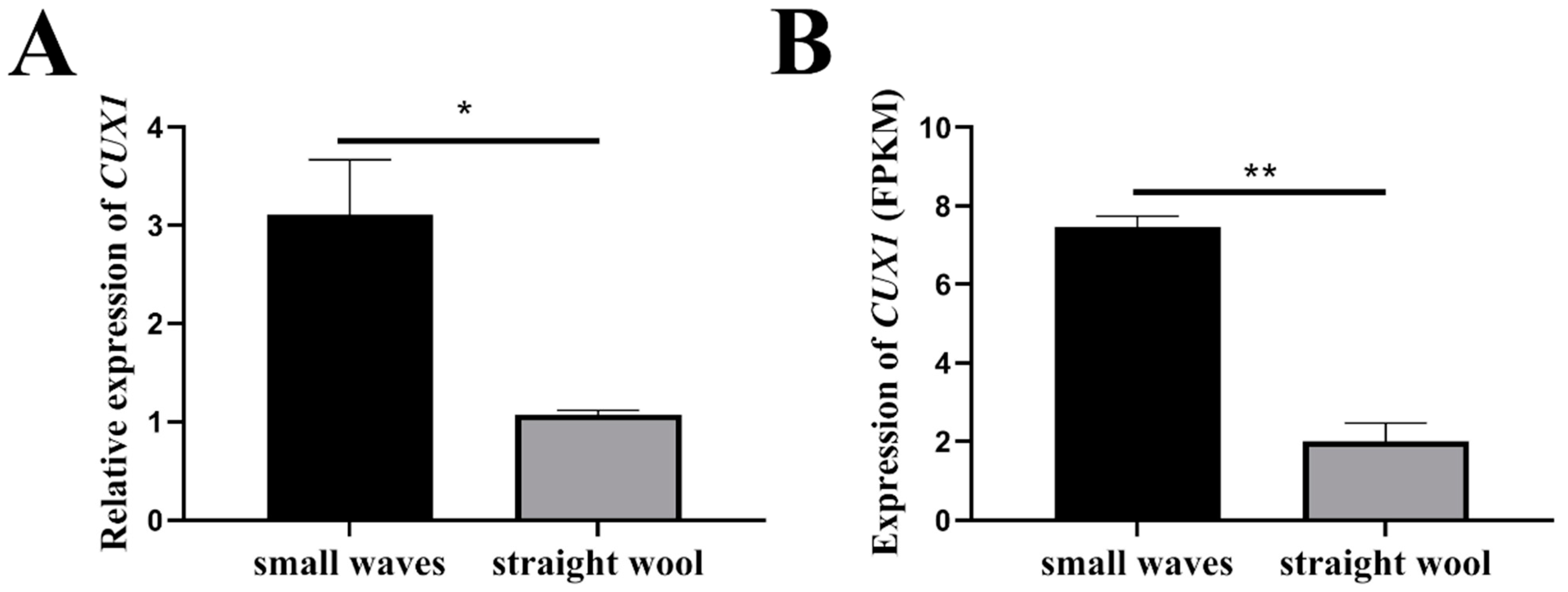{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Sample Collection
2.2. Quantitative Real-Time PCR
2.3. Cell Culture
2.4. Determination of CUX1 Core Promoter Region
2.5. DNA Methylation Detection in CUX1 Core Promoter Region
2.6. Statistical Analyses
3. Results
3.1. CUX1 Was Identified as a Potential Candidate Gene Regulating Lambskin Pattern of Hu Sheep
3.2. Determination of CUX1 Core Promoter Region
3.3. DNA Methylation Detection in CUX1 Core Promoter Region
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, W.; Ni, R.; Yin, J.F.; Musa, H.H.; Ding, T.J.; Chen, L. Genome Array of Hair Follicle Genes in Lambskin with Different Patterns. PLoS ONE 2013, 8, e68840. [Google Scholar] [CrossRef]
- Lv, X.Y.; Chen, W.H.; Sun, W.; Hussain, Z.; Wang, S.H.; Wang, J.Y. Analysis of lncRNAs Expression Profiles in Hair Follicle of Hu Sheep Lambskin. Animals 2020, 10, 1035. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Hu, T.Y.; He, M.L.; Gu, Y.F.; Cao, X.K.; Yuan, Z.H.; Lv, X.Y.; Getachew, T.; Quan, K.; Sun, W. Defining ovine dermal papilla cell markers and identifying key signaling pathways regulating its intrinsic properties. Front. Vet. Sci. 2023, 10, 1127501. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Ullrich, R.; Paus, R. Molecular principles of hair follicle induction and morphogenesis. Bioessays 2005, 27, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Carmon, K.S.; Gong, X.; Lin, Q.S.; Thomas, A.; Liu, Q.Y. R-spondins function as ligands of the orphan receptors LGR4 and LGR5 to regulate Wnt/β-catenin signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 11452–11457. [Google Scholar] [CrossRef]
- Ito, M.; Nguyen, J.; Myung, P.; Garza, L.; Andl, T.; Millar, S.; Cotsarelis, G. Wnt modulates the differentiation status of interfollicular epidermal melanocytes arising from melanocyte stem cells in the hair follicle bulge after wounding or UVB irradiation. J. Invest. Dermatol. 2008, 128, S156. [Google Scholar]
- Aubin-Houzelstein, G. Notch Signaling and the Developing Hair Follicle. Notch Signal. Embryol. Cancer 2012, 727, 142–160. [Google Scholar] [CrossRef]
- Botchkarev, V.A.; Sharov, A.A. BMP signaling in the control of skin development and hair follicle growth. Differentiation 2004, 72, 512–526. [Google Scholar] [CrossRef]
- Jing, J.; Xu, P.; Xu, J.L.; Ding, Y.X.; Yang, X.S.; Jin, X.Q.; Zhou, L.J.; Chen, Y.H.; Wu, X.J.; Lu, Z.F. Expression and localization of Sox10 during hair follicle morphogenesis and induced hair cycle. Int. J. Med. Sci. 2021, 18, 3498–3505. [Google Scholar] [CrossRef]
- Sriwiriyanont, P.; Lynch, K.A.; Maier, E.A.; Hahn, J.M.; Supp, D.M.; Boyce, S.T. Morphogenesis of chimeric hair follicles in engineered skin substitutes with human keratinocytes and murine dermal papilla cells. Exp. Dermatol. 2012, 21, 783–785. [Google Scholar] [CrossRef]
- Li, Y.; Lv, X.Y.; Wang, S.H.; Cao, X.K.; Yuan, Z.H.; Getachew, T.; Mwacharo, J.M.; Haile, A.; Sun, W. BMP7 Functions to Regulate Proliferation of Dermal Papilla Cells in Hu Sheep. Genes 2022, 13, 201. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Sun, Q.L.; Wan, L.; Wang, X.; Feng, Y.; Luo, J.D.; Wu, H.L. CUX1, A Controversial Player in Tumor Development. Front. Oncol. 2020, 10, 738. [Google Scholar] [CrossRef] [PubMed]
- Topka, S.; Glassmann, A.; Weisheit, G.; Schuller, U.; Schilling, K. The Transcription Factor Cux1 in Cerebellar Granule Cell Development and Medulloblastoma Pathogenesis. Cerebellum 2014, 13, 698–712. [Google Scholar] [CrossRef]
- Ramdzan, Z.M.; Ginjala, V.; Pinder, J.B.; Chung, D.; Donovan, C.M.; Kaur, S.; Leduy, L.; Dellaire, G.; Ganesan, S.; Nepveu, A. The DNA repair function of CUX1 contributes to radioresistance. Oncotarget 2017, 8, 19021–19038. [Google Scholar] [CrossRef] [PubMed]
- Krug, S.; Weissbach, J.; Blank, A.; Perren, A.; Haybaeck, J.; Fendrich, V.; Rinke, A.; Gress, T.M.; Rosendahl, J.; Michl, P. CUX1-Transcriptional Master Regulator of Tumor Progression in Pancreatic Neuroendocrine Tumors. Cancers 2020, 12, 1957. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.R.; Soucie, E.; Moon, N.S.; Martin-Soudant, N.; Berube, G.; Leduy, L.; Nepveu, A. Exon/intron structure and alternative transcripts of the CUTL1 gene. Gene 2000, 241, 75–85. [Google Scholar] [CrossRef]
- Tufarelli, C.; Fujiwara, Y.; Zappulla, D.C.; Neufeld, E.J. Hair defects and pup loss in mice with targeted deletion of the first cut repeat domain of the Cux/CDP homeoprotein gene. Dev. Biol. 1998, 200, 69–81. [Google Scholar] [CrossRef]
- Feng, F.; Zhao, Z.Q.; Zhou, Y.F.; Cheng, Y.H.; Wu, X.J.; Heng, X.Y. CUX1 Facilitates the Development of Oncogenic Properties Via Activating Wnt/β-Catenin Signaling Pathway in Glioma. Front. Mol. Biosci. 2021, 8, 705008. [Google Scholar] [CrossRef]
- Katoh, M.; Katoh, M. Transcriptional mechanisms of WNT5A based on NF-kappa B, Hedgehog, TGF β, and Notch signaling cascades. Int. J. Mol. Med. 2009, 23, 763–769. [Google Scholar] [CrossRef]
- Zhou, H.; Huang, S.A.; Lv, X.Y.; Wang, S.H.; Cao, X.K.; Yuan, Z.H.; Getachew, T.; Mwacharo, J.M.; Haile, A.; Quan, K.; et al. Effect of CUX1 on the Proliferation of Hu Sheep Dermal Papilla Cells and on the Wnt/β-Catenin Signaling Pathway. Genes 2023, 14, 423. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Sennett, R.; Rezza, A.; Clavel, C.; Grisanti, L.; Zemla, R.; Najam, S.; Rendl, M. Wnt/β-catenin signaling in dermal condensates is required for hair follicle formation. Dev. Biol. 2014, 385, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.Y.; Huang, S.A.; Lv, X.Y.; Wang, S.H.; Getachew, T.; Mwacharo, J.M.; Haile, A.; Sun, W. miR-143 Targeting CUX1 to Regulate Proliferation of Dermal Papilla Cells in Hu Sheep. Genes 2021, 12, 2017. [Google Scholar] [CrossRef]
- Takahashi, Y.; Valencia, M.M.; Yu, Y.; Ouchi, Y.; Takahashi, K.; Shokhirev, M.N.; Lande, K.; Williams, A.E.; Fresia, C.; Kurita, M.; et al. Transgenerational inheritance of acquired epigenetic signatures at CpG islands in mice. Cell 2023, 186, 715–731. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Barahona, V.; Joshi, R.S.; Esteller, M. Use of DNA methylation profiling in translational oncology. Semin. Cancer Biol. 2022, 83, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Papanicolau-Sengos, A.; Aldape, K. DNA Methylation Profiling: An Emerging Paradigm for Cancer Diagnosis. Annu. Rev. Pathol-Mech. 2022, 17, 295–321. [Google Scholar] [CrossRef]
- Kim, J.Y.; Tavare, S.; Shibata, D. Human hair genealogies and stem cell latency. BMC Biol. 2006, 4, 2. [Google Scholar] [CrossRef]
- Bai, L.Y.; Sun, H.T.; Jiang, W.X.; Yang, L.P.; Liu, G.Y.; Zhao, X.Y.; Hu, H.M.; Wang, J.Y.; Gao, S.X. DNA methylation and histone acetylation are involved in Wnt10b expression during the secondary hair follicle cycle in Angora rabbits. J. Anim. Physiol. Anim. Nutr. 2021, 105, 599–609. [Google Scholar] [CrossRef]
- Zhao, B.H.; Li, J.L.; Liu, M.; Yang, N.S.; Bao, Z.Y.; Zhang, X.Y.; Dai, Y.Y.; Cai, J.W.; Chen, Y.; Wu, X.S. DNA Methylation Mediates lncRNA2919 Regulation of Hair Follicle Regeneration. Int. J. Mol. Sci. 2022, 23, 9481. [Google Scholar] [CrossRef]
- Wang, S.H.; Li, F.; Liu, J.W.; Zhang, Y.L.; Zheng, Y.J.; Ge, W.; Qu, L.; Wang, X. Integrative Analysis of Methylome and Transcriptome Reveals the Regulatory Mechanisms of Hair Follicle Morphogenesis in Cashmere Goat. Cells 2020, 9, 969. [Google Scholar] [CrossRef]
- Briollais, L.; Rustand, D.; Allard, C.; Wu, Y.Y.; Xu, J.X.; Rajan, S.G.; Hivert, M.F.; Doyon, M.; Bouchard, L.; McGowan, P.O.; et al. DNA methylation mediates the association between breastfeeding and early-life growth trajectories. Clin. Epigenetics 2021, 13, 231. [Google Scholar] [CrossRef]
- Lv, X.Y.; Gao, W.; Jin, C.Y.; Wang, Y.; Chen, W.H.; Wang, L.H.; Zou, S.X.; Sheng, S.X.; Chen, L.; Sun, W. Divergently expressed RNA identification and interaction prediction of long non-coding RNA and mRNA involved in Hu sheep hair follicle. Sci. Rep. 2019, 9, 7283. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.L.; Wang, S.H.; Cao, X.K.; Yuan, Z.H.; Getachew, T.; Mwacharo, J.M.; Haile, A.; Lv, X.Y.; Sun, W. The Effect of EGR1 on the Proliferation of Dermal Papilla Cells. Genes 2022, 13, 1242. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhao, C.T.; Wang, Y.; Yuan, X.B. The Transcription Factor Cux1 Regulates Dendritic Morphology of Cortical Pyramidal Neurons. PLoS ONE 2010, 5, e10596. [Google Scholar] [CrossRef] [PubMed]
- Ramdzan, Z.M.; Nepveu, A. CUX1, a haploinsufficient tumour suppressor gene overexpressed in advanced cancers. Nat. Rev. Cancer 2014, 14, 673–682. [Google Scholar] [CrossRef]
- Sansregret, L.; Nepveu, A. The multiple roles of CUX1: Insights from mouse models and cell-based assays. Gene 2008, 412, 84–94. [Google Scholar] [CrossRef]
- Ong, M.L.; Tuan, T.A.; Poh, J.; Teh, A.L.; Chen, L.; Pan, H.; MacIsaac, J.L.; Kobor, M.S.; Chong, Y.S.; Kwek, K.; et al. Neonatal amygdalae and hippocampi are influenced by genotype and prenatal environment, and reflected in the neonatal DNA methylome. Genes. Brain Behav. 2019, 18, e12576. [Google Scholar] [CrossRef]
- Smyth, L.J.; McKay, G.J.; Maxwell, A.P.; McKnight, A.J. DNA hypermethylation and DNA hypomethylation is present at different loci in chronic kidney disease. Epigenetics 2014, 9, 366–376. [Google Scholar] [CrossRef]
- Pacht, E. Characterization of DNA Hydroxymethylation in the Cattle and Sheep Brain. Ph.D. Thesis, University of Vermont, Burlington, VT, USA, 2023. [Google Scholar]
- Lin, Q.L.; Ding, H.; Zheng, Z.; Gu, Z.Q.; Ma, J.H.; Chen, L.; Chan, P.; Cai, Y.N. Promoter methylation analysis of seven clock genes in Parkinson’s disease. Neurosci. Lett. 2012, 507, 147–150. [Google Scholar] [CrossRef]
- Wang, H.F.; Feng, H.Y.; Sun, J.; Zhou, Y.J.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Age-associated changes in DNA methylation and expression of the TNF α gene in pigs. Genes Genet. Syst. 2018, 93, 191–198. [Google Scholar] [CrossRef]
- Smith, J.; Sen, S.; Weeks, R.J.; Eccles, M.R.; Chatterjee, A. Promoter DNA Hypermethylation and Paradoxical Gene Activation. Trends Cancer 2020, 6, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Shimada, A.; Okumura, A.; Yarnasaki, S.; Iwata, Y.; Koizumi, N.; Nishihara, M.; Mishiba, K. A 64-bp sequence containing the GAAGA motif is essential for CaMV-35S promoter methylation in gentian. Bba-Gene Regul. Mech. 2017, 1860, 861–869. [Google Scholar] [CrossRef]
- Wu, Z.C.; Shi, D.F.; Jin, J.; Fan, H.R.; Bao, W.B.; Wu, S.L. DNA Methylation of Pig FUT3 Promoter Alters mRNA Expression to Regulate E. coli F18 Susceptibility. Genes 2021, 12, 1586. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.P.; Reid, A.M.A.; Wilson, P.W.; Dunn, I.C. Identification of the Core Promoter and Variants Regulating Chicken CCKAR Expression. Genes 2022, 13, 1083. [Google Scholar] [CrossRef] [PubMed]
- Muise-Helmericks, R.C.; Rosen, N. Identification of a novel repressive element in the proximal lck promoter. J. Biol. Chem. 1995, 270, 27538–27543. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shi, T.; Zhou, Y.; Liu, M.; Klaus, S.; Lan, X.Y.; Lei, C.Z.; Chen, H. A novel PAX7 10-bp indel variant modulates promoter activity, gene expression and contributes to different phenotypes of Chinese cattle. Sci. Rep. 2018, 8, 1724. [Google Scholar] [CrossRef]
- Rishi, V.; Bhattacharya, P.; Chatterjee, R.; Rozenberg, J.; Zhao, J.F.; Glass, K.; Fitzgerald, P.; Vinson, C. CpG methylation of half-CRE sequences creates C/EBP α binding sites that activate some tissue-specific genes. Proc. Natl. Acad. Sci. USA 2010, 107, 20311–20316. [Google Scholar] [CrossRef]
- Bui, C.; Barter, M.J.; Scott, J.L.; Xu, Y.B.; Galler, M.; Reynard, L.N.; Rowan, A.D.; Young, D.A. cAMP response element-binding (CREB) recruitment following a specific CpG demethylation leads to the elevated expression of the matrix metalloproteinase 13 in human articular chondrocytes and osteoarthritis. Faseb J. 2012, 26, 3000–3011. [Google Scholar] [CrossRef]
- Murayama, A.; Sakura, K.; Nakama, M.; Yasuzawa-Tanaka, K.; Fujita, E.; Tateishi, Y.; Wang, Y.A.; Ushijima, T.; Baba, T.; Shibuya, K.; et al. A specific CpG site demethylation in the human interleukin 2 gene promoter is an epigenetic memory. Embo J. 2006, 25, 1081–1092. [Google Scholar] [CrossRef]
- Hashimoto, K.; Otero, M.; Imagawa, K.; de Andres, M.C.; Coico, J.M.; Roach, H.I.; Oreffo, R.O.C.; Marcu, K.B.; Goldring, M.B. Regulated Transcription of Human Matrix Metalloproteinase 13 (MMP13) and Interleukin-1 β (IL1B) Genes in Chondrocytes Depends on Methylation of Specific Proximal Promoter CpG Sites. J. Biol. Chem. 2013, 288, 10061–10072. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, X.; Li, Y.; Chen, W.; Wang, S.; Cao, X.; Yuan, Z.; Getachew, T.; Mwacharo, J.; Haile, A.; Li, Y.; et al. Association between DNA Methylation in the Core Promoter Region of the CUT-like Homeobox 1 (CUX1) Gene and Lambskin Pattern in Hu Sheep. Genes 2023, 14, 1873. https://doi.org/10.3390/genes14101873
Lv X, Li Y, Chen W, Wang S, Cao X, Yuan Z, Getachew T, Mwacharo J, Haile A, Li Y, et al. Association between DNA Methylation in the Core Promoter Region of the CUT-like Homeobox 1 (CUX1) Gene and Lambskin Pattern in Hu Sheep. Genes. 2023; 14(10):1873. https://doi.org/10.3390/genes14101873
Chicago/Turabian StyleLv, Xiaoyang, Yue Li, Weihao Chen, Shanhe Wang, Xiukai Cao, Zehu Yuan, Tesfaye Getachew, Joram Mwacharo, Aynalem Haile, Yutao Li, and et al. 2023. "Association between DNA Methylation in the Core Promoter Region of the CUT-like Homeobox 1 (CUX1) Gene and Lambskin Pattern in Hu Sheep" Genes 14, no. 10: 1873. https://doi.org/10.3390/genes14101873