
Comparative Studies on Duplicated foxl2 Paralogs in Spotted Knifejaw Oplegnathus punctatus Show Functional Diversification

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Sample Collection
2.2. Identification of FOXL Family Genes in Spotted Knifejaw
2.3. Phylogenetic Analysis of FOXL Genes
2.4. Synteny Analyses of Vertebrate foxl2 and foxl2l
2.5. Protein Sequence Alignment and Motif Scan Analyses
2.6. RNA Extraction, cDNA Synthesis, and Tissue Distribution Analysis
2.7. Positive Selection Test for Duplicated foxl2 Genes
3. Results
3.1. Identification of Two foxl2 Paralogs in Spotted Knifejaw
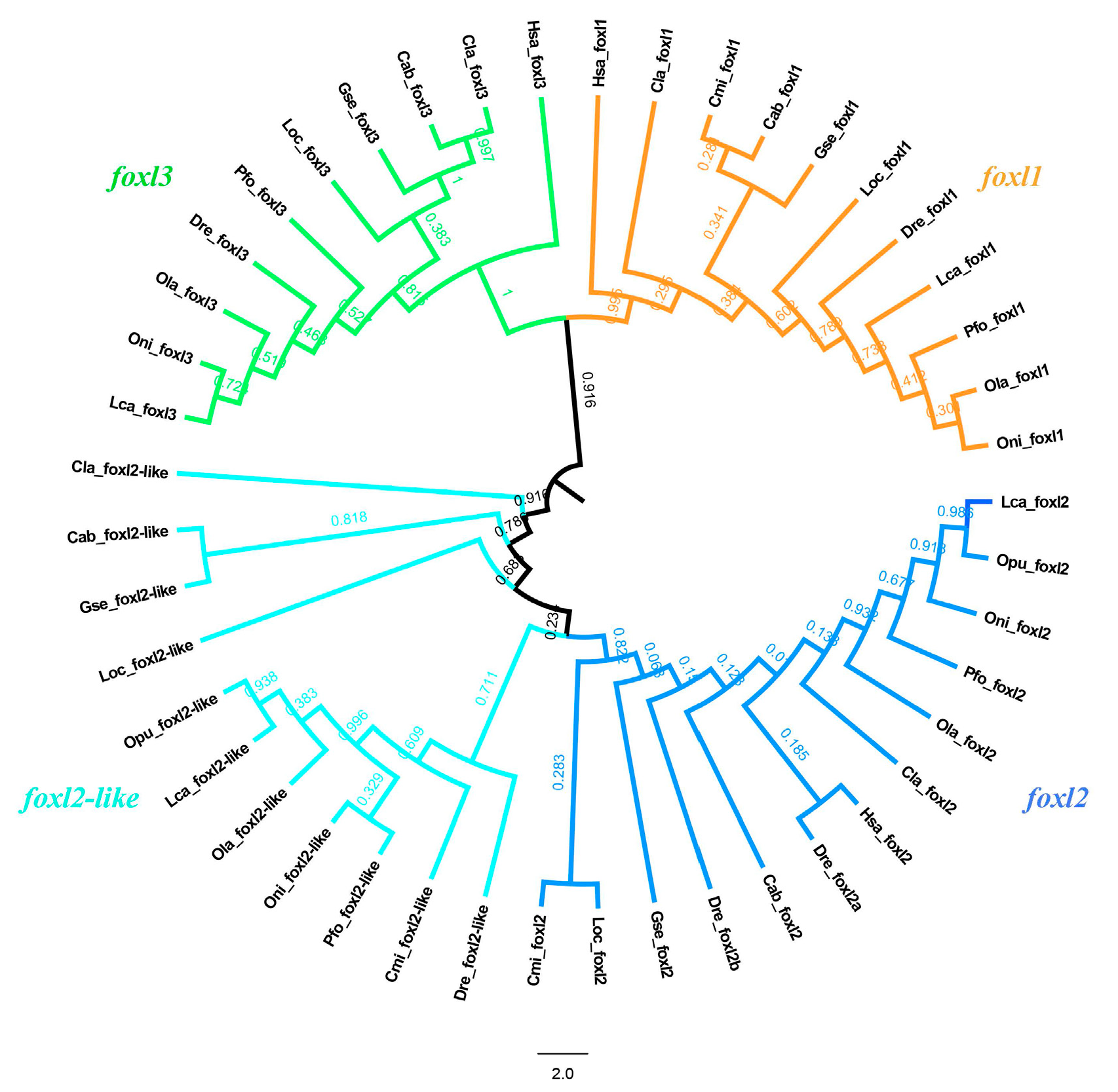
3.2. Phylogeny Analysis of FOXL Genes
3.3. Synteny of foxl2 and foxl2l Paralogs
3.4. Protein Sequences Alignment and Motifs in foxl2 and foxl2l
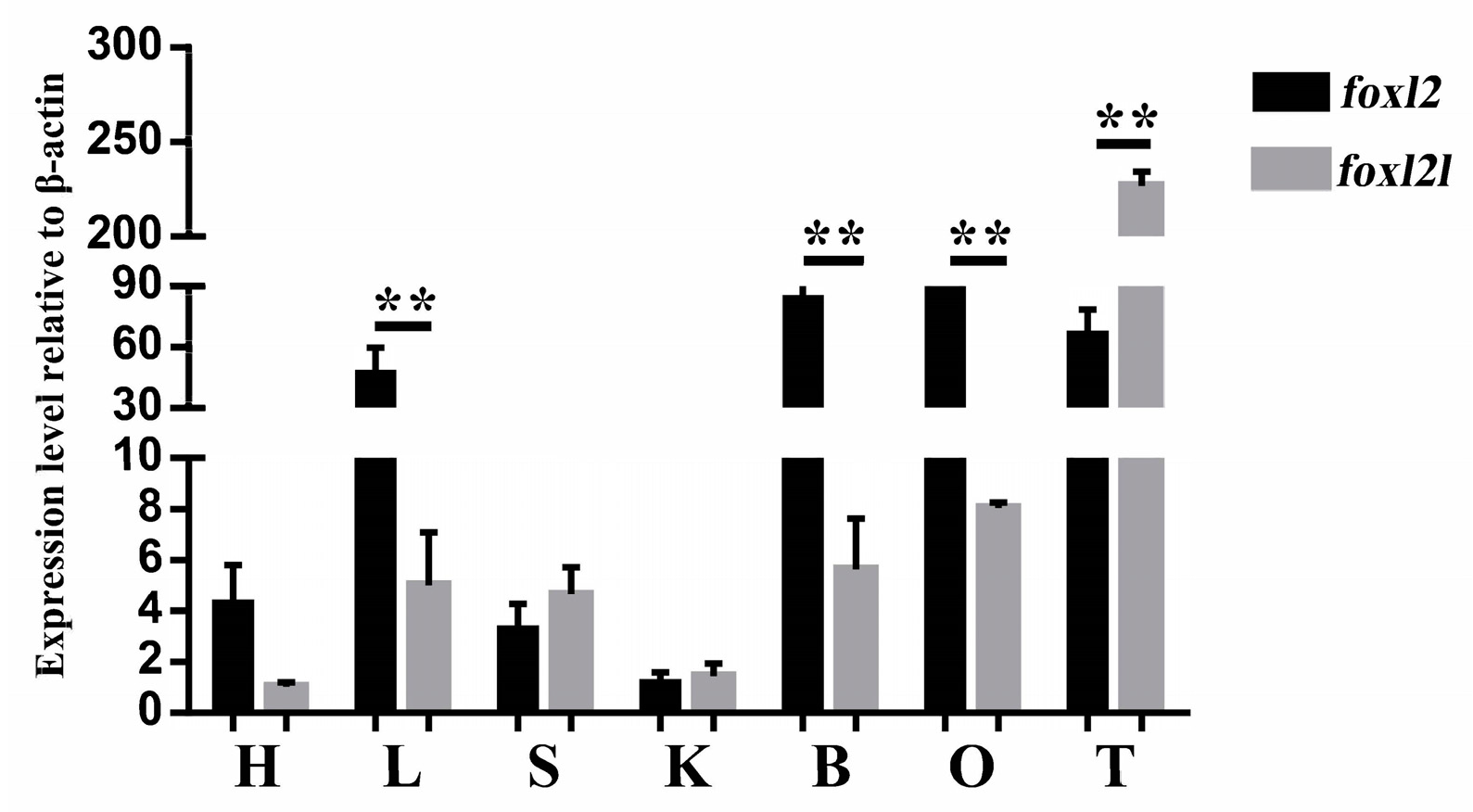
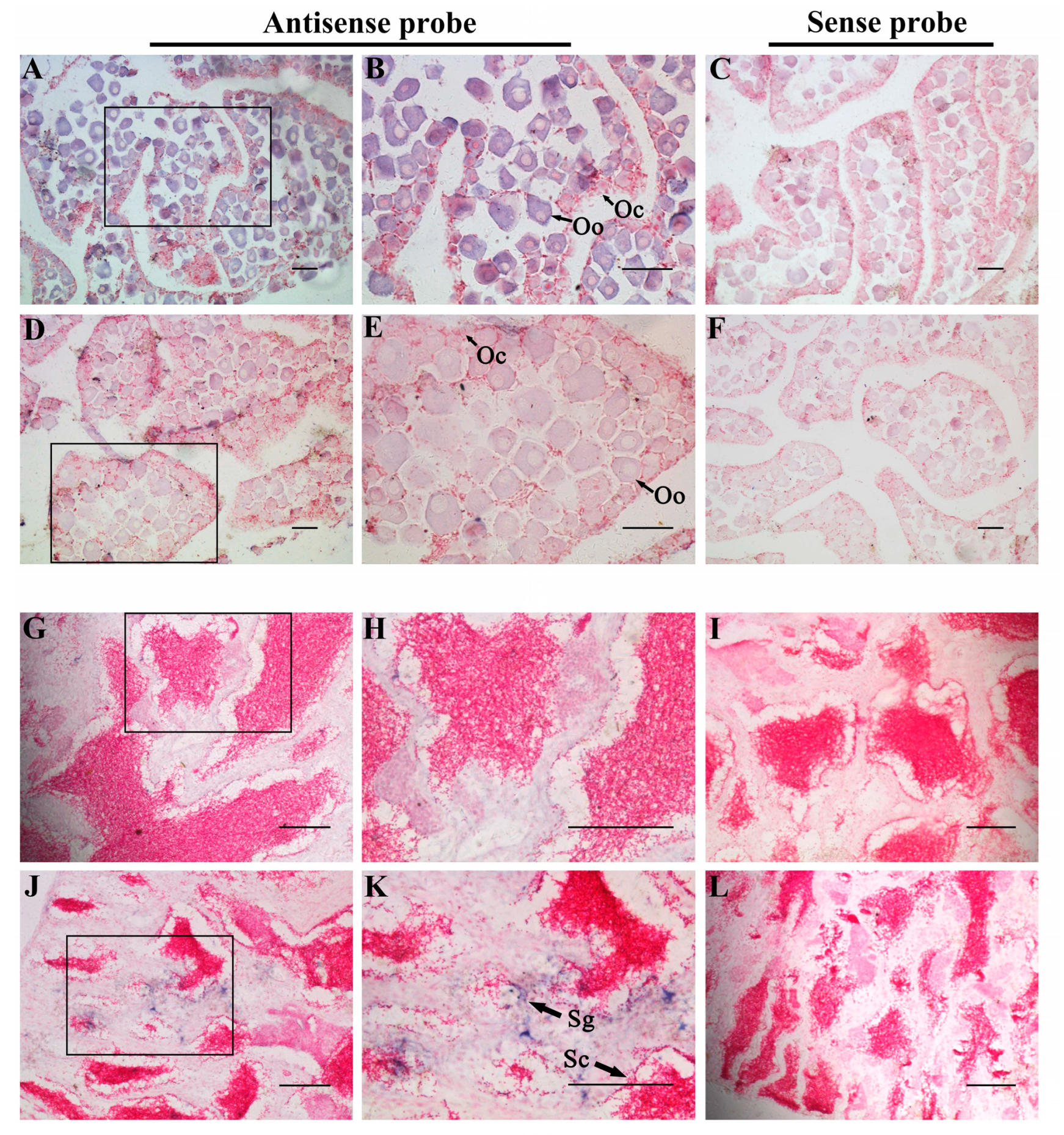
3.5. Tissue Distribution of Two foxl2 Paralogs in Spotted Knifejaw
3.6. Molecular Evolution of Teleost foxl2 and foxl2l
4. Discussion
4.1. Expansion of Fox Family Genes
4.2. foxl2 and foxl2l Are Generated from WGD at the Base of Vertebrate Radiation
4.3. Functional Diversification of Spotted Knifejaw foxl2 and foxl2l
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weigel, D.; Jürgens, G.; Küttner, F.; Seifert, E.; Jäckle, H. The homeotic gene fork head encodes a nuclear protein and is expressed in the terminal regions of the Drosophila embryo. Cell 1989, 57, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Gisselbrecht, S.S.; Rogers, J.M.; Hartl, D.L.; Bulyk, M.L. DNA-binding specificity changes in the evolution of forkhead transcription factors. Proc. Natl. Acad. Sci. USA 2013, 110, 12349–12354. [Google Scholar] [CrossRef] [PubMed]
- Mazet, F.; Yu, J.-K.; Liberles, D.A.; Holland, L.Z.; Shimeld, S.M. Phylogenetic relationships of the Fox (Forkhead) gene family in the Bilateria. Gene 2003, 316, 79–89. [Google Scholar] [CrossRef]
- Shen, X.; Cui, J.; Gong, Q. Fox gene loci in Takifugu rubripes and Tetraodon nigroviridis genomes and comparison with those of medaka and zebrafish genomes. Genome 2011, 54, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Hannenhalli, S.; Kaestner, K.H. The evolution of Fox genes and their role in development and disease. Nat. Rev. Genet. 2009, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, P.; Mahlapuu, M. Forkhead transcription factors: Key players in development and metabolism. Dev. Biol. 2002, 250, 1–23. [Google Scholar] [CrossRef]
- Labbé, E.; Silvestri, C.; Hoodless, P.A.; Wrana, J.L.; Attisano, L. Smad2 and Smad3 positively and negatively regulate TGFβ-dependent transcription through the forkhead DNA-binding protein FAST2. Mol. Cell 1998, 2, 109–120. [Google Scholar] [CrossRef]
- Kops, G.J.; Burgering, B.M. Forkhead transcription factors: New insights into protein kinase B (c-akt) signaling. J. Mol. Med. 1999, 77, 656–665. [Google Scholar] [CrossRef]
- Perreault, N.; Katz, J.P.; Sackett, S.D.; Kaestner, K.H. Foxl1 controls the Wnt/β-catenin pathway by modulating the expression of proteoglycans in the gut. J. Biol. Chem. 2001, 276, 43328–43333. [Google Scholar] [CrossRef]
- Baron, D.; Batista, F.; Chaffaux, S.; Cocquet, J.; Cotinot, C.; Cribiu, E.; De Baeree, E.; Guiguen, Y.; Jaubert, F.; Pailhoux, E. Foxl2 gene and the development of the ovary: A story about goat, mouse, fish and woman. Reprod. Nutr. Dev. 2005, 45, 377–382. [Google Scholar] [CrossRef]
- Bentsi-Barnes, I.K.; Kuo, F.-T.; Barlow, G.M.; Pisarska, M.D. Human forkhead L2 represses key genes in granulosa cell differentiation including aromatase, P450scc, and cyclin D2. Fertil. Steril. 2010, 94, 353–356. [Google Scholar] [CrossRef]
- Kuo, F.-T.; Bentsi-Barnes, I.K.; Barlow, G.M.; Pisarska, M.D. Mutant Forkhead L2 (FOXL2) proteins associated with premature ovarian failure (POF) dimerize with wild-type FOXL2, leading to altered regulation of genes associated with granulosa cell differentiation. Endocrinology 2011, 152, 3917–3929. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Suh, D.-S.; Lee, K.; Bae, J. Positive cross talk between FOXL2 and antimüllerian hormone regulates ovarian reserve. Fertil. Steril. 2014, 102, 847–855.e1. [Google Scholar] [CrossRef] [PubMed]
- Boulanger, L.; Pannetier, M.; Gall, L.; Allais-Bonnet, A.; Elzaiat, M.; Le Bourhis, D.; Daniel, N.; Richard, C.; Cotinot, C.; Ghyselinck, N.B. FOXL2 is a female sex-determining gene in the goat. Curr. Biol. 2014, 24, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Liu, L.; Wang, J.; Wei, H.; Li, Y.; Zhang, L.; Guo, Z.; Li, Y.; Liu, T.; Zeng, Q. Ancient homomorphy of molluscan sex chromosomes sustained by reversible sex-biased genes and sex determiner translocation. Nat. Ecol. Evol. 2022, 6, 1891–1906. [Google Scholar] [CrossRef]
- Uda, M.; Ottolenghi, C.; Crisponi, L.; Garcia, J.E.; Deiana, M.; Kimber, W.; Forabosco, A.; Cao, A.; Schlessinger, D.; Pilia, G. Foxl2 disruption causes mouse ovarian failure by pervasive blockage of follicle development. Hum. Mol. Genet. 2004, 13, 1171–1181. [Google Scholar] [CrossRef]
- Zhang, X.; Li, M.; Ma, H.; Liu, X.; Shi, H.; Li, M.; Wang, D. Mutation of foxl2 or cyp19a1a results in female to male sex reversal in XX Nile tilapia. Endocrinology 2017, 158, 2634–2647. [Google Scholar] [CrossRef]
- Govoroun, M.S.; Pannetier, M.; Pailhoux, E.; Cocquet, J.; Brillard, J.P.; Couty, I.; Batellier, F.; Cotinot, C. Isolation of chicken homolog of the FOXL2 gene and comparison of its expression patterns with those of aromatase during ovarian development. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2004, 231, 859–870. [Google Scholar]
- Rhen, T.; Metzger, K.; Schroeder, A.; Woodward, R. Expression of putative sex-determining genes during the thermosensitive period of gonad development in the snapping turtle, Chelydra serpentina. Sex. Dev. 2007, 1, 255–270. [Google Scholar] [CrossRef]
- Okada, E.; Yoshimoto, S.; Ikeda, N.; Kanda, H.; Tamura, K.; Shiba, T.; Takamatsu, N.; Ito, M. Xenopus W-linked DM-W induces Foxl2 and Cyp19 expression during ovary formation. Sex. Dev. 2009, 3, 38–42. [Google Scholar] [CrossRef]
- Crespo, B.; Lan-Chow-Wing, O.; Rocha, A.; Zanuy, S.; Gómez, A. foxl2 and foxl3 are two ancient paralogs that remain fully functional in teleosts. Gen. Comp. Endocrinol. 2013, 194, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Jia, D.; Hu, Q.; Li, D. Foxl3, a target of miR-9, stimulates spermatogenesis in spermatogonia during natural sex change in Monopterus albus. Endocrinology 2016, 157, 4388–4399. [Google Scholar] [CrossRef]
- Nishimura, T.; Sato, T.; Yamamoto, Y.; Watakabe, I.; Ohkawa, Y.; Suyama, M.; Kobayashi, S.; Tanaka, M. foxl3 is a germ cell-intrinsic factor involved in sperm-egg fate decision in medaka. Science 2015, 349, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-J.; Wang, Y.; Li, Z.; Zhou, L.; Gui, J.-F. Sequential, divergent, and cooperative requirements of foxl2a and foxl2b in ovary development and maintenance of zebrafish. Genetics 2017, 205, 1551–1572. [Google Scholar] [CrossRef] [PubMed]
- von Schalburg, K.R.; Yasuike, M.; Yazawa, R.; De Boer, J.G.; Reid, L.; So, S.; Robb, A.; Rondeau, E.B.; Phillips, R.B.; Davidson, W.S. Regulation and expression of sexual differentiation factors in embryonic and extragonadal tissues of Atlantic salmon. BMC Genom. 2011, 12, 31. [Google Scholar] [CrossRef]
- von Schalburg, K.R.; Yasuike, M.; Davidson, W.S.; Koop, B.F. Regulation, expression and characterization of aromatase (cyp19b1) transcripts in ovary and testis of rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2010, 155, 118–125. [Google Scholar] [CrossRef]
- Kikuchi, M.; Nishimura, T.; Saito, D.; Shigenobu, S.; Takada, R.; Gutierrez-Triana, J.A.; Cerdán, J.L.M.; Takada, S.; Wittbrodt, J.; Suyama, M. Novel components of germline sex determination acting downstream of foxl3 in medaka. Dev. Biol. 2019, 445, 80–89. [Google Scholar] [CrossRef]
- Dehal, P.; Boore, J.L. Two rounds of whole genome duplication in the ancestral vertebrate. PLoS Biol. 2005, 3, e314. [Google Scholar] [CrossRef]
- Hoegg, S.; Meyer, A. Hox clusters as models for vertebrate genome evolution. Trends Genet. 2005, 21, 421–424. [Google Scholar] [CrossRef]
- Force, A.; Lynch, M.; Pickett, F.B.; Amores, A.; Yan, Y.-l.; Postlethwait, J. Preservation of duplicate genes by complementary, degenerative mutations. Genetics 1999, 151, 1531–1545. [Google Scholar] [CrossRef]
- Cocquet, J.; De Baere, E.; Gareil, M.; Pannetier, M.; Xia, X.; Fellous, M.; Veitia, R. Structure, evolution and expression of the FOXL2 transcription unit. Cytogenet. Genome Res. 2003, 101, 206–211. [Google Scholar] [CrossRef]
- Du, X.; Wang, B.; Liu, X.; Liu, X.; He, Y.; Zhang, Q.; Wang, X. Comparative transcriptome analysis of ovary and testis reveals potential sex-related genes and pathways in spotted knifejaw Oplegnathus punctatus. Gene 2017, 637, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Chenna, R.; Sugawara, H.; Koike, T.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Canestro, C.; Albalat, R.; Irimia, M.; Garcia-Fernàndez, J. Impact of gene gains, losses and duplication modes on the origin and diversification of vertebrates. In Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Hoffmann, F.G.; Opazo, J.C.; Storz, J.F. Whole-genome duplications spurred the functional diversification of the globin gene superfamily in vertebrates. Mol. Biol. Evol. 2011, 29, 303–312. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef]
- Ohno, S. Evolution by Gene Duplication; Springer Science & Business Media: Berlin, Germany, 1970. [Google Scholar]
- Bailey, J.A.; Gu, Z.; Clark, R.A.; Reinert, K.; Samonte, R.V.; Schwartz, S.; Adams, M.D.; Myers, E.W.; Li, P.W.; Eichler, E.E. Recent segmental duplications in the human genome. Science 2002, 297, 1003–1007. [Google Scholar] [CrossRef]
- Long, M.; Betrán, E.; Thornton, K.; Wang, W. The origin of new genes: Glimpses from the young and old. Nat. Rev. Genet. 2003, 4, 865–875. [Google Scholar] [CrossRef]
- Samonte, R.V.; Eichler, E.E. Segmental duplications and the evolution of the primate genome. Nat. Rev. Genet. 2002, 3, 65–72. [Google Scholar] [CrossRef]
- Aury, J.-M.; Jaillon, O.; Duret, L.; Noel, B.; Jubin, C.; Porcel, B.M.; Ségurens, B.; Daubin, V.; Anthouard, V.; Aiach, N. Global trends of whole-genome duplications revealed by the ciliate Paramecium tetraurelia. Nature 2006, 444, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.H.; Shields, D.C. Molecular evidence for an ancient duplication of the entire yeast genome. Nature 1997, 387, 708–712. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.-P.; Rosenberg, H.F. Adaptive evolution of a duplicated pancreatic ribonuclease gene in a leaf-eating monkey. Nat. Genet. 2002, 30, 411–415. [Google Scholar] [CrossRef]
- Baron, D.; Cocquet, J.; Xia, X.; Fellous, M.; Guiguen, Y.; Veitia, R.A. An evolutionary and functional analysis of FoxL2 in rainbow trout gonad differentiation. J. Mol. Endocrinol. 2004, 33, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, R.; Wang, R.; Chen, S. Transcriptomics analysis revealing candidate networks and genes for the body size sexual dimorphism of Chinese tongue sole (Cynoglossus semilaevis). Funct. Integr. Genom. 2018, 18, 327–339. [Google Scholar] [CrossRef]
- Dong, X.; Chen, S.; Ji, X.; Shao, C. Molecular cloning, characterization and expression analysis of Sox9a and Foxl2 genes in half-smooth tongue sole (Cynoglossussemilaevis). Acta Oceanol. Sin. 2011, 30, 68–77. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Horiguchi, R.; Nozu, R.; Nakamura, M. Expression and localization of forkhead transcriptional factor 2 (Foxl2) in the gonads of protogynous wrasse, Halichoeres trimaculatus. Biol. Sex Differ. 2010, 1, 1–9. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree | Model | lnL | κ | Null | LRT | df | p-Value | Site | BEB |
|---|---|---|---|---|---|---|---|---|---|
| foxl2 | M0 | −3160.8085 | 2.55419 | NA | |||||
| M1a | −3154.2940 | 2.60048 | NA | ||||||
| M2a | −3154.2940 | 2.60048 | M1a | 0 | 2 | 1 | |||
| M3 | −3134.7381 | 2.53049 | M0 | 52.14 | 4 | 1.29 × 10−10 | |||
| M7 | −3134.8327 | 2.53409 | NA | ||||||
| M8a | −3134.7717 | 2.53271 | NA | ||||||
| M8 | −3134.8345 | 2.53410 | M7 | 0.004 | 2 | 0.998 | |||
| M8a | 0.13 | 1 | 0.718 | ||||||
| foxl2l | M0 | −3681.4694 | 1.87394 | NA | |||||
| M1a | −3658.2278 | 1.95570 | NA | ||||||
| M2a | −3658.2278 | 1.95570 | M1a | 0 | 2 | 1 | |||
| M3 | −3612.7257 | 1.93108 | M0 | 137.5 | 4 | 0.0000 | 41S | 0.954 * | |
| M7 | −3615.4003 | 1.94152 | NA | ||||||
| M8a | −3615.1257 | 1.94138 | NA | ||||||
| M8 | −3614.8491 | 1.94016 | M7 | 1.1 | 2 | 0.577 | |||
| M8a | 0.55 | 1 | 0.458 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, X.; Yu, H.; Wang, Y.; Liu, J.; Zhang, Q. Comparative Studies on Duplicated foxl2 Paralogs in Spotted Knifejaw Oplegnathus punctatus Show Functional Diversification. Genes 2023, 14, 1847. https://doi.org/10.3390/genes14101847
Du X, Yu H, Wang Y, Liu J, Zhang Q. Comparative Studies on Duplicated foxl2 Paralogs in Spotted Knifejaw Oplegnathus punctatus Show Functional Diversification. Genes. 2023; 14(10):1847. https://doi.org/10.3390/genes14101847
Chicago/Turabian StyleDu, Xinxin, Haiyang Yu, Yujue Wang, Jinxiang Liu, and Quanqi Zhang. 2023. "Comparative Studies on Duplicated foxl2 Paralogs in Spotted Knifejaw Oplegnathus punctatus Show Functional Diversification" Genes 14, no. 10: 1847. https://doi.org/10.3390/genes14101847