
Distinct Gut Microbiome Induced by Different Feeding Regimes in Weaned Piglets

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sample Collection
2.3. Serum Metabolism Indicators
2.4. DNA Extraction and 16S rRNA Sequencing
2.5. Microbial Community Analysis
2.6. Statistical Analysis
3. Results
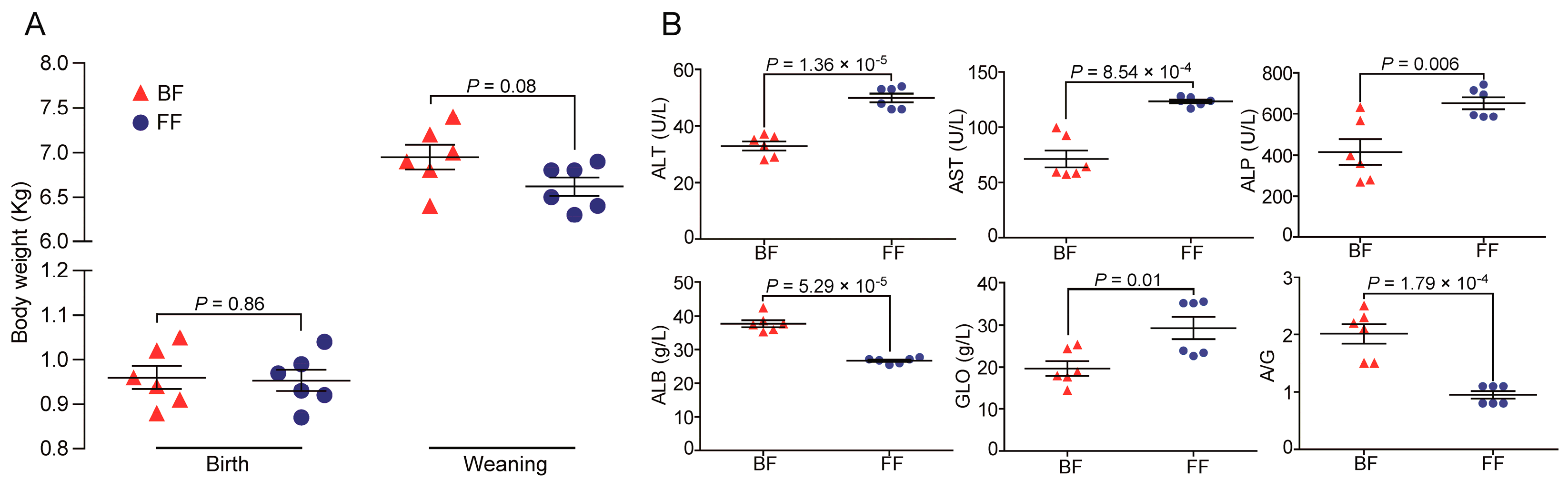
3.1. Phenotype Characteristics
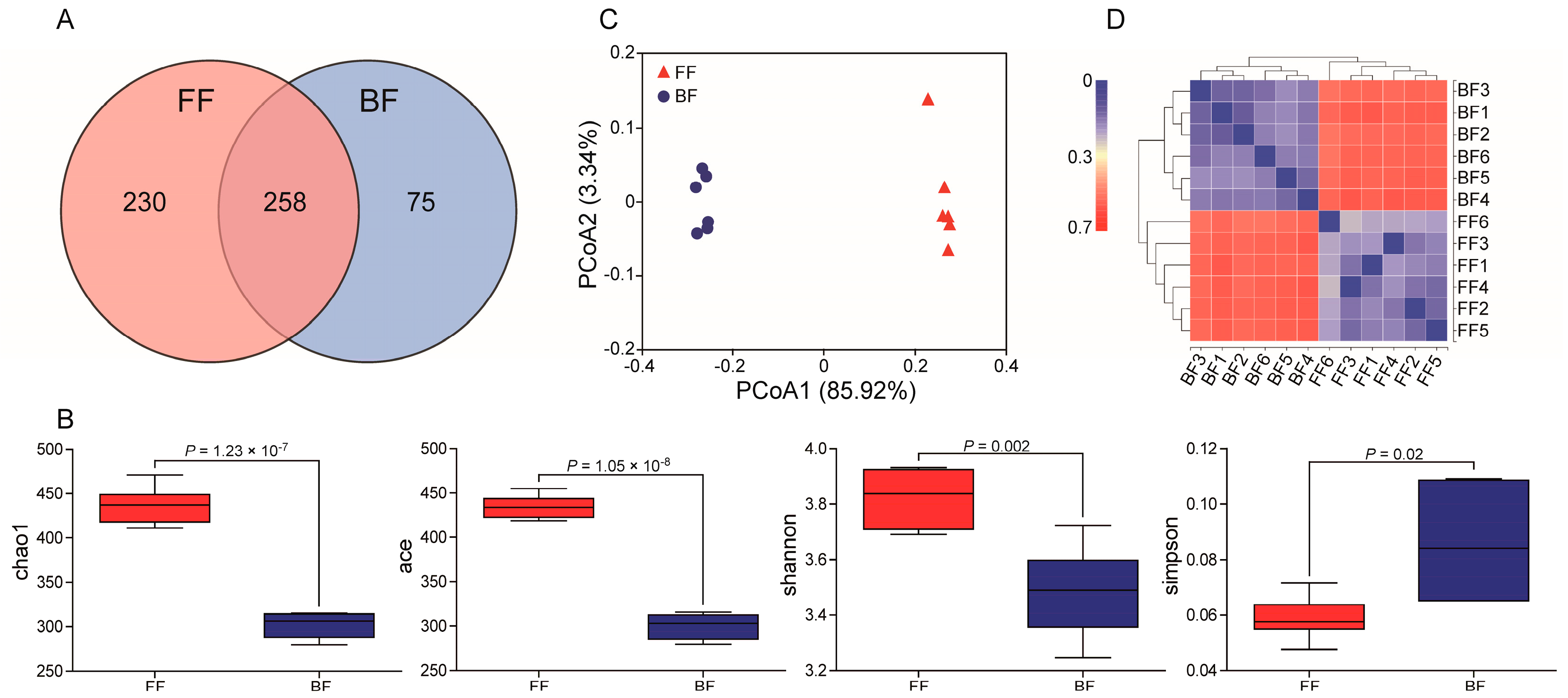
3.2. Microbiota Compositions
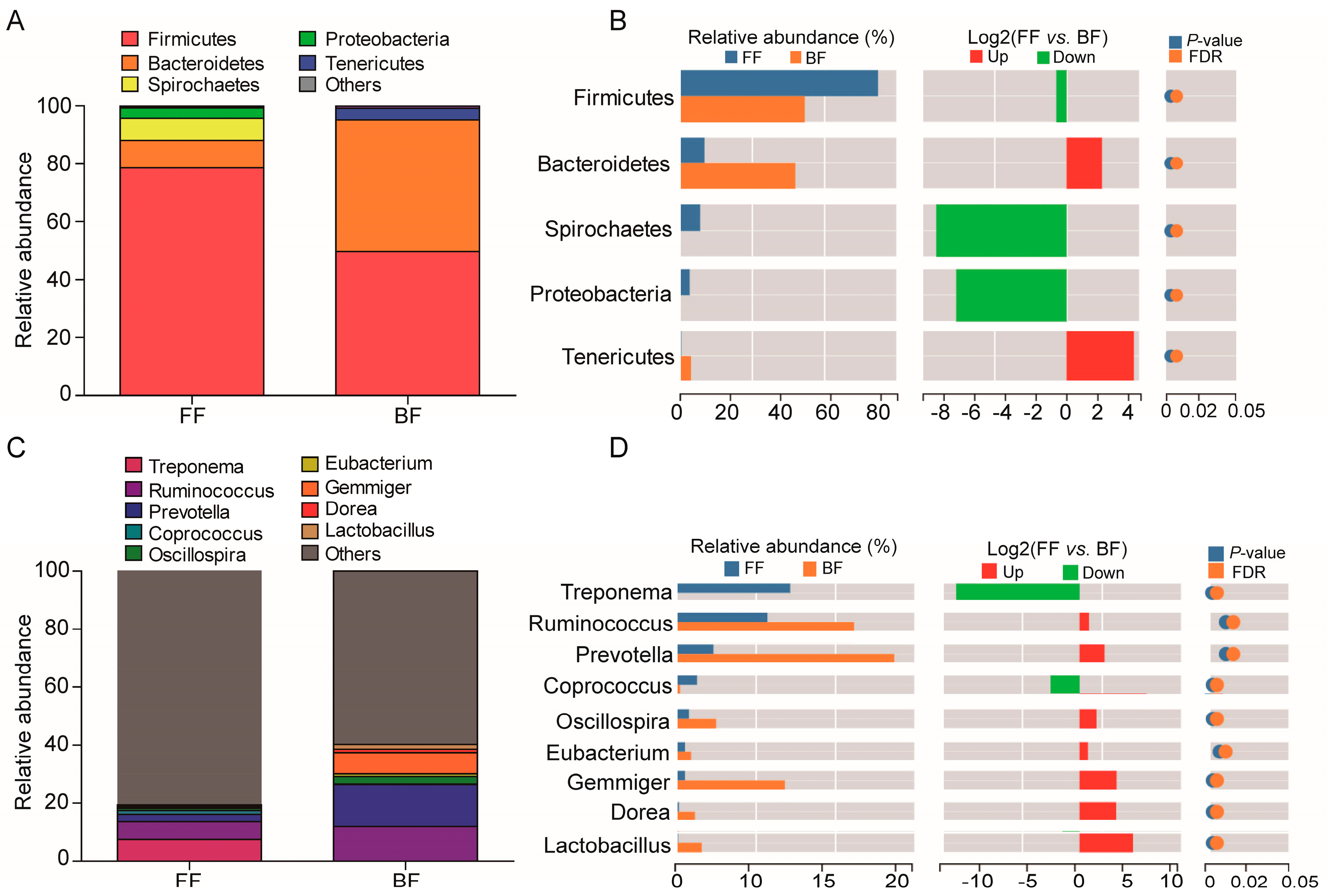
3.3. Microbial Phyla and Genera
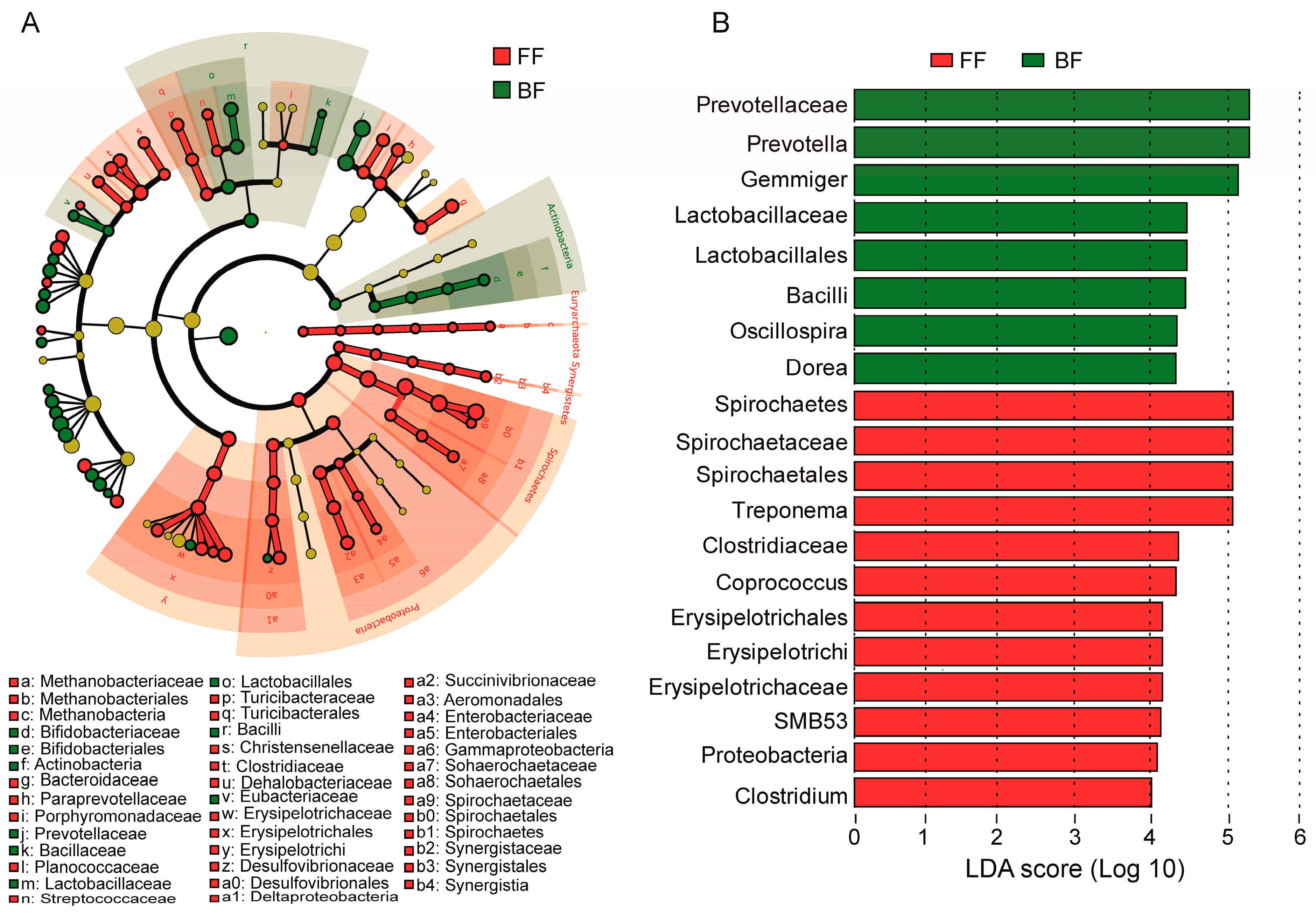
3.4. Differences in Microbiota
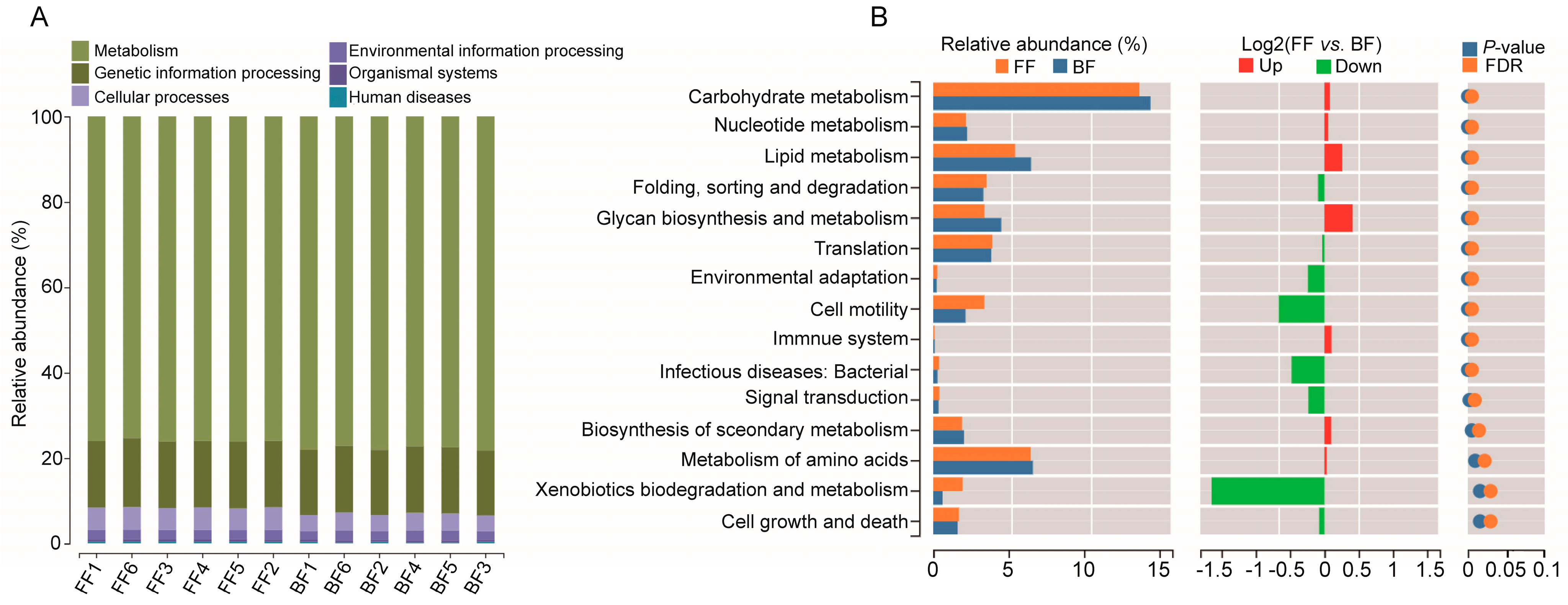
3.5. Potential Functional Prediction
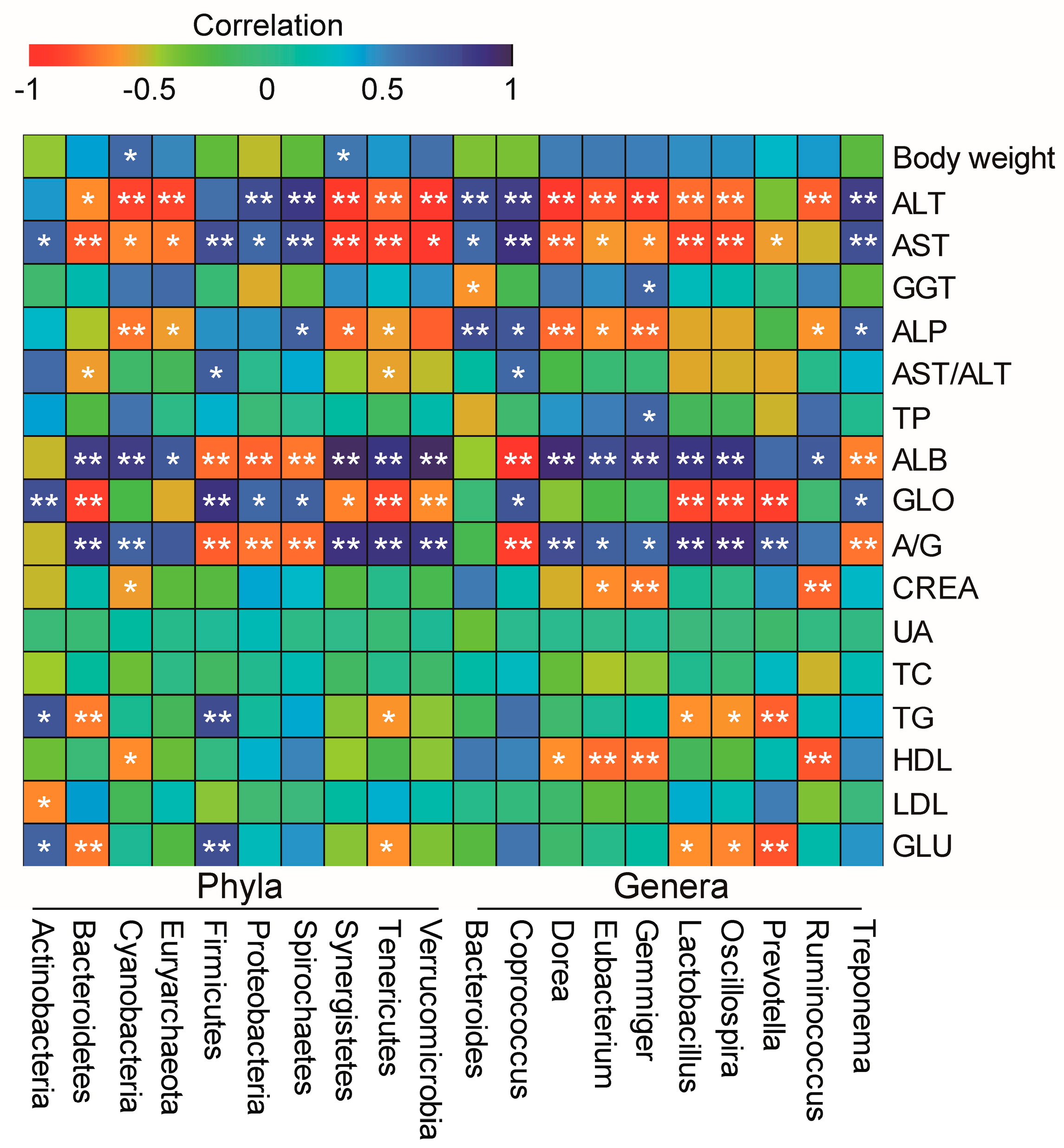
3.6. Associations between Microbiota and Phenotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martin, C.R.; Ling, P.R.; Blackburn, G.L. Review of infant feeding: Key features of breast milk and Infant formula. Nutrients 2016, 8, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Li, M.; Wang, T.; Liang, Y.; Zhong, Z.; Wang, X.; Zhou, Q.; Chen, L.; Lang, Q.; He, Z.; et al. Lactation-related microRNA expression profiles of porcine breast milk exosomes. PLoS ONE 2012, 7, e43691. [Google Scholar] [CrossRef] [PubMed]
- Parigi, S.M.; Eldh, M.; Larssen, P.; Gabrielsson, S.; Villablanca, E.J. Breast milk and solid food shaping intestinal immunity. Front. Immunol. 2015, 6, 415. [Google Scholar] [CrossRef] [Green Version]
- Moossavi, S.; Sepehri, S.; Robertson, B.; Bode, L.; Goruk, S.; Field, C.J.; Lix, L.M.; de Souza, R.J.; Becker, A.B.; Mandhane, P.J.; et al. Composition and variation of the human milk microbiota are influenced by maternal and early-Life factors. Cell. Host. Microbe. 2019, 25, 324–335. [Google Scholar] [CrossRef] [Green Version]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K.; et al. Association Between Breast Milk Bacterial Communities and Establishment and Development of the Infant Gut Microbiome. J. AMA. Pediatr. 2017, 171, 647–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Gallego, C.; Garcia-Mantrana, I.; Salminen, S.; Collado, M.C. The human milk microbiome and factors influencing its composition and activity. Semin. Fetal. Neonatal. Med. 2016, 21, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Yan, F.; Wang, N.; Song, Y.; Yue, Y.; Guan, J.; Li, B.; Huo, G. Distinct gut microbiota and metabolite profiles induced by different feeding methods in healthy chinese infants. Front. Microbiol. 2020, 11, 714. [Google Scholar] [CrossRef]
- Bode, L.; Mcguire, M.; Rodriguez, J.M.; Geddes, D.T.; Hassiotou, F.; Hartmann, P.E.; McGuire, M.K. It’s alive: Microbes and cells in human milk and their potential benefits to mother and infant. Adv. Nutr. 2014, 5, 571–573. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.; Makino, H.; Cetinyurek Yavuz, A.; Ben-Amor, K.; Roelofs, M.; Ishikawa, E.; Kubota, H.; Swinkels, S.; Sakai, T.; Oishi, K.; et al. Early-life events, including mode of delivery and type of feeding, siblings and gender, shape the developing gut microbiota. PLoS ONE 2016, 11, e0158498. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, L.; Moles, L.; Gueimonde, M.; Rodriguez, J.M. Perinatal microbiomes influence on preterm birth and preterms’ health: Influencing factors and modulation strategies. J. Pediatr. Gastroenterol. Nutr. 2016, 63, e193–e203. [Google Scholar] [CrossRef]
- O’Sullivan, A.; Farver, M.; Smilowitz, J.T. The influence of early infant-feeding practices on the intestinal microbiome and body composition in infants. Erratum Nutr. Metab. Insights 2016, 8, 87. [Google Scholar] [PubMed]
- Rendina, D.N.; Lubach, G.R.; Phillips, G.J.; Lyte, M.; Coe, C.L. Maternal and breast milk influences on the infant gut microbiome, enteric health and growth outcomes of rhesus monkeys. J. Pediatr. Gastr. Nutr. 2019, 69, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Dewey, K.G.; Heinig, M.J.; Nommsen, L.A.; Peerson, J.M.; Lönnerdal, B. Breast-fed infants are leaner than formula-fed infants at 1 y of age: The DARLING study. Am. J. Clin. Nutr. 1993, 57, 140–145. [Google Scholar] [CrossRef]
- Thompson, F.M.; Catto-smith, A.G.; Moore, D.; Davidson, G.; Cummins, A.G. Epithelial growth of the small intestine in human infants. J. Pediatr. Gastroenterol. Nutr. 1998, 26, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Kanno, T.; Koyanagi, N.; Katoku, Y.; Yonekubo, A.; Yajima, T.; Kuwata, T.; Kitagawa, H.; Harada, E. Simplified preparation of a refined milk formula comparable to rat’s milk: Influence of the formula on development of the gut and brain in artificially reared rat pups. J. Pediatr. Gastroenterol. Nutr. 1997, 24, 242–252. [Google Scholar] [CrossRef]
- Dvorak, B.; McWilliam, D.L.; Williams, C.S.; Dominguez, J.A.; Machen, N.W.; McCuskey, R.S.; Philipps, A.F. Artificial formula induces precocious maturation of the small intestine of artificially reared suckling rats. J. Pediatr. Gastroenterol. Nutr. 2000, 31, 162–169. [Google Scholar] [CrossRef]
- Boudry, G.; Morise, A.; Seve, B.; LE Huërou-Luron, I. Effect of milk formula protein content on intestinal barrier function in a porcine model of LBW neonates. Pediatr. Res. 2011, 69, 4–9. [Google Scholar] [CrossRef] [Green Version]
- Pieper, R.; Scharek-Tedin, L.; Zetzsche, A.; Röhe, I.; Kröger, S.; Vahjen, W.; Zentek, J. Bovine milk-based formula leads to early maturation-like morphological, immunological, and functional changes in the jejunum of neonatal piglets. J. Anim. Sci. 2016, 94, 989–999. [Google Scholar] [CrossRef] [Green Version]
- Hvistendahl, M. Pigs as stand-ins for microbiome studies. Science 2012, 336, 1250. [Google Scholar]
- Poroyko, V.; White, J.R.; Wang, M.; Donovan, S.; Alverdy, J.; Liu, D.C.; Morowitz, M.J. Gut microbial gene expression in mother-fed and formula-fed piglets. PLoS ONE 2010, 5, e12459. [Google Scholar] [CrossRef] [Green Version]
- Timmerman, H.M.; Rutten, N.; Boekhorst, J.; Saulnier, D.M.; Kortman, G.; Contractor, N.; Kullen, M.; Floris, E.; Harmsen, H.; Vlieger, A.M.; et al. Intestinal colonisation patterns in breastfed and formula-fed infants during the first 12 weeks of life reveal sequential microbiota signatures. Sci. Rep. 2017, 7, 8327. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, Z.; Zhang, W.; Zhang, C.; Zhang, Y.; Mei, H.; Zhuo, N.; Wang, H.; Wang, L.; Wu, D. Comparison of gut microbiota in exclusively breast-fed and formula-fed babies: A study of 91 term infants. Sci. Rep. 2020, 10, 15792. [Google Scholar] [CrossRef]
- Heinig, M.J.; Nommsen, L.A.; Peerson, J.M.; Lonnerdal, B.; Dewey, K.G. Energy and protein intakes of breast-fed and formula-fed infants during the first year of life and their association with growth velocity: The DARLING Study. Am. J. Clin. Nutr. 1993, 58, 152–161. [Google Scholar] [CrossRef] [Green Version]
- le Huërou-Luron, I.; Blat, S.; Boudry, G. Breast- v. formula-feeding: Impacts on the digestive tract and immediate and long-term health effects. Nutr. Res. Rev. 2010, 23, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Ekelund, U.; Ong, K.K.; Linné, Y.; Neovius, M.; Brage, S.; Dunger, D.B.; Wareham, N.J.; Rössner, S. Association of weight gain in infancy and early childhood with metabolic risk in young adults. J. Clin. Endocrinol. Metab. 2007, 92, 98–103. [Google Scholar] [CrossRef] [Green Version]
- Michaelsen, K.F.; Greer, F.R. Protein needs early in life and long-term health. Am. J. Clin. Nutr. 2014, 99, 718S–722S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Chen, T.; Jiang, R.; Zhao, A.; Wu, Q.; Kuang, J.; Sun, D.; Ren, Z.; Li, M.; Zhao, M.; et al. Hyocholic acid species improve glucose homeostasis through a distinct TGR5 and FXR signaling mechanism. Cell. Metab. 2021, 33, 791–803.e7. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.; Mercado, V.; Rios, M.; Arboleda, R.; Gomara, R.; Muinos, W.; Reeves-Garcia, J.; Hernandez, E. Breast milk is better than formula milk in preventing parenteral nutrition-associated liver disease in infants receiving prolonged parenteral nutrition. J. Pediatr. Gastr. Nutr. 2013, 57, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Saarinen, U.M.; Pelkonen, P.; Siimes, M.A. Serum immunoglobulin A in healthy infants: An accelerated postnatal increase in formula-fed compared to breast-fed infants. J. Pediatr. 1979, 95, 410–412. [Google Scholar] [CrossRef]
- Roger, L.C.; Costabile, A.; Holland, D.T.; Hoyles, L.; McCartney, A.L. Examination of faecal Bifidobacterium populations in breast- and formula-fed infants during the first 18 months of life. Microbiology 2010, 156, 3329–3341. [Google Scholar] [CrossRef] [Green Version]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell. Host. Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorvitovskaia, A.; Holmes, S.P.; Huse, S.M. Interpreting Prevotella and Bacteroides as biomarkers of diet and lifestyle. Microbiome 2016, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 4578–4585. [Google Scholar] [CrossRef] [Green Version]
- Levin, A.M.; Sitarik, A.R.; Havstad, S.L.; Fujimura, K.E.; Wegienka, G.; Cassidy-Bushrow, A.E.; Kim, H.; Zoratti, E.M.; Lukacs, N.W.; Boushey, H.A.; et al. Joint effects of pregnancy, sociocultural, and environmental factors on early life gut microbiome structure and diversity. Sci. Rep-UK 2016, 6, 31775. [Google Scholar] [CrossRef] [Green Version]
- Walker, W.A.; Iyengar, R.S. Breast milk, microbiota and intestinal immune homeostasis. Pediatr. Res. 2015, 77, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Ponzo, V.; Ferrocino, I.; Zarovska, A.; Amenta, M.B.; Leone, F.; Monzeglio, C.; Rosato, R.; Pellegrini, M.; Gambino, R.; Cassader, M.; et al. The microbiota composition of the offspring of patients with gestational diabetes mellitus (GDM). PLoS ONE 2019, 14, e0226545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coppa, G.V.; Gabrielli, O.; Zampini, L.; Galeazzi, T.; Ficcadenti, A.; Padella, L.; Santoro, L.; Soldi, S.; Carlucci, A.; Bertino, E.; et al. Oligosaccharides in 4 different milk groups, Bifidobacteria, and Ruminococcus obeum. J. Pediatr. Gastr. Nutr. 2011, 53, 80–87. [Google Scholar] [CrossRef] [PubMed]
- de Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [Green Version]
- Solbach, P.; Chhatwal, P.; Woltemate, S.; Tacconelli, E.; Buhl, M.; Autenrieth, I.B.; Vehreschild, M.; Jazmati, N.; Gerhard, M.; Stein-Thoeringer, C.K.; et al. Microbiota-associated risk factors for clostridioides difficile acquisition in hospitalized patients: A prospective, multicentric study. Clin. Infect. Dis. 2021, 73, e2625–e2634. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Gao, Z.; Yang, F.; Yang, Y.; Chen, L.; Han, L.; Zhao, N.; Xu, J.; Wang, X.; Ma, Y.; et al. Antidiabetic effects of gegen qinlian decoction via the gut microbiota are attributable to its key ingredient berberine. Genom. Proteom. Bioinform. 2020, 18, 721–736. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Zhao, X.; Cui, X.; Mao, X.; Tang, N.; Jiao, C.; Wang, D.; Zhang, Y.; Ye, Z.; Zhang, H. Characterization of fungal and bacterial dysbiosis in young adult Chinese patients with Crohn’s disease. Therap. Adv. Gastroenter. 2020, 13, 1756284820971202. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Sabino, J.; Deroover, L.; Vandermeulen, G.; Luypaerts, A.; Ferrante, M.; Vermeire, S. P774 Metagenomics and metabolomics of patients with inflammatory bowel disease and their unaffected relatives. J. Crohns. Colitis. 2017, 11, 476–477. [Google Scholar] [CrossRef] [Green Version]
- Breuninger, T.A.; Wawro, N.; Breuninger, J.; Reitmeier, S.; Clavel, T.; Six-Merker, J.; Pestoni, G.; Rohrmann, S.; Rathmann, W.; Peters, A.; et al. Associations between habitual diet, metabolic disease, and the gut microbiota using latent Dirichlet allocation. Microbiome 2021, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Steed, H.; Macfarlane, S. Bacterial metabolism and health-related effects of galacto-oligosaccharides and other prebiotics. J. Appl. Microbiol. 2008, 104, 305–344. [Google Scholar] [CrossRef]
- Madan, J.C.; Hoen, A.G.; Lundgren, S.N.; Farzan, S.F.; Cottingham, K.L.; Morrison, H.G.; Sogin, M.L.; Li, H.; Moore, J.H.; Karagas, M.R. Association of cesarean delivery and formula supplementation with the intestinal microbiome of 6-week-old infants. JAMA Pediatr. 2016, 170, 212–219. [Google Scholar] [CrossRef]
- Ardeshir, A.; Narayan, N.R.; Méndez-Lagares, G.; Lu, D.; Rauch, M.; Huang, Y.; Van Rompay, K.K.; Lynch, S.V.; Hartigan-O’Connor, D.J. Breast-fed and bottle-fed infant rhesus macaques develop distinct gut microbiotas and immune systems. Sci. Transl. Med. 2014, 6, 252ra120. [Google Scholar] [CrossRef] [Green Version]
- Bezirtzoglou, E.; Tsiotsias, A.; Welling, G.W. Microbiota profile in feces of breast- and formulafed newborns by using fluorescence in situ hybridization (FISH). Anaerobe 2011, 17, 478–482. [Google Scholar] [CrossRef]
- Fanaro, S.; Boehm, G.; Garssen, J.; Knol, J.; Mosca, F.; Stahl, B.; Vigi, V. Galacto-oligosaccharides and long-chain fructo-oligosaccharides as prebiotics in infant formulas: A review. Acta. Paediatr. 2005, 94, 22–26. [Google Scholar] [CrossRef]
- Pärtty, A.; Luoto, R.; Kalliomäki, M.; Salminen, S.; Isolauri, E. Effects of early prebiotic and probiotic supplementation on development of gut microbiota and fussing and crying in preterm infants: A randomized, double-blind, placebo-controlled trial. J. Pediatr-US. 2013, 163, 1272–1277. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Huang, J.; Jin, Q.; Guo, Z.; Liu, Y.; Cheong, L.; Xu, X.; Wang, X. Lipid composition analysis of milk fats from different mammalian species: Potential for use as human milk fat substitutes. J. Agric. Food. Chem. 2013, 61, 7070–7080. [Google Scholar] [CrossRef] [PubMed]
- Luoto, R.; Ruuskanen, O.; Waris, M.; Kalliomäki, M.; Salminen, S.; Isolauri, E. Prebiotic and probiotic supplementation prevents rhinovirus infections in preterm infants: A randomized, placebo-controlled trial. J. Allergy. Clin. Immunol. 2014, 133, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Siy, P.N.; Larson, R.T.; Zembsch, T.E.; Lee, X.; Paskewitz, S.M. High Prevalence of Borrelia mayonii (Spirochaetales: Spirochaetaceae) in Field-Caught Tamias striatus (Rodentia: Sciuridae) From Northern Wisconsin. J. Med. Entomol. 2021, 58, 2504–2507. [Google Scholar] [CrossRef]
- Lee, S.H.; Goo, Y.K.; Geraldino, P.; Kwon, O.D.; Kwak, D. Molecular Detection and Characterization of Borrelia garinii (Spirochaetales: Borreliaceae) in Ixodes nipponensis (Ixodida: Ixodidae) Parasitizing a Dog in Korea. Pathogens 2019, 8, 289. [Google Scholar] [CrossRef] [Green Version]
- Allenspach, K.; House, A.; Smith, K.; McNeill, F.M.; Hendricks, A.; Elson-Riggins, J.; Riddle, A.; Steiner, J.M.; Werling, D.; Garden, O.A.; et al. Evaluation of mucosal bacteria and histopathology, clinical disease activity and expression of Toll-like receptors in German shepherd dogs with chronic enteropathies. Vet. Microbiol. 2010, 146, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Dinh, D.M.; Volpe, G.E.; Duffalo, C.; Bhalchandra, S.; Tai, A.K.; Kane, A.V.; Wanke, C.A.; Ward, H.D. Intestinal microbiota, microbial translocation, and systemic inflammation in chronic HIV infection. J. Infect. Dis. 2015, 211, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan, N.; Knotts, T.A.; Pesapane, R.; Ramsey, J.J.; Castle, S.; Clifford, D.; Foley, J. Conservation Implications of Shifting Gut Microbiomes in Captive-Reared Endangered Voles Intended for Reintroduction into the Wild. Microorganisms 2018, 6, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnusson, K.R.; Hauck, L.; Jeffrey, B.M.; Elias, V.; Humphrey, A.; Nath, R.; Perrone, A.; Bermudez, L.E. Relationships between diet-related changes in the gut microbiome and cognitive flexibility. Neuroscience 2015, 300, 128–140. [Google Scholar] [CrossRef]
- Cani, P.D.; Van Hul, M.; Lefort, C.; Depommier, C.; Rastelli, M.; Everard, A. Microbial regulation of organismal energy homeostasis. Nat. Metab. 2019, 1, 34–46. [Google Scholar] [CrossRef] [Green Version]
- González, R.; Klaassens, E.S.; Malinen, E.; de Vos, W.M.; Vaughan, E.E. Differential transcriptional response of Bifidobacterium longum to human milk, formula milk, and galactooligosaccharide. Appl. Environl. Microb. 2018, 74, 4686–4694. [Google Scholar] [CrossRef] [Green Version]
- Eidels, L.; Osborn, M.J. Lipopolysaccharide and aldoheptose biosynthesis in transketolase mutants of Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 1971, 68, 1673–1677. [Google Scholar] [CrossRef] [PubMed]
- Baumann-Dudenhoeffer, A.M.; D’Souza, A.W.; Tarr, P.I.; Warner, B.B.; Dantas, G. Infant diet and maternal gestational weight gain predict early metabolic maturation of gut microbiomes. Nat. Med. 2018, 24, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.S.; Shah, V.S.; Kelly, L.E. Arginine supplementation for prevention of necrotising enterocolitis in preterm infants. Cochrane. Database. Syst. Rev. 2017, 4, CD004339. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, Y.; Ma, F.; Sun, M.; Mu, G.; Tuo, Y. The ameliorative effect of Lactobacillus plantarum Y44 oral administration on inflammation and lipid metabolism in obese mice fed with a high fat diet. Food. Funct. 2020, 11, 5024–5039. [Google Scholar] [CrossRef]
- Mohamed, K.; El-Katcha, M.; Ismail, E.Y.; Soltan, M.A. Effect of dietary probiotics supplementation on growth performance, immune response, some blood parameters and carcass quality of growing rabbits. Alex. J. Vet. Sci. 2011, 34, 153–169. [Google Scholar]
- Xu, R.H.; Xiu, L.; Zhang, Y.L.; Du, R.P.; Wang, X. Probiotic and hepatoprotective activity of lactobacillus isolated from Mongolian camel milk products. Benef. Microbes 2019, 10, 699–710. [Google Scholar] [CrossRef]
- Yue, S.; Zhao, D.; Peng, C.; Tan, C.; Wang, Q.; Gong, J. Effects of theabrownin on serum metabolites and gut microbiome in rats with a high-sugar diet. Food. Function. 2019, 10, 7063–7080. [Google Scholar] [CrossRef]
- Moreno-Navarrete, J.M.; Serino, M.; Blasco-Baque, V.; Azalbert, V.; Barton, R.H.; Cardellini, M.; Latorre, J.; Ortega, F.; Sabater-Masdeu, M.; Burcelin, R.; et al. Gut Microbiota Interacts with Markers of Adipose Tissue Browning, Insulin Action and Plasma Acetate in Morbid Obesity. Mol. Nutr. Food. Res. 2018, 62, 170721. [Google Scholar] [CrossRef]
- Holmes, E.; Nicholson, J. Variation in gut microbiota strongly influences individual rodent phenotypes. Toxicol. Sci. 2005, 87, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Olivares, P.; Pacheco, A.; Aranha, L.N.; Oliveira, B.; Santos, A.A.; Santos, P.; Neto, J.; Rosa, G.; Oliveira, G. Gut microbiota of adults with different metabolic phenotypes. Nutrition 2021, 90, 111293. [Google Scholar] [CrossRef]
- Stanislawski, M.A.; Dabelea, D.; Lange, L.A.; Wagner, B.D.; Lozupone, C.A. Gut microbiota phenotypes of obesity. NPJ Biofilms Microbiomes 2019, 5, 18. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Long, X.; Liao, Q.; Chai, J.; Zhang, T.; Chen, L.; He, H.; Yuan, Y.; Wan, K.; Wang, J.; et al. Distinct Gut Microbiome Induced by Different Feeding Regimes in Weaned Piglets. Genes 2023, 14, 49. https://doi.org/10.3390/genes14010049
Zhang J, Long X, Liao Q, Chai J, Zhang T, Chen L, He H, Yuan Y, Wan K, Wang J, et al. Distinct Gut Microbiome Induced by Different Feeding Regimes in Weaned Piglets. Genes. 2023; 14(1):49. https://doi.org/10.3390/genes14010049
Chicago/Turabian StyleZhang, Jie, Xi Long, Qinfeng Liao, Jie Chai, Tinghuan Zhang, Li Chen, Hang He, Yancong Yuan, Kun Wan, Jinyong Wang, and et al. 2023. "Distinct Gut Microbiome Induced by Different Feeding Regimes in Weaned Piglets" Genes 14, no. 1: 49. https://doi.org/10.3390/genes14010049