
Antimicrobial Resistance and Genomic Epidemiology of tet(X4)-Bearing Bacteria of Pork Origin in Jiangsu, China

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Antimicrobial Susceptibility Testing
2.3. Conjugation Experiments
2.4. Whole Genome Sequencing
2.5. Bioinformatics Analysis
2.6. Data Availability
3. Results
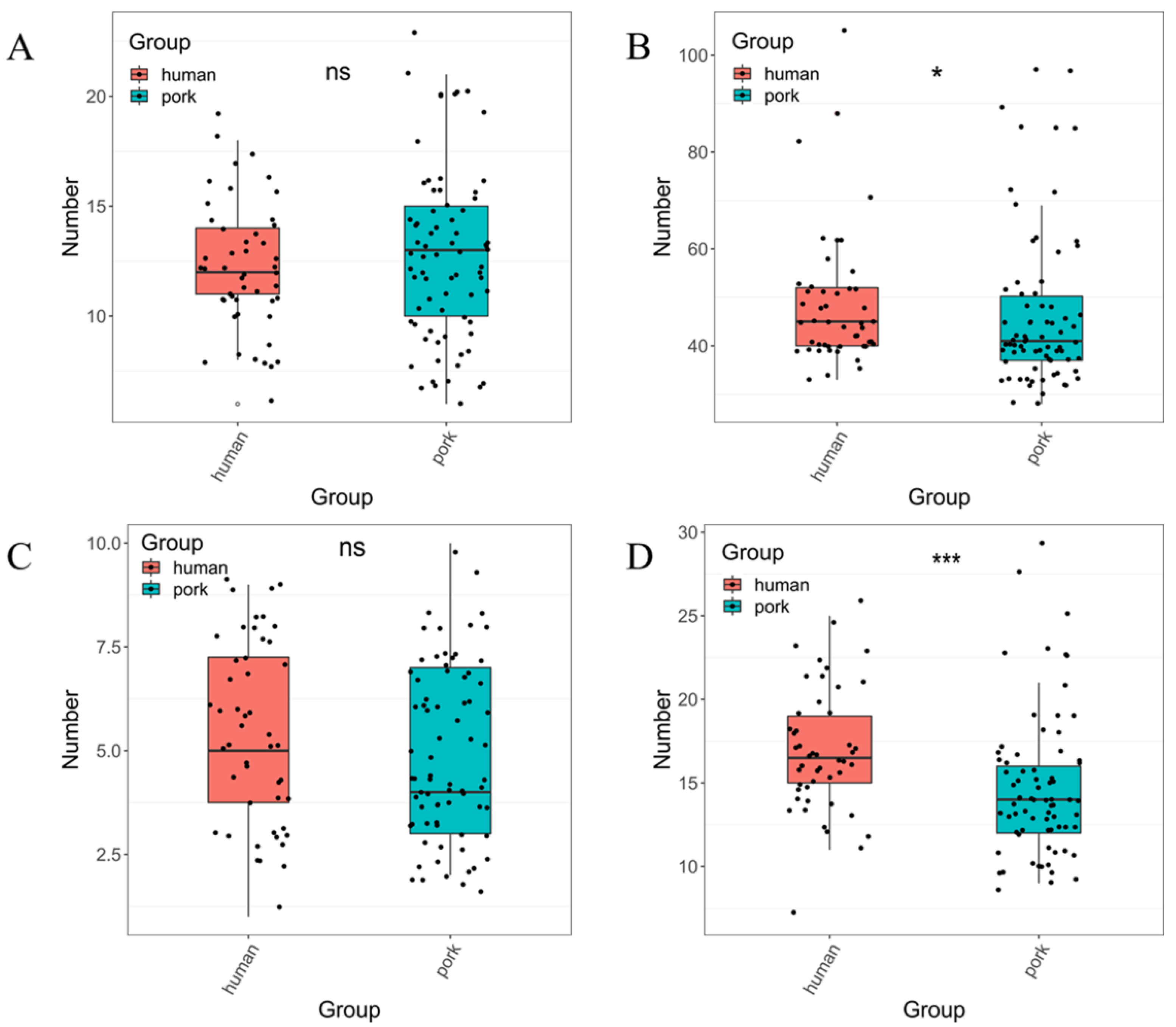
3.1. Characterization of tet(X4)-Bearing Isolates among Pork
3.2. Transferability of the tet(X4) Gene
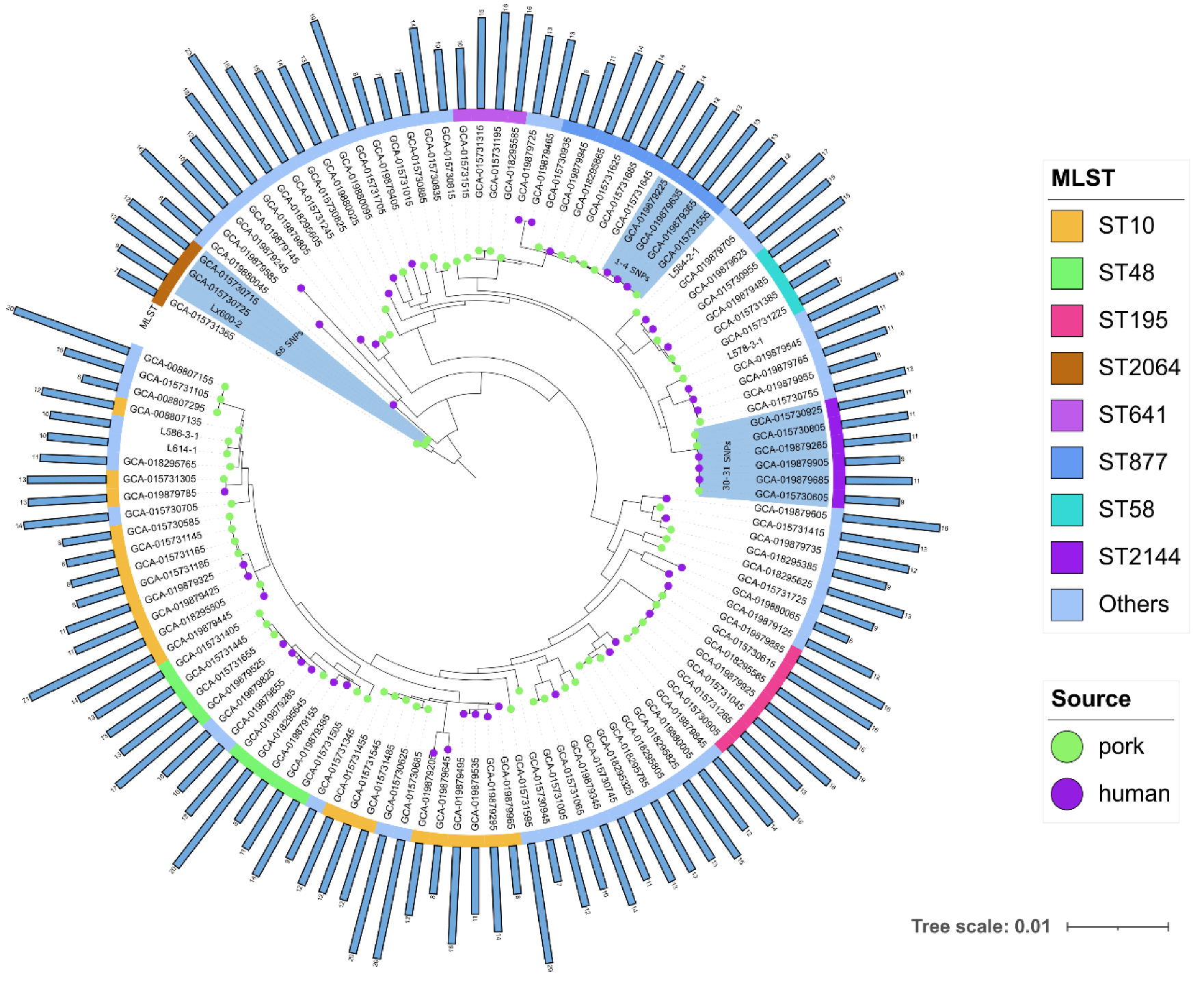
3.3. Phylogenetic Analysis of tet(X)-Positive E. coli
3.4. Genome Sequence Features of tet(X)-Positive E. coli
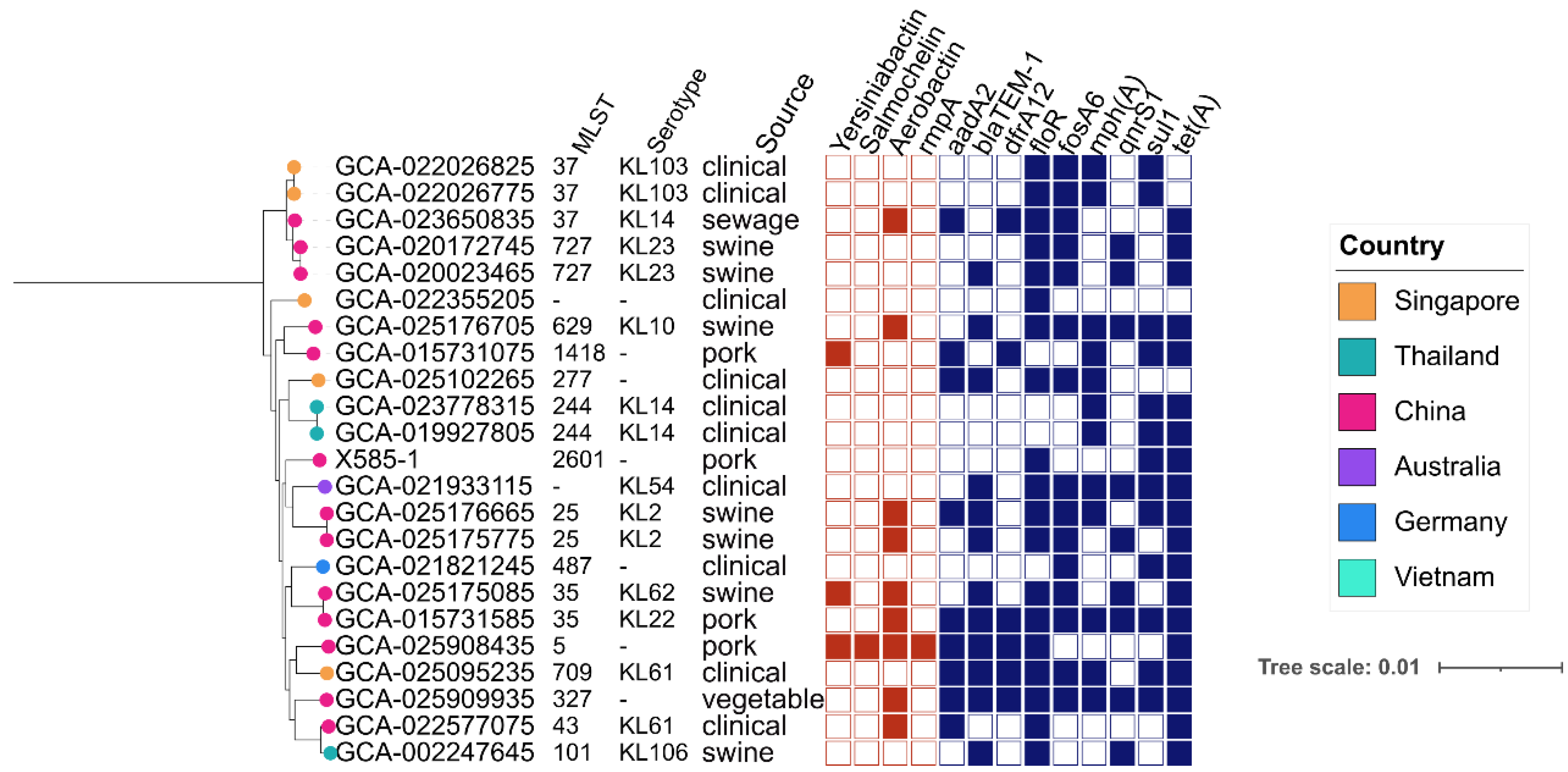
3.5. Phylogenetic Analysis of tet(X)-Positive K. pneumoniae
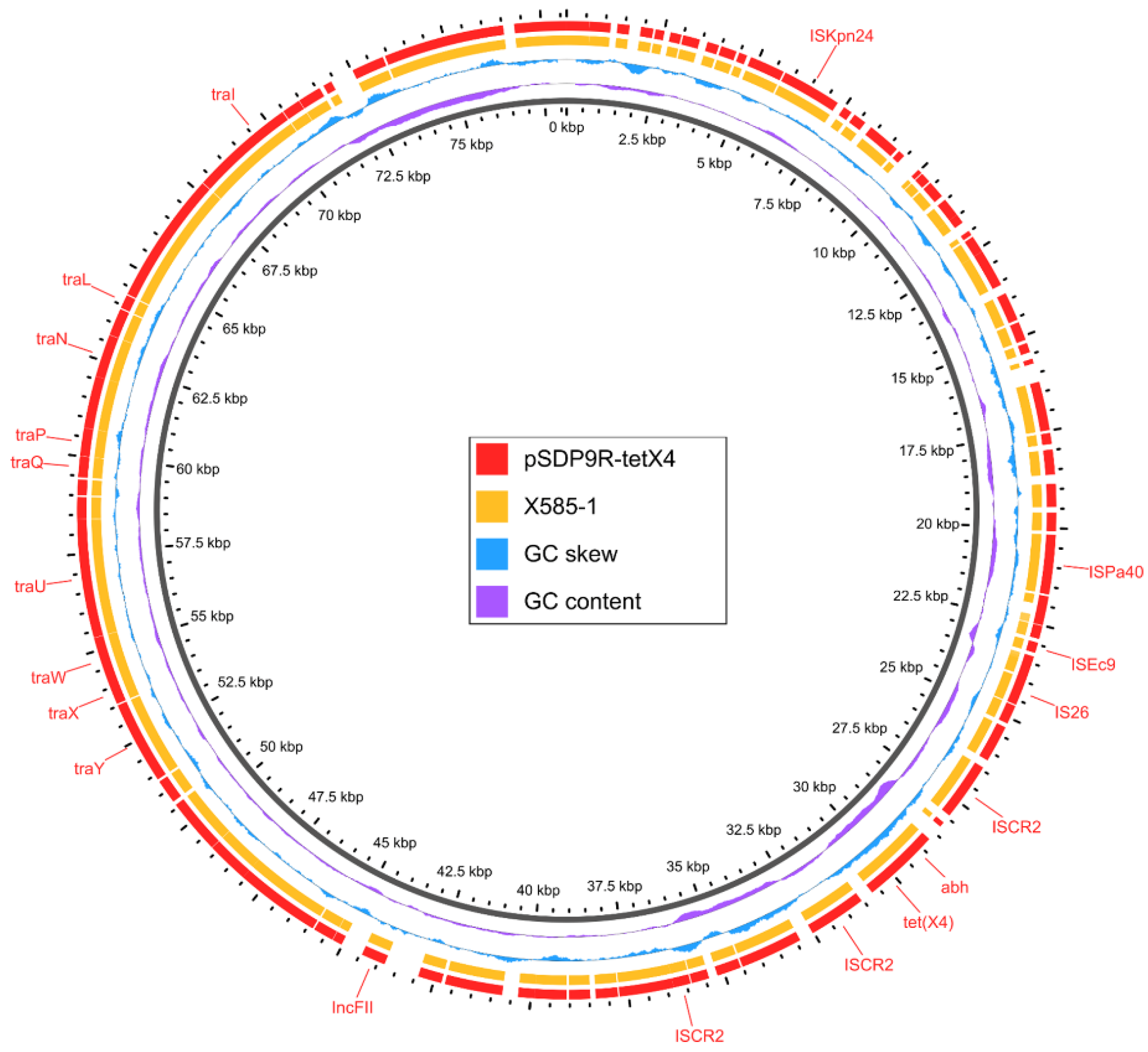
3.6. The Genetic Context of tet(X4) Carried by K. pneumoniae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Laxminarayan, R.; Sridhar, D.; Blaser, M.; Wang, M.; Woolhouse, M. Achieving global targets for antimicrobial resistance. Science 2016, 353, 874–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karageorgopoulos, D.E.; Falagas, M.E. Current control treatment of multidrug-resistant Acinetobacter baumannii infections. Lancet Infect. Dis. 2008, 8, 751–762. [Google Scholar] [CrossRef]
- Markley, J.L.; Wencewicz, T.A. Tetracycline-Inactivating Enzymes. Front. Microbiol. 2018, 9, 1058. [Google Scholar] [CrossRef] [Green Version]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [Green Version]
- Ahn, C.; Yoon, S.S.; Yong, T.S.; Jeong, S.H.; Lee, K. The Resistance Mechanism and Clonal Distribution of Tigecycline-Nonsusceptible Klebsiella pneumoniae Isolates in Korea. Yonsei Med. J. 2016, 57, 641–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, M.W.; Ruzin, A.; Feyfant, E.; Rush, T.S., III; O’Connell, J.; Bradford, P.A. Functional, biophysical, and structural bases for antibacterial activity of tigecycline. Antimicrob. Agents Chemother. 2006, 50, 2156–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, T.; Wang, R.; Liu, D.; Walsh, T.R.; Zhang, R.; Lv, Y.; Ke, Y.; Ji, Q.; Wei, R.; Liu, Z.; et al. Emergence of plasmid-mediated high-level tigecycline resistance genes in animals and humans. Nat. Microbiol. 2019, 4, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Lu, X.; Peng, K.; Liu, Z.; Li, Y.; Liu, Y.; Xiao, X.; Wang, Z. Deciphering the Structural Diversity and Classification of the Mobile Tigecycline Resistance Gene tet(X)-Bearing Plasmidome among Bacteria. mSystems 2020, 5, e00134-20. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Du, P.; Du, Y.; Sun, H.; Zhang, P.; Wan, Y.; Lin, Q.; Fanning, S.; Cui, S.; Wu, Y. Detection of plasmid-mediated tigecycline-resistant gene tet(X4) in Escherichia coli from pork, Sichuan and Shandong Provinces, China, February 2019. Eurosurveillance 2019, 24, 1900340. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Li, Y.; Peng, K.; Yin, Y.; Liu, Y.; He, T.; Bai, L.; Wang, Z. Comprehensive Genomic Investigation of Tigecycline Resistance Gene tet(X4)-Bearing Strains Expanding among Different Settings. Microbiol. Spectr. 2021, 9, e0163321. [Google Scholar] [CrossRef]
- Chen, C.; Cui, C.-Y.; Yu, J.-J.; He, Q.; Wu, X.-T.; He, Y.-Z.; Cui, Z.-H.; Li, C.; Jia, Q.-L.; Shen, X.-G.; et al. Genetic diversity and characteristics of high-level tigecycline resistance Tet(X) in Acinetobacter species. Genome Med. 2020, 12, 111. [Google Scholar] [CrossRef] [PubMed]
- Zhai, W.; Tian, Y.; Lu, M.; Zhang, M.; Song, H.; Fu, Y.; Ma, T.; Sun, C.; Bai, L.; Wang, Y.; et al. Presence of Mobile Tigecycline Resistance Gene tet(X4) in Clinical Klebsiella pneumoniae. Microbiol. Spectr. 2022, 10, e0108121. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Clinical and Laboratory Standards Institute [CLSI]. In Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Carattoli, A.; Zankari, E.; Garcìa-Fernandez, A.; Larsen, M.; Lund, O.; Voldby Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids. Antimicrob using PlasmidFinder and plasmid multilocus sequence typing. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinheinz, K.A.; Joensen, K.G.; Larsen, M.V. Applying the ResFinder and VirulenceFinder web-services for easy identification of acquired antibiotic resistance and E. coli virulence genes in bacteriophage and prophage nucleotide sequences. Bacteriophage 2014, 4, e27943. [Google Scholar] [CrossRef] [Green Version]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, Y.; Lu, J.; Wang, Q.; Cui, Y.; Wang, Y.; Quan, J.; Zhao, D.; Du, X.; Liu, H.; et al. Clinical relevance and plasmid dynamics of mcr-1-positive Escherichia coli in China: A multicentre case-control and molecular epidemiological study. Lancet Microbe 2020, 1, e24–e33. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing Large Minimum Evolution Trees with Profiles instead of a Distance Matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Sun, C.; Cui, M.; Zhang, S.; Liu, D.; Fu, B.; Li, Z.; Bai, R.; Wang, Y.; Wang, H.; Song, L.; et al. Genomic epidemiology of animal-derived tigecycline-resistant Escherichia coli across China reveals recent endemic plasmid-encoded tet(X4) gene. Commun. Biol. 2020, 3, 412. [Google Scholar] [CrossRef]
- Ding, Y.; Saw, W.-Y.; Tan, L.W.L.; Moong, D.K.N.; Nagarajan, N.; Teo, Y.Y.; Seedorf, H. Emergence of tigecycline- and eravacycline-resistant Tet(X4)-producing Enterobacteriaceae in the gut microbiota of healthy Singaporeans. J. Antimicrob. Chemother. 2020, 75, 3480–3484. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wang, T.; Shao, D.; Song, H.; Zhai, W.; Sun, C.; Zhang, Y.; Zhang, M.; Fu, Y.; Zhang, R.; et al. Structural diversity of the ISCR2-mediated rolling-cycle transferable unit carrying tet(X4). Sci. Total. Environ. 2022, 826, 154010. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Q.; Peng, K.; Liu, Y.; Xiao, X.; Mohsin, M.; Li, R.; Wang, Z. Distribution and genomic characterization of tigecycline-resistant tet(X4)-positive Escherichia coli of swine farm origin. Microb. Genom. 2021, 7, 000667. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Chen, K.; Ye, L.; Yang, X.; Xu, Q.; Yang, C.; Dong, N.; Chan, E.W.; Sun, Q.; Shu, L.; et al. Conjugation of Virulence Plasmid in Clinical Klebsiella pneumoniae Strains through Formation of a Fusion Plasmid. Adv. Biosyst. 2020, 4, e1900239. [Google Scholar] [CrossRef]
- Yang, X.; Chan, E.W.-C.; Zhang, R.; Chen, S. A conjugative plasmid that augments virulence in Klebsiella pneumoniae. Nat. Microbiol. 2019, 4, 2039–2043. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Li, Y.; Bu, K.; Wang, M.; Wang, Z.; Li, R. Antimicrobial Resistance and Genomic Epidemiology of tet(X4)-Bearing Bacteria of Pork Origin in Jiangsu, China. Genes 2023, 14, 36. https://doi.org/10.3390/genes14010036
Li Y, Li Y, Bu K, Wang M, Wang Z, Li R. Antimicrobial Resistance and Genomic Epidemiology of tet(X4)-Bearing Bacteria of Pork Origin in Jiangsu, China. Genes. 2023; 14(1):36. https://doi.org/10.3390/genes14010036
Chicago/Turabian StyleLi, Yuhan, Yan Li, Kefan Bu, Mianzhi Wang, Zhiqiang Wang, and Ruichao Li. 2023. "Antimicrobial Resistance and Genomic Epidemiology of tet(X4)-Bearing Bacteria of Pork Origin in Jiangsu, China" Genes 14, no. 1: 36. https://doi.org/10.3390/genes14010036