
Genome-Wide Identification of GYF-Domain Encoding Genes in Three Brassica Species and Their Expression Responding to Sclerotinia sclerotiorum in Brassica napus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of GYFs
2.2. Gene Structure, Motif Predicjtion, Cis-Element Prediction, Sequence Alignment, and Phylogenetic Analysis
2.3. Synteny Analysis and Chromosome Distribution Visualization of GYFs
2.4. Plant Growth and Infection Treatment
2.5. RNA Extract and RT-qPCR
2.6. RT-qPCR Results Analysis of BnaGYFs Induced by S. Sclerotiorum
3. Results
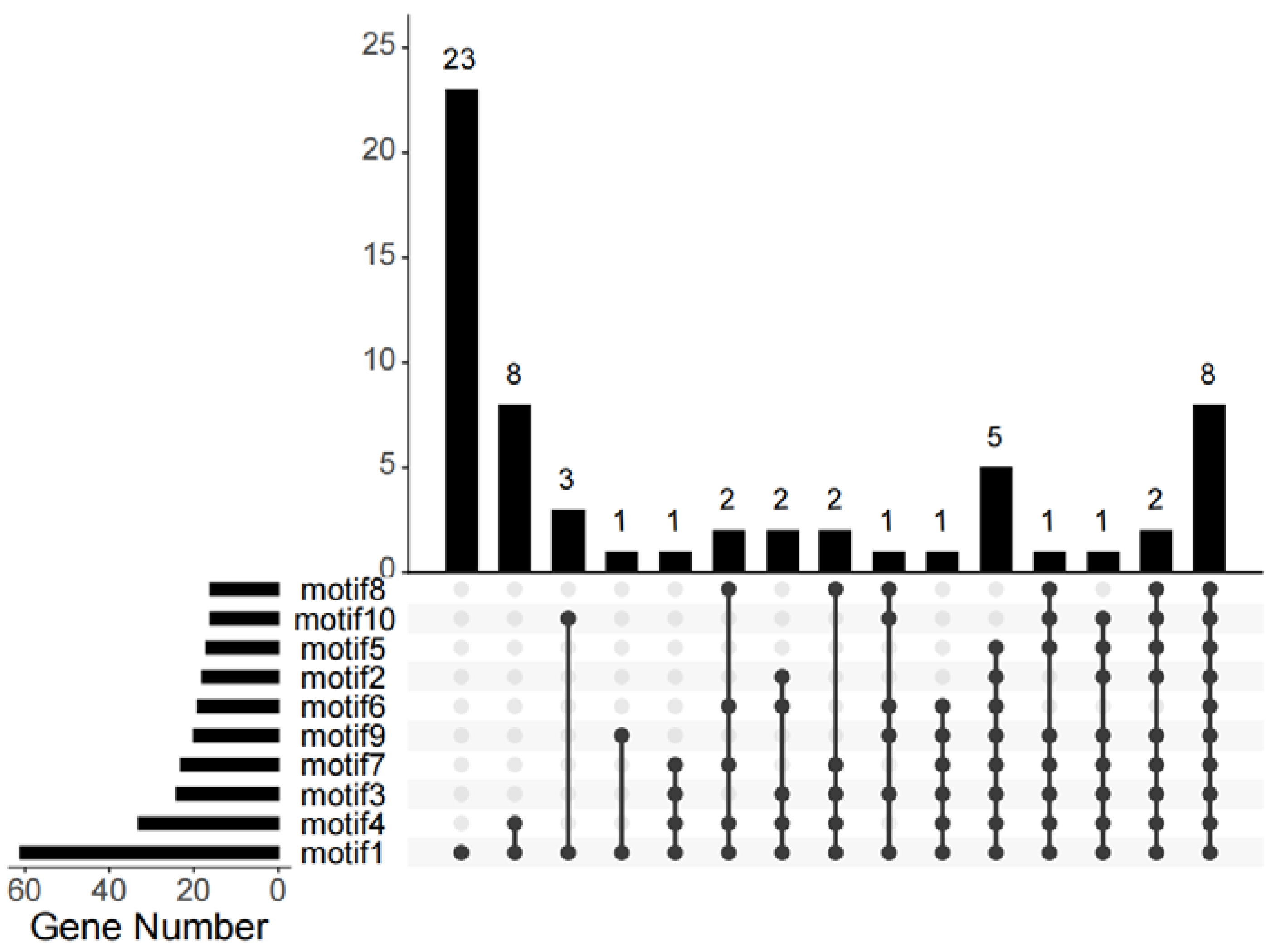
3.1. Identification and Motif Analysis of GYFs
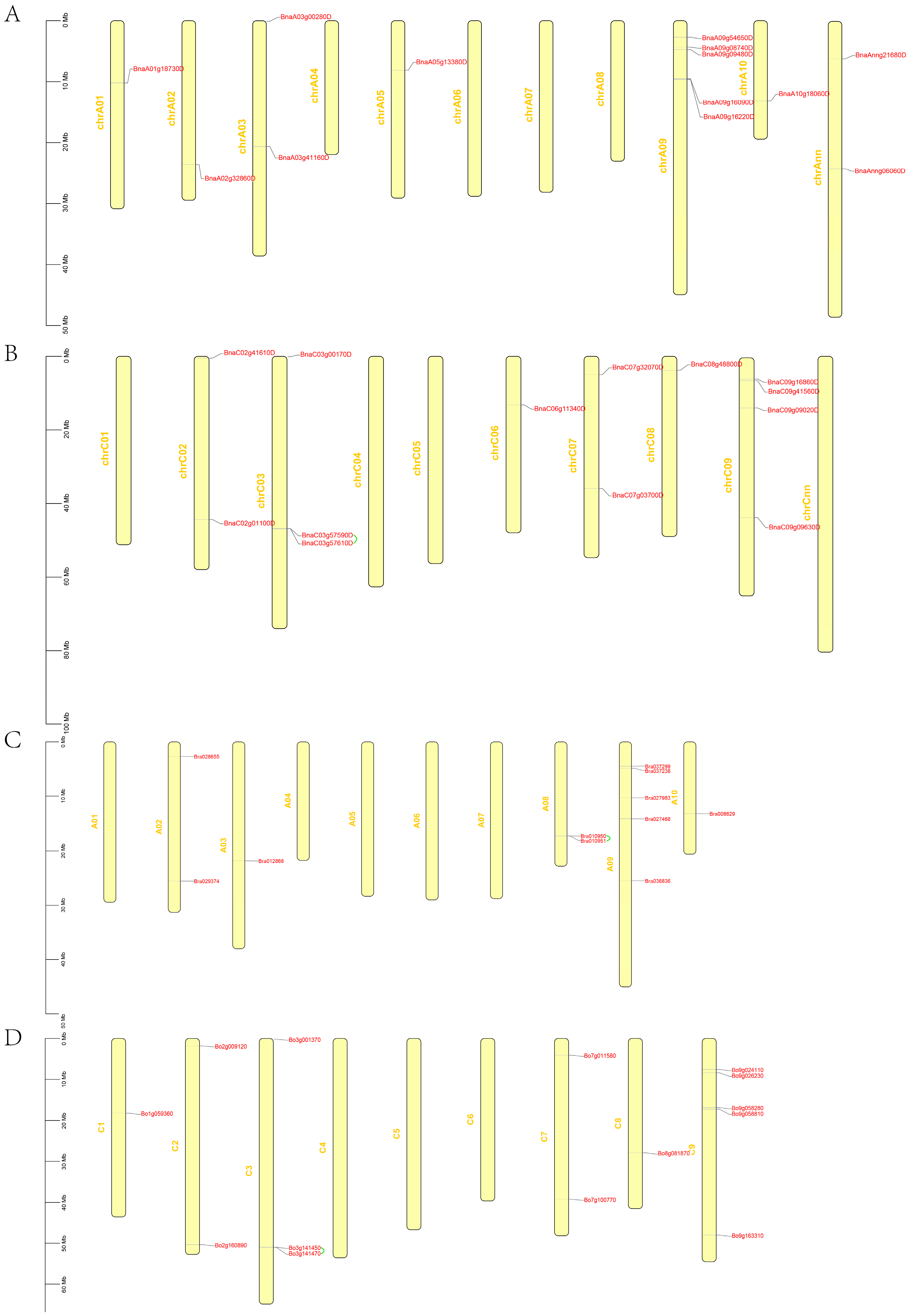
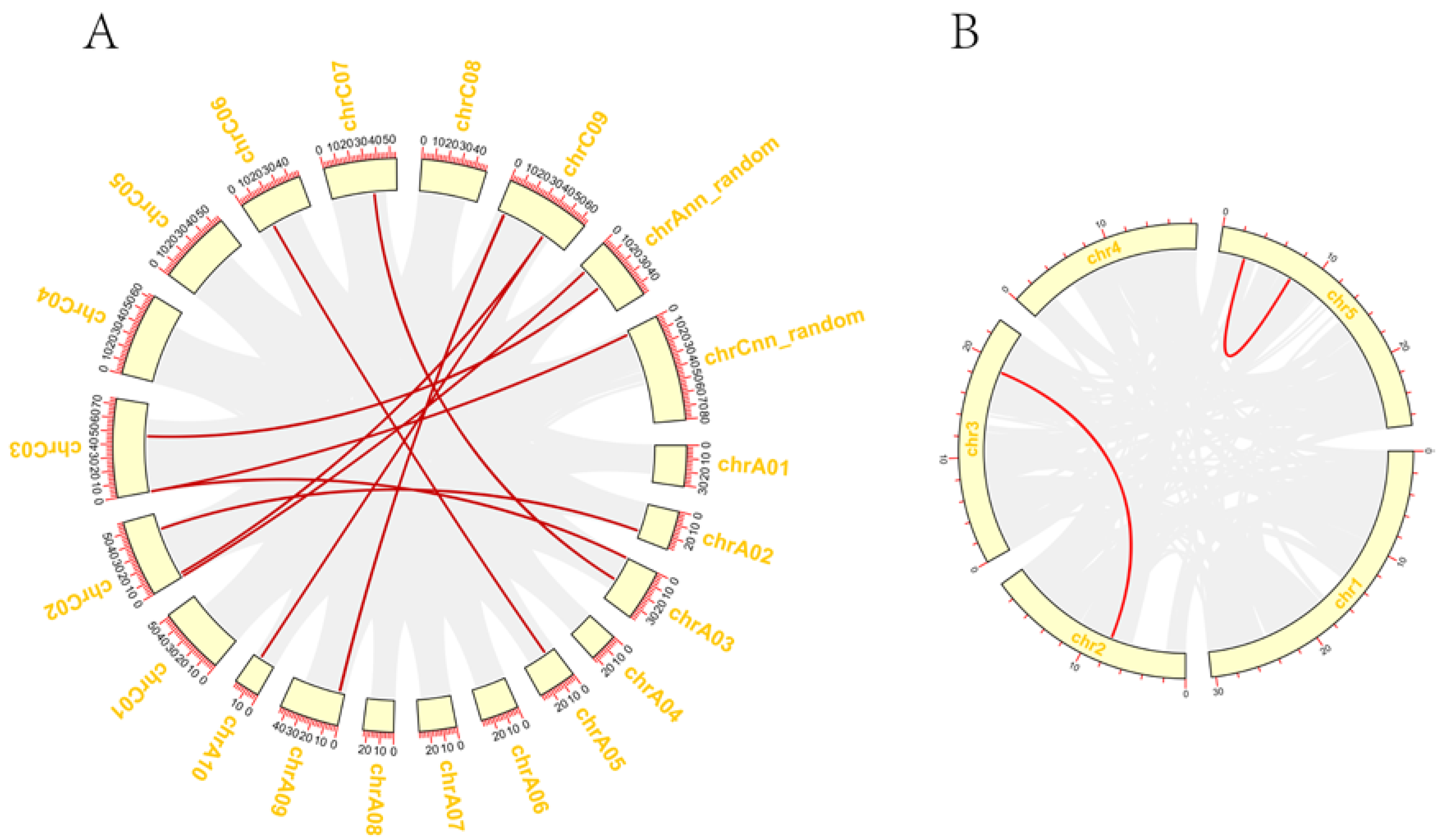
3.2. Chromosomal Localization and Collinearity Analysis of GYFs
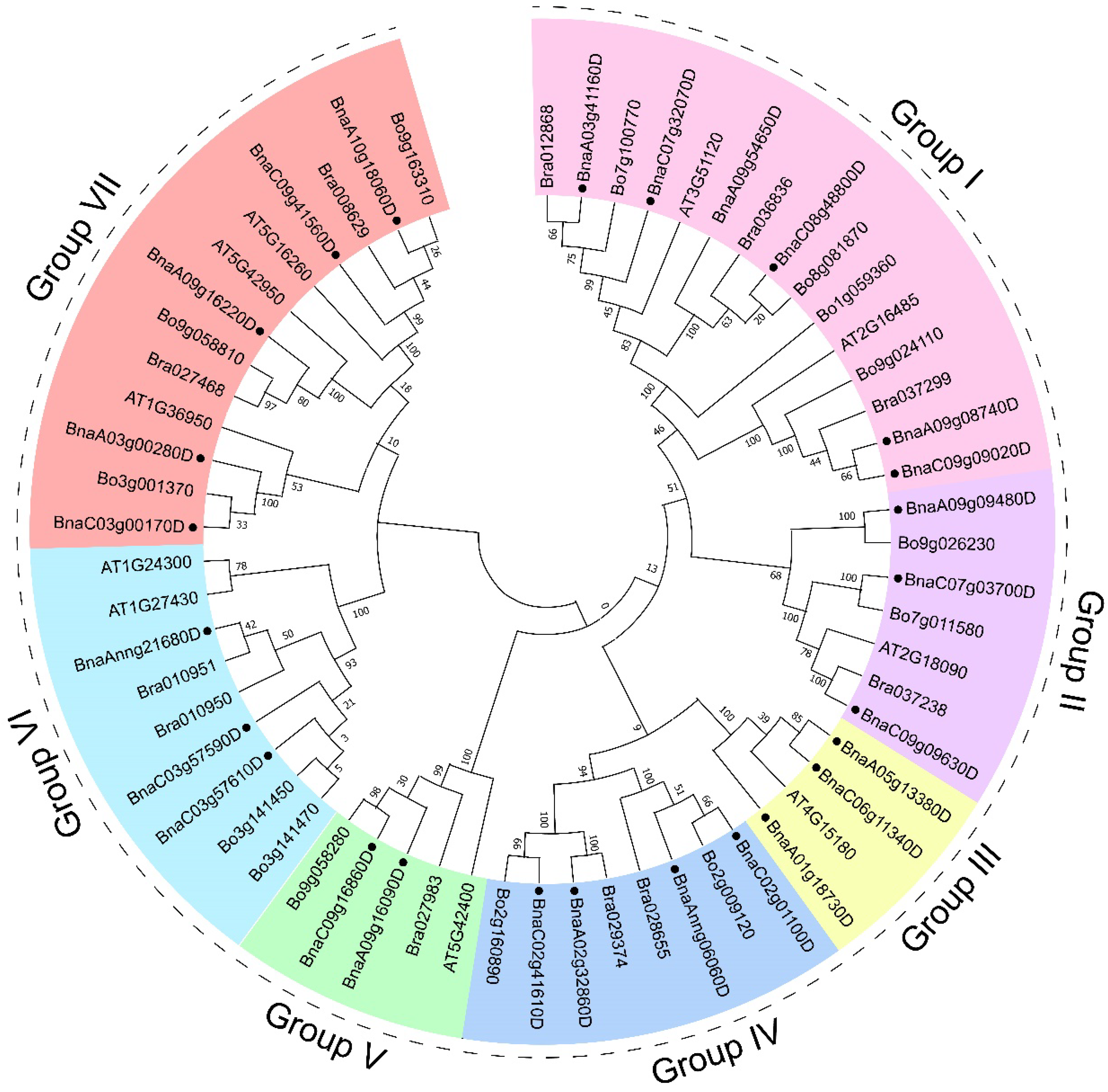
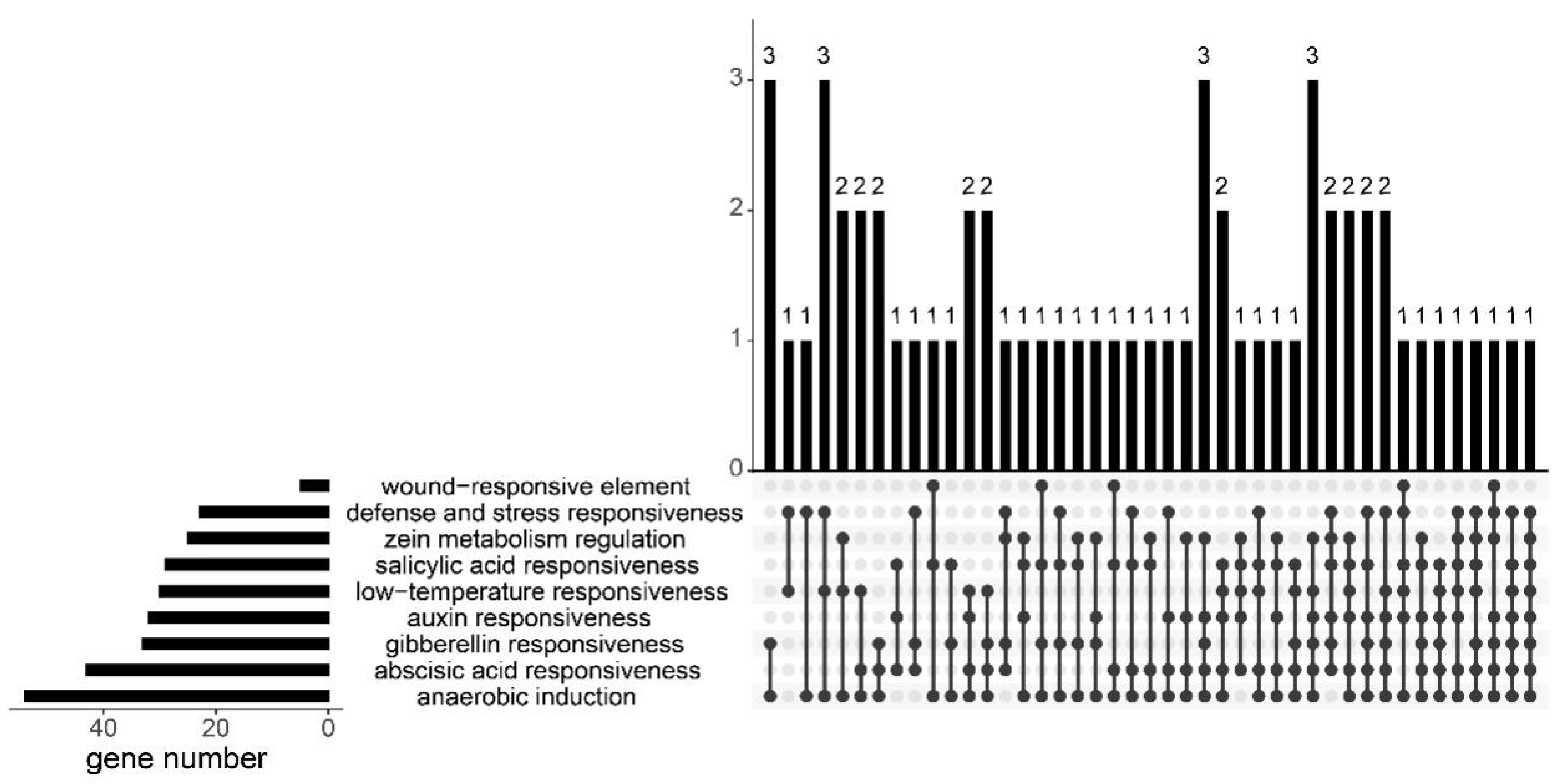
3.3. Phylogenetic and Promoter Cis-Acting Elements Analysis of GYFs
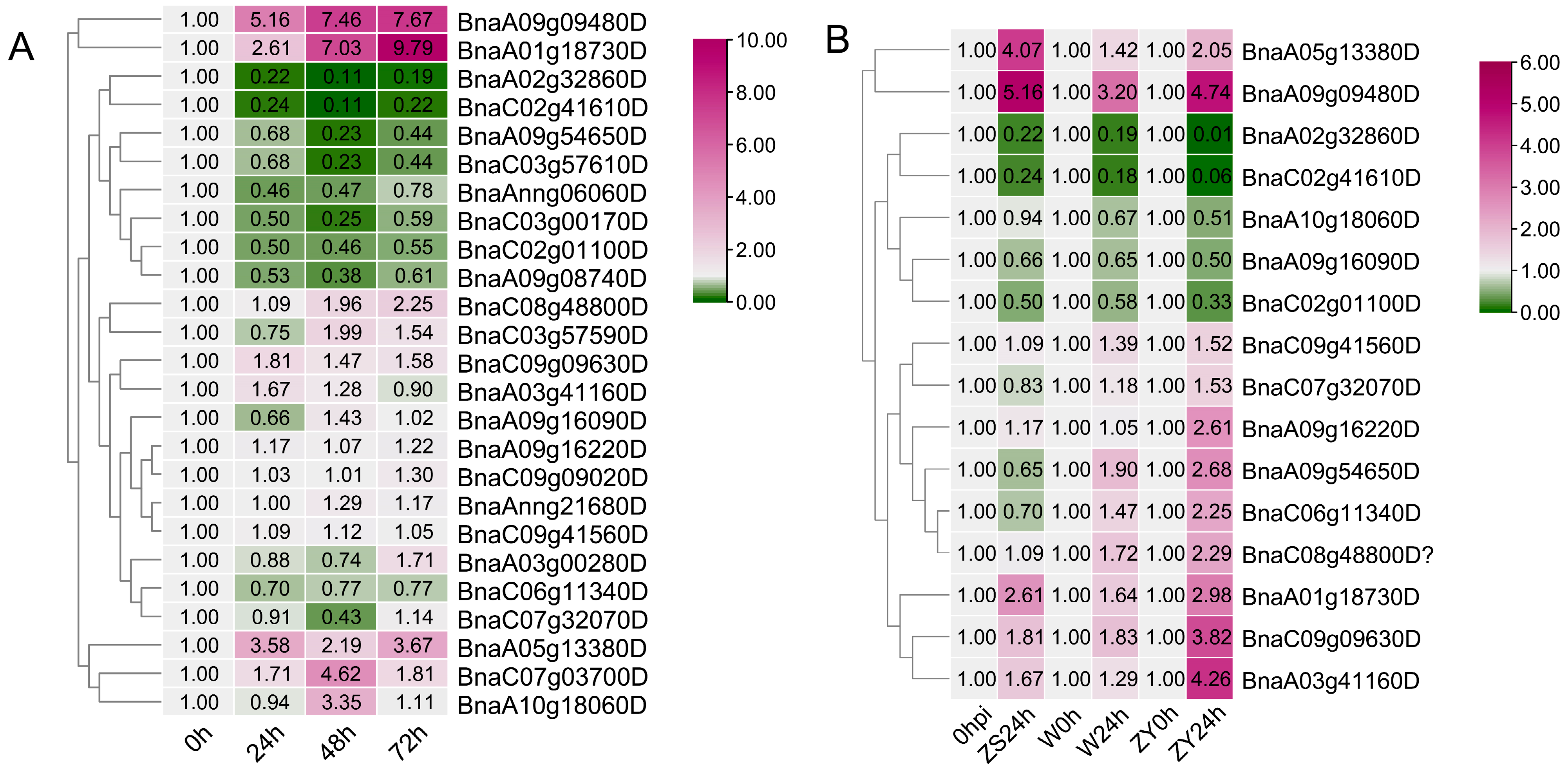
3.4. Expression of BnaGYFs under S. Sclerotiorum Induction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nishizawa, K.; Freund, C.; Li, J.; Wagner, G.; Reinherz, E.L. Identification of a proline-binding motif regulating CD2-triggered T lymphocyte activation. Proc. Natl. Acad. Sci. USA 1998, 95, 14897–14902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kofler, M.M.; Freund, C. The GYF domain. FEBS J. 2006, 273, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Hale, V.A.; Guiney, E.L.; Goldberg, L.Y.; Haduong, J.H.; Kwartler, C.S.; Scangos, K.W.; Goutte, C. Notch signaling is antagonized by SAO-1, a novel GYF-domain protein that interacts with the E3 ubiquitin ligase SEL-10 in Caenorhabditis elegans. Genetics 2012, 190, 1043–1057. [Google Scholar] [CrossRef] [Green Version]
- Ash, M.R.; Faelber, K.; Kosslick, D.; Albert, G.I.; Roske, Y.; Kofler, M.; Schuemann, M.; Krause, E.; Freund, C. Conserved β-hairpin recognition by the GYF Domains of Smy2 and GIGYF2 in mRNA surveillance and vesicular transport complexes. Structure 2010, 18, 944–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, M.; Ler, L.W.; Fabian, M.R.; Siddiqui, N.; Mullin, M.; Henderson, V.C.; Alain, T.; Fonseca, B.D.; Karashchuk, G.; Bennett, C.F.; et al. A novel 4EHP-GIGYF2 translational repressor complex is essential for mammalian development. Mol. Cell. Biol. 2012, 32, 3585–3593. [Google Scholar] [CrossRef] [Green Version]
- Giovannone, B.; Tsiaras, W.G.; de la Monte, S.; Klysik, J.; Lautier, C.; Karashchuk, G.; Goldwurm, S.; Smith, R.J. GIGYF2 gene disruption in mice results in neurodegeneration and altered insulin-like growth factor signaling. Hum. Mol. Genet. 2009, 18, 4629–4639. [Google Scholar] [CrossRef] [Green Version]
- Geiger, J.A.; Carvalho, L.; Campos, I.; Santos, A.C.; Jacinto, A. Hole-in-one mutant phenotypes link EGFR/ERK signaling to epithelial tissue repair in Drosophila. PLoS ONE 2011, 6, e28349. [Google Scholar] [CrossRef] [Green Version]
- Amaya, R.C.C.; Hubbe, P.; Mandel, N.; Béthune, J. 4EHP-independent repression of endogenous mRNAs by the RNA-binding protein GIGYF2. Nucleic Acids Res. 2018, 46, 5792–5808. [Google Scholar]
- Freund, C.; Kühne, R.; Yang, H.; Park, S.; Reinherz, E.L.; Wagner, G. Dynamic interaction of CD2 with the GYF and the SH3 domain of compartmentalized effector molecules. EMBO J. 2002, 21, 5985–5995. [Google Scholar] [CrossRef] [Green Version]
- Pontier, D.; Picart, C.; Roudier, F.; Garcia, D.; Lahmy, S.; Azevedo, J.; Alart, E.; Laudié, M.; Karlowski, W.M.; Cooke, R. NERD, a plant-specific GW protein, defines an additional RNAi-dependent chromatin-based pathway in Arabidopsis. Mol. Cell 2012, 48, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, M.; Neriya, Y.; Keima, T.; Iwabuchi, N.; Koinuma, H.; Hagiwara-Komoda, Y.; Ishikawa, K.; Himeno, M.; Maejima, K.; Yamaji, Y.; et al. EXA1, a GYF domain protein, is responsible for loss-of-susceptibility to plantago asiatica mosaic virus in Arabidopsis thaliana. Plant J. 2016, 88, 120–131. [Google Scholar] [CrossRef]
- Wu, Z.S.; Huang, S.; Zhang, X.B.; Wu, D.; Xia, S.T.; Li, X. Regulation of plant immune receptor accumulation through translational repression by a glycine-tyrosine-phenylalanine (GYF) domain protein. Elife 2017, 6, e23684. [Google Scholar] [CrossRef] [PubMed]
- Matsui, H.; Nomura, Y.; Egusa, M.; Hamada, T.; Hyon, G.S.; Kaminaka, H.; Watanabe, Y.; Ueda, T.; Trujillo, M.; Shirasu, K.; et al. The GYF domain protein PSIG1 dampens the induction of cell death during plant-pathogen interactions. PLoS Genet. 2017, 13, e1007037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheer, H.; de Almeida, C.; Ferrier, E.; Simonnot, Q.; Poirier, L.; Pflieger, D.; Sement, F.M.; Koechler, S.; Piermaria, C.; Krawczyk, P. The TUTase URT1 connects decapping activators and prevents the accumulation of excessively deadenylated mRNAs to avoid siRNA biogenesis. Nat. Commun. 2021, 12, 1298. [Google Scholar] [CrossRef] [PubMed]
- Waterworth, W.M.; Wilson, M.; Wang, D.; Nuhse, T.; Warward, S.; Selley, J.; West, C.E. Phosphoproteomic analysis reveals plant DNA damage signaling pathways with a functional role for histone H2AX phosphorylation in plant growth under genotoxic stress. Plant J. 2019, 100, 1007–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayum, M.A.; Kim, H.T.; Nath, U.K.; Park, J.I.; Kho, K.H.; Cho, Y.G.; Nou, I.S. Research on biotic and abiotic stress related genes exploration and prediction in Brassica rapa and B. oleracea: A review. Plant Breed. Biotechnol. 2016, 4, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.R.; Zaman, R.; Avice, J.C.; Ourry, A.; Kim, T.H. Sulfur use efficiency is a significant determinant of drought stress tolerance in relation to photosynthetic activity in Brassica napus cultivars. Front. Plant Sci. 2016, 7, 459. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.K.; Das, A.K.; Gantait, S.; Kumar, V.; Gurel, E. Applications of carbon nanomaterials in the plant system: A perspective view on the pros and cons. Sci. Total Environ. 2019, 667, 485–499. [Google Scholar] [CrossRef]
- Ding, L.N.; Li, T.; Guo, X.J.; Li, M.; Liu, X.Y.; Cao, J.; Tan, X.L. Sclerotinia stem rot resistance in rapeseed: Recent progress and future prospects. J. Agric. Food Chem. 2021, 69, 2965–2978. [Google Scholar] [CrossRef]
- Boland, G.J.; Hall, R. Index of plant hosts of Sclerotinia sclerotiorum. Can. J. Plant Pathol. 1994, 16, 93–108. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.H.J.; Nelson, B.D. Sclerotinia sclerotiorum (lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, R.D. Pectolytic enzymes of Sclerotinia sclerotiorum and their localization in infected bean. Can. J. Bot. 1976, 54, 2630–2641. [Google Scholar] [CrossRef]
- Riou, C.; Freyssinet, G.; Fevre, M. Production of cell wall-degrading enzymes by the phytopathogenic fungus Sclerotinia sclerotiorum. Appl. Environ. Microbiol. 1991, 57, 1478–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collmer, A.; Keen, N.T. The role of pectic enzymes in plant pathogenesis. Annu. Rev. Phytopathol. 1986, 24, 383–409. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Chalhoub, B.; Denoeud, F.; Liu, S.Y.; Parkin, I.A.P.; Tang, H.B.; Wang, X.Y.; Chiquet, J.; Belcram, H.; Tong, C.B.; Samans, B. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Nagaharu, U. Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. Jpn. J. Bot. 1935, 7, 389–452. [Google Scholar]
- Huang, Y.; Liu, C.; Shen, W.H.; Ruan, Y. Phylogenetic analysis and classification of the Brassica rapa SET-domain protein family. BMC Plant Biol. 2011, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- Alam, I.; Liu, C.C.; Ge, H.L.; Batool, K.; Yang, Y.Q.; Lu, Y.H. Genome wide survey, evolution and expression analysis of PHD finger genes reveal their diverse roles during the development and abiotic stress responses in Brassica rapa L. BMC Genom. 2019, 20, 773. [Google Scholar] [CrossRef] [Green Version]
- Regmi, R.; Newman, T.E.; Kamphuis, L.G.; Derbyshire, M.C. Identification of B. napus small RNAs responsive to infection by a necrotrophic pathogen. BMC Plant Biol. 2021, 21, 366. [Google Scholar] [CrossRef]
- Chen, R.; Tu, Z.; He, C.; Nie, X.; Li, K.; Fei, S.; Song, B.; Nie, B.; Xie, C. Susceptibility factor StEXA1 interacts with StnCBP to facilitate potato virus Y accumulation through the stress granule-dependent RNA regulatory pathway in potato. Hortic. Res. 2022, 9, uhac159. [Google Scholar] [CrossRef]
- Chen, H.; Wang, T.; He, X.; Cai, X.; Lin, R.; Liang, J.; Wu, J.; King, G.; Wang, X. BRAD V3.0: An upgraded Brassicaceae database. Nucleic Acids Res. 2022, 50, D1432–D1441. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, A.; Saqib, S.; Huang, H.; Zaman, W.; Lü, S.; Zhao, H. Genome-wide comparative analysis of long-chain acyl-CoA synthetases (LACSs) gene family: A focus on identification, evolution and expression profiling related to lipid synthesis. Plant Physiol. Biochem. 2021, 161, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar] [PubMed]
- Li, D.H.; Zaman, W.; Lu, J.J.; Niu, Q.Q.; Zhang, X.H.; Ayaz, A.; Saqib, S.; Yang, B.; Zhang, J.X.; Zhao, H.Y.; et al. Natural lupeol level variation among castor accessions and the upregulation of lupeol synthesis in response to light. Ind. Crops Prod. 2023, 192, 116090. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, Q.; Yang, Q.; Liu, H.; Li, Q.; Yi, X.; Cheng, Y.; Guo, L.; Fan, C.; Zhou, Y. Comparative transcriptomic analysis uncovers the complex genetic network for resistance to Sclerotinia sclerotiorum in Brassica napus. Sci. Rep. 2017, 7, 42829. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Z.; Liu, G.H.; Zheng, Y.B.; Wang, X.F.; Yang, Q. Breeding of the Brassica napus cultivar Zhongshuang 9 with high-resistance to Sclerotinia sclerotiorum and dynamics of its important defense enzyme activity. Sci. Agric. Sin. 2004, 37, 23–28. [Google Scholar]
- Li, W.; Lu, J.; Yang, C.; Xia, S. Identification of receptor-like proteins induced by Sclerotinia sclerotiorum in Brassica napus. Front. Plant Sci. 2022, 13, 944763. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Duan, S.; Jian, Y.; Xu, J.; Hu, J.; Zhang, Z.; Lin, T.; Cheng, F.; Li, G. Genome-wide identification and gene expression analysis of the 14-3-3 gene family in potato (Solanum tuberosum L.). BMC Genom. 2022, 23, 811. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Boutte, J.; Maillet, L.; Chaussepied, T.; Letort, S.; Aury, J.M.; Belser, C.; Boideau, F.; Brunet, A.; Coriton, O.; Deniot, G. Genome size variation and comparative genomics reveal intraspecific diversity in Brassica rapa. Front. Plant Sci. 2020, 11, 577536. [Google Scholar] [CrossRef]
- Badet, T.; Voisin, D.; Mbengue, M.; Barascud, M.; Sucher, J.; Sadon, P.; Balagué, C.; Roby, D.; Raffaele, S. Parallel evolution of the POQR prolyl oligo peptidase gene conferring plant quantitative disease resistance. PLoS Genet. 2017, 13, e1007143. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Qin, L.; Lu, J.; Xia, Y.; Tang, X.; Lu, X.; Xia, S. Genome-Wide Identification of GYF-Domain Encoding Genes in Three Brassica Species and Their Expression Responding to Sclerotinia sclerotiorum in Brassica napus. Genes 2023, 14, 224. https://doi.org/10.3390/genes14010224
Zhang X, Qin L, Lu J, Xia Y, Tang X, Lu X, Xia S. Genome-Wide Identification of GYF-Domain Encoding Genes in Three Brassica Species and Their Expression Responding to Sclerotinia sclerotiorum in Brassica napus. Genes. 2023; 14(1):224. https://doi.org/10.3390/genes14010224
Chicago/Turabian StyleZhang, Xiaobo, Lei Qin, Junxing Lu, Yunong Xia, Xianyu Tang, Xun Lu, and Shitou Xia. 2023. "Genome-Wide Identification of GYF-Domain Encoding Genes in Three Brassica Species and Their Expression Responding to Sclerotinia sclerotiorum in Brassica napus" Genes 14, no. 1: 224. https://doi.org/10.3390/genes14010224