
Potential Involvement of LncRNAs in Cardiometabolic Diseases



Center for RNA Medicine, Department of Clinical Medicine, Aalborg University, DK-2450 Copenhagen SV, Denmark
*
Author to whom correspondence should be addressed.
Genes 2023, 14(1), 213; https://doi.org/10.3390/genes14010213
Submission received: 9 December 2022
/
Revised: 10 January 2023
/
Accepted: 11 January 2023
/
Published: 13 January 2023
(This article belongs to the Special Issue Genetics of Cardiovascular Metabolism)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Characterized by cardiovascular disease and diabetes, cardiometabolic diseases are a major cause of mortality around the world. As such, there is an urgent need to understand the pathogenesis of cardiometabolic diseases. Increasing evidence suggests that most of the mammalian genome are transcribed as RNA, but only a few percent of them encode for proteins. All of the RNAs that do not encode for proteins are collectively called non-protein-coding RNAs (ncRNAs). Among these ncRNAs, long ncRNAs (lncRNAs) are considered as missing keys to understand the pathogeneses of various diseases, including cardiometabolic diseases. Given the increased interest in lncRNAs, in this study, we will summarize the latest trend in the lncRNA research from the perspective of cardiometabolism and disease by focusing on the major risk factors of cardiometabolic diseases: obesity, cholesterol, diabetes, and hypertension. Because genetic inheritance is unavoidable in cardiometabolic diseases, we paid special attention to the genetic factors of lncRNAs that may influence cardiometabolic diseases.
1. Introduction
Cardiometabolic diseases (CMD) account for 31% of all global deaths [1]. These life-long and life-threatening diseases are associated with an unhealthy lifestyle (e.g., obesity, physical inactivity, smoking), leading to diabetes mellitus and cardiovascular disease (CVD) [2], and more than 56,000 Danes are diagnosed with CVD annually [3]. In CMD (e.g., arrhythmic disorders, cardiomyopathies, Fabry disease, familial hypercholesterolemia, thoracic aortic aneurysms and dissections [4]), the genetic inheritance is unavoidable. For example, siblings of patients with cardiovascular disease (CVD) have a 40% increase of risk for CVD [5]. Thus, significant efforts have been spent to understand the genetic causes of CMD by genome sequencing the patients suffering from CMD. Through the usages of linkage disequilibrium (LD) and genome-wide association studies (GWAS), a number of loci are found to be associated with CMD [6,7,8,9,10]. Yet, only few are directly in the exons of protein-coding genes, leaving behind the large number of CMD-suspected loci to be unexplained.
The term junk DNA is long gone, and we now know that most of the mammalian genome are transcribed as RNA. Yet only few percent of them correspond to the exons of protein-coding genes, leaving behind the large number of RNAs as non-protein-coding (ncRNAs). These ncRNAs include ribosomal RNAs (rRNAs), transfer RNAs (tRNAs), short ncRNAs (e.g., small nuclear RNAs (snRNAs), small nucleolar RNAs (snoRNAs)), microRNAs (miRNAs), and other longer ncRNAs [11]. Long non-coding RNAs (lncRNAs) are any ncRNAs longer than 200 nucleotides (nt). Because the protein-centered research has not been able to fully elucidate the pathogeneses of CMD nor offer an ultimate cure for CMD, there are growing interests to study lncRNAs in the context of cardiovascular metabolism and CMD. Because lncRNAs occupy a large part of the human genome in comparison to protein-coding genes, increasing evidence suggests that disease-suspected loci based on LD and GWAS are in the loci with lncRNAs [12,13], including the well-known example of the lncRNA, ANRIL [official gene symbol: CDKN2B-AS1 (CDKN2B antisense RNA 1)], which corresponds to the Chr9p21 risk locus for coronary artery disease based on GWAS studies [14,15,16,17,18,19,20,21]. Due to the decreased costs of performing both whole genome and transcriptome sequencing (i.e., DNA sequencing (DNA-seq) and RNA-seq, respectively), many lncRNAs have been identified in the disease-suspected loci, which calls for further investigation on how the mutations in lncRNAs themselves affect the susceptibility of each individual to a particular genetic disorder.
Here, we summarize the recent findings of lncRNAs in cardiovascular metabolism and CMD. Because there are many review articles summarizing lncRNAs in CVD, we will focus our attention on the lncRNAs that are involved in risk factors of CMD; namely, obesity, cholesterol, diabetes, and hypertension. To cope with the growing interest to study genetic causes of CMD, we will pay special attention to lncRNAs that lay within the CMD-suspected loci.
2. Obesity-Associated lncRNAs
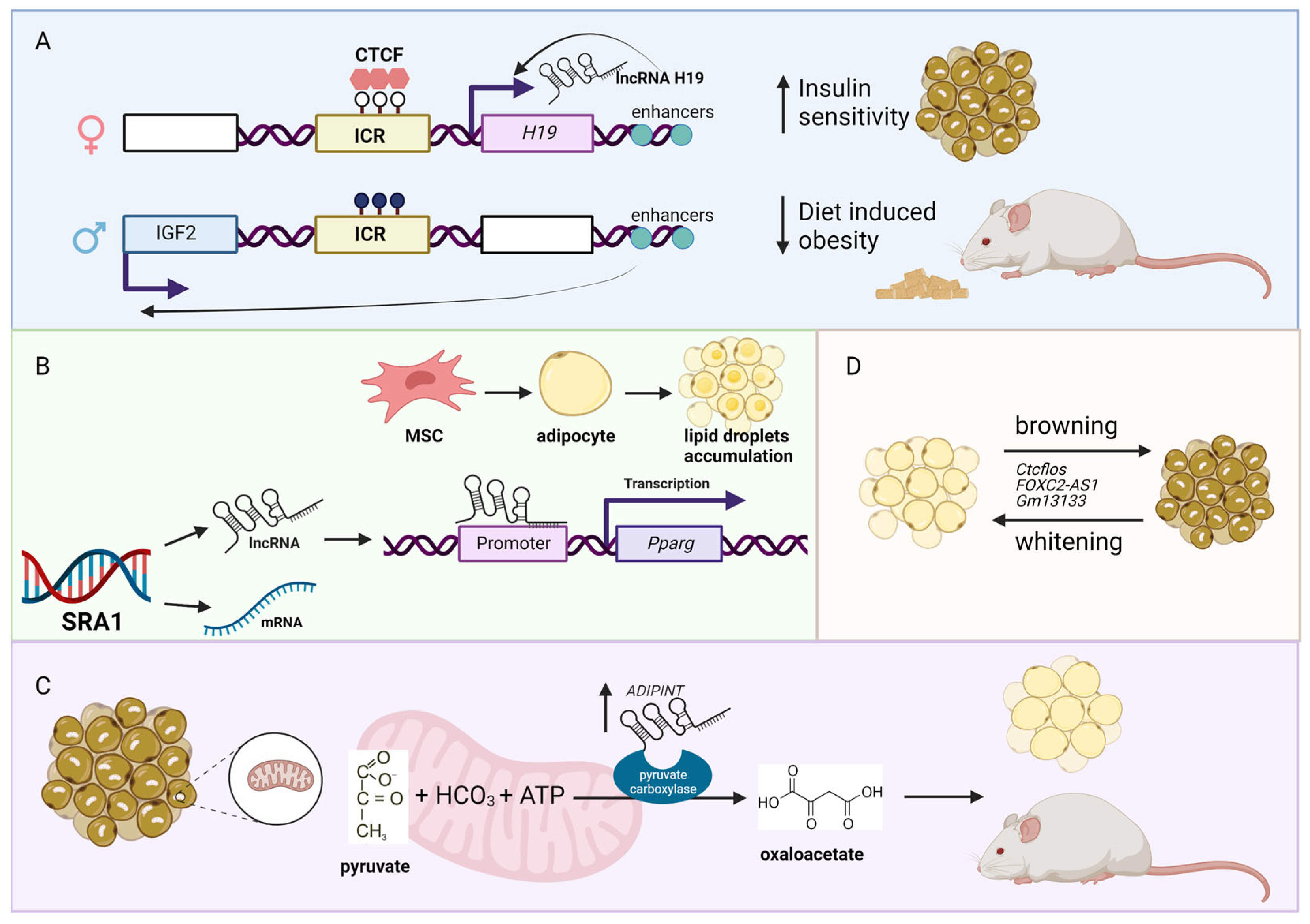
Due to the increased consumption of high energy and calorie food (e.g., fat, oil, and sugar) coupled with physical inactivity, the number of obese people has increased dramatically worldwide in recent years [22,23,24]. Because obesity is an underlying cause and risk factor for many diseases (e.g., type II diabetes, CVD), the interest to study obesity-associated lnRNAs has increased recently. For example, the expression of one of the most well-studied lncRNA, H19 imprinted maternally expressed transcript (H19), is reduced in adipocytes of obese individuals [25]. The functional role of H19 in CMD is well described, which includes negatively regulating hypertrophy [26,27,28] and pyroptosis (a form of cell death initiated by inflammation) [29] in cardiomyocytes. The reduced expression of H19 inversely correlates with body mass index (BMI) in humans. Interestingly, H19 overexpressing mice showed a protective effect against diet-induced obesity and improved insulin sensitivity in brown but not in white adipocytes (Figure 1A). Because BMI is a measure of body fat based on the height and weight of a person, it is used as an indicator of obesity. As such, GWAS and LD studies have been performed to identify BMI-associated loci [30,31]. Not surprisingly, several lncRNAs are shown to be in the BMI-associated loci, such as LncOb rs10487505 variant between BMI and leptin level in pediatric non-alcoholic fatty liver disease [32], lncRNA transcript of the transcription factor 7 like 2 (lncRNA-TCF7L2) between BMI and biopolar disorder [33], MCHR2 antisense RNA 1 (MCHR2-AS1) between BMI and psychiatric patients [34], and the hypermethylation of the promoter of the lncRNA, TSPOAP1, SUPT4H1and RNF43 antisense RNA 1 (TSPOAP1-AS1), associated with obesity in overweight and obese Korean individuals [35]. These are just a few examples of BMI-associated lncRNAs reported to date, which most likely increase as more previously published GWAS data are re-analyzed from the standpoint of lncRNAs.
Obesity is characterized by the increased number of white adipocytes and adipocyte gene expression [36,37]. For example, the whole-body knockout mice of the lncRNA steroid receptor RNA activator 1 (Sra1) are resistant to high fat diet-induced obesity, with decreased epididymal and subcutaneous white adipose tissue (WAT) mass, and increased lean content (i.e., decreased percent of body fat) [38]. Mechanistically, it was shown previously that Sra1 functions as a transcriptional coactivator of peroxisome proliferator-activated receptor gamma (Pparg), which is a master regulator of adipogenesis [39] (Figure 1B). It should be noted that SRA1 encodes for both lncRNA and protein-coding transcripts (https://www.ensembl.org/Homo_sapiens/Gene/Summary?db=core;g=ENSG00000213523;r=5:140537340-140557677; accessed on 11 January 2023). Another example is adipocyte associated pyruvate carboxylase interacting lncRNA (ADIPINT), whose expression is increased in obesity and linked to fat cell size, adipose insulin resistance, and pyruvate carboxylase activity [40]. Mechanistically, ADIPINT binds to pyruvate carboxylase, which is a mitochondrial enzyme that catalyzes the ATP-dependent carboxylation of pyruvate to oxaloacetate in order to regulate lipid metabolism (Figure 1C). As adipocytes can be easily isolated and cultured, there are several screening studies to find lncRNAs associated with adipogenesis identifying hundreds of differentially expressed lncRNAs [41,42,43,44,45,46]. Thus, further functional and mechanistic studies are needed to elucidate the functional importance of these adipose-related lncRNAs.
As mentioned above, there are two types of adipocytes: brown and white adipocytes. While white adipocytes store excessive energy as trigycerides, brown adipocytes have significantly more mitochondria than white adipocytes and burn energy to generate heat [47]. Because of the benefits of having more brown adipocytes over white ones for reducing obesity, the interest to study brown adipose tissue (BAT) has increased in recent years [48]. For example, another well-studied lncRNA, X inactive specific transcript (XIST), regulates brown preadipocytes differentiation by directly binding to CCAAT enhancer binding protein alpha (CEBPA, also known as C/EBPα) [49]. Besides BAT, browning of WAT (also called as adipose tissue browning) is gaining attention as a potential therapeutic target to induce thermogenically active adipocytes in WAT to increase energy expenditure and counteract weight gain [50]. During adipose tissue browning, several lncRNAs have been identified and functionally studied, including CCCTC-binding factor (zinc finger protein)-like, opposite strand (Ctcflos) [51], FOXC2 antisense RNA 1 (FOXC2-AS1) [52], and predicted gene 13133 (Gm13133) [53] (Figure 1D). Thus, lncRNAs might hold keys to reduce obesity, thereby reducing CMD.
3. Cholesterol-Associated lncRNAs
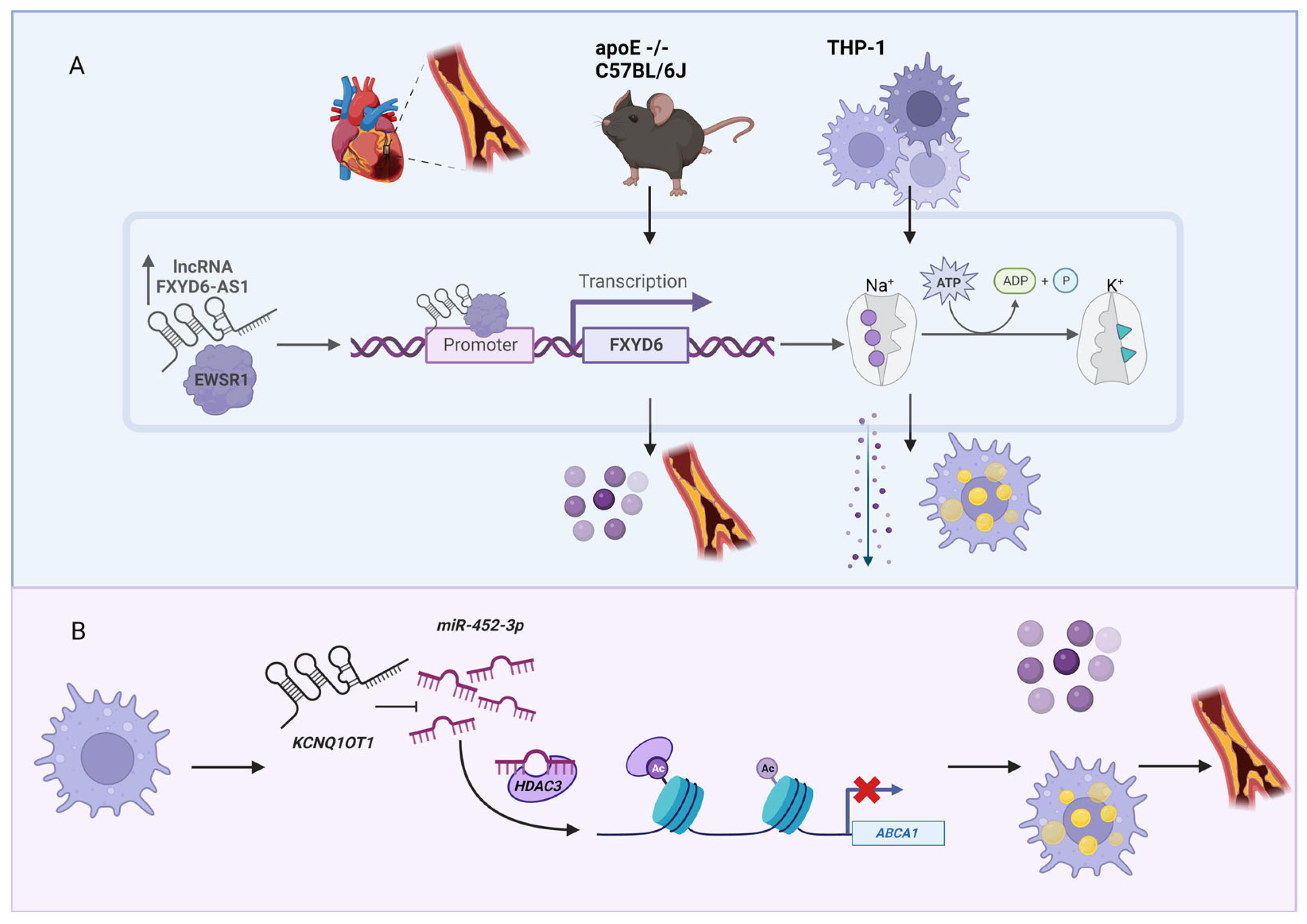
Although cholesterol is needed for our body, an excessive amount of cholesterol in blood leads to coronary artery disease (CAD, also known as atherosclerosis, a type of cardiovascular disease), which is caused by the blockade of arteries by cholesterol and other substances that form atherosclerotic plaques [54]. Because CAD is the third leading cause of mortality worldwide [55], there is a high interest to understand the pathogenesis and potentially find cures for CAD. Besides the most well-known CAD-associated lncRNA, ANRIL, as mentioned in the Introduction section, there are several lncRNAs associated with CAD [56,57]. For example, the expression of the lncRNA FXYD6 antisense RNA 1 (FXYD6-AS1, also known as RP11-728F11.4), is increased in atherosclerosis plaques [58] (Figure 2A). In the cultured monocyte-derived macrophages (THP-1 cells), lentivirus-induced overexpression of FXYD6-AS1 resulted in higher sodium–potassium adenosine triphosphatase (Na+/K+-ATPase) activity, intracellular cholesterol accumulation, and increased proinflammatory cytokine production. These phenotypes were also observed in apolipoprotein E deficient (apoE)−/−mice on a C57BL/6J genetic background fed with a Western diet (a defined mouse model of atherosclerosis as well as non-alcoholic steatohepatitis [59,60]), where overexpression of FXYD6-AS1 increased proinflammatory cytokine production and augmented atherosclerotic lesions. Mechanistically, FXYD6-AS1 binds EWS RNA-binding protein 1 (EWSR1), which binds to the promoter region of FXYD domain-containing ion transport regulator 6 (FXYD6), the modulator of Na+/K+-ATPase. Using the similar in vitro and in vivo experimental systems, another well-known lncRNA, KCNQ1 opposite strand/antisense transcript 1 (KCNQ1OT1), was also shown to be involved in CAD. KCNQ1OT1 functions as a microRNA (miRNA) sponge to sequester miR-452-3p, which binds to histone deacetylase 3 (HDAC3) and thereby inhibits the expression of ATP binding cassette subfamily A member 1 (ABCA1) (Figure 2B). This coordinated pathway in macrophages results in lipid accumulation and accelerates the atherosclerosis development [61].
As exemplified by ANRIL, there are several polymorphisms in lncRNA genes associated with CAD [62,63], including growth arrest specific 5 (GAS5) [64], H19 [65,66,67], HOX transcript antisense RNA (HOTAIR) [68], metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) [69,70,71], and tumor protein p53 pathway corepressor 1 (TP53COR1, also known as lincRNA-p21) [72]. According to the latest annotation provided by the Ensembl database [73] (https://www.ensembl.org/Homo_sapiens/Gene/Summary?db=core;g=ENSG00000251562;r=11:65497688-65506516; accessed on 11 January 2023), there are 17 transcripts (isoforms) of MALAT1. The length of the longest transcript is 8762 nucleotides (nt). As such, a simple search of the NCBI database of genetic variation, dbSNP database [74] (https://www.ncbi.nlm.nih.gov/snp/?term=malat1; accessed on 11 January 2023), indicates that there are 7341 records (dbSNP RefSNP IDs), suggesting that there are many sequence variants within MALAT1 because it is a long transcript. When the term MALAT1 was searched via the ClinVar database [75] (freely available, public archive of human genetic variants and interpretations of their significance to disease) (https://www.ncbi.nlm.nih.gov/clinvar/?term=malat1%5Bgene%5D&redir=gene; accessed on 11 January 2023), 14 records (e.g., ependymoma and glycogen storage disease, type V, Bardet-Biedl syndrome) were found, further suggesting that the lncRNA polymorphisms are associated with various diseases, not only CAD. Not surprisingly, the longer the lncRNA, the more genetic variations are found, which increases the chance of finding associated diseases. To this end, re-analysis of published GWAS studies and their data sets is important to further identify lncRNAs in disease-suspected loci.
4. Diabetes-Associated lncRNAs
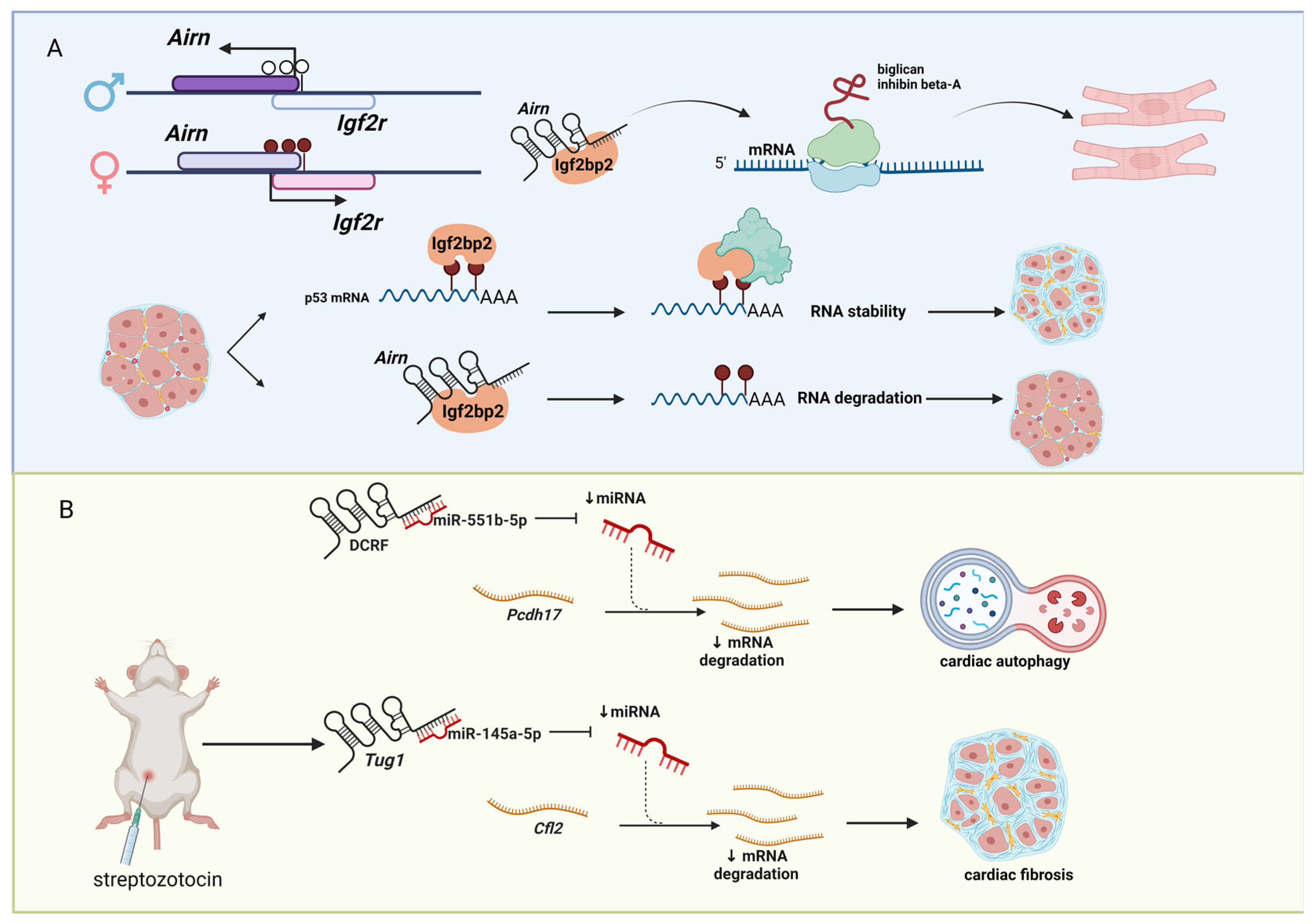
Type II diabetes (T2D) is a chronic disease condition with high blood glucose level caused by the inability of the pancreas to produce enough insulin to regulate glucose level [76,77]. The prevalence of T2D is high, affecting approximately 6.28% of the world’s population [78]. The causes of T2D include obesity, physical inactivity, and genetic factors [79,80,81]. If left untreated, T2D can cause other health problems, including CVD [82], high blood pressure (hypertension) [83], vision loss (diabetic retinopathy) [84], and kidney disease [85]. Thus, intensive T2D research is ongoing to understand the impact of T2D to other health problems, including CMD. Not surprisingly, several lncRNAs are implicated to be involved in the pathogenesis of T2D [86,87,88,89]. For example, the lncRNA, antisense Igf2r RNA (Airn, also known as Air), is a paternally imprinted gene located in antisense orientation to the imprinted, but maternally derived, insulin-like growth factor 2 receptor (Igf2r) protein-coding gene (Figure 3A). We previously showed that Airn binds to the RNA-binding protein, insulin-like growth factor 2 mRNA binding protein 2 (Igf2bp2, also known as IMP2), and controls the translation of messenger RNAs (mRNAs) (e.g., biglycan, inhibin β-A) to regulate the cardiomyocyte resistance to stress [90]. Recently, another group reported that Airn alleviates diabetic cardiac fibrosis by regulating degradation of transformation-related protein 53 (Trp53, also known as p53) mRNA in N6-methyladenosine (m6A) RNA methylation manner because Igf2bp2 is a m6A reader [91]. This study and others clearly link the importance of lncRNAs in pathogenesis of T2D. Furthermore, lncRNAs can be the determinants to understand how RNA metabolisms are regulated through RNA modifications (epitranscriptomics), as we recently reviewed [92].
Diabetic cardiomyopathy (DCM) is a type of CVD caused by diabetes, resulting in contractile dysfunction and leading to heart failure [93]. Several lncRNAs have been identified to be involved in the pathogenesis of DCM (Figure 3B). For example, the lncRNA DCM-related factor (DCRF) functions as a miRNA sponge to sequester miR-551b-5p, which targets protocadherin 17 (Pcdh17) to regulate autophagy in rat cardiomyocytes [94]. Another example is the lncRNA taurine upregulated gene 1 (Tug1). It is up-regulated in the DCM mouse model based on the intraperitoneally injection of streptozotocin (an antibiotic that produces pancreatic islet β-cell destruction [95]), while down-regulation of Tug1 improved cardiac function and myocardial fibrosis in the DCM mice by decreasing the levels of extracellular matrices (e.g., collagens). Mechanistically, Tug1 binds miR-145a-5p, which targets cofilin 2, muscle (Cfl2) [96]. Besides these two lncRNAs, there are other lncRNAs reported to function as miRNA sponges in DCM [97,98,99,100], suggesting a popular mechanism of action of lncRNAs to be investigated. In this regard, several screening studies to identify lncRNAs functioning as miRNA sponges in DCM have been conducted [101,102,103]. However, it should be noted that many lncRNAs are lowly expressed compared to mRNAs and miRNAs. Thus, it is advised that researchers investigating lncRNAs as miRNA sponges to report the copy numbers of lncRNAs and their target miRNAs to be sure that it is possible for the target lncRNA to be abundantly present in a target cell compared to the available miRNA in the corresponding cell to sequester the target miRNA, which gives rise to the phenotypes that are observed in their experimental systems.
5. Hypertension-Associated lncRNAs
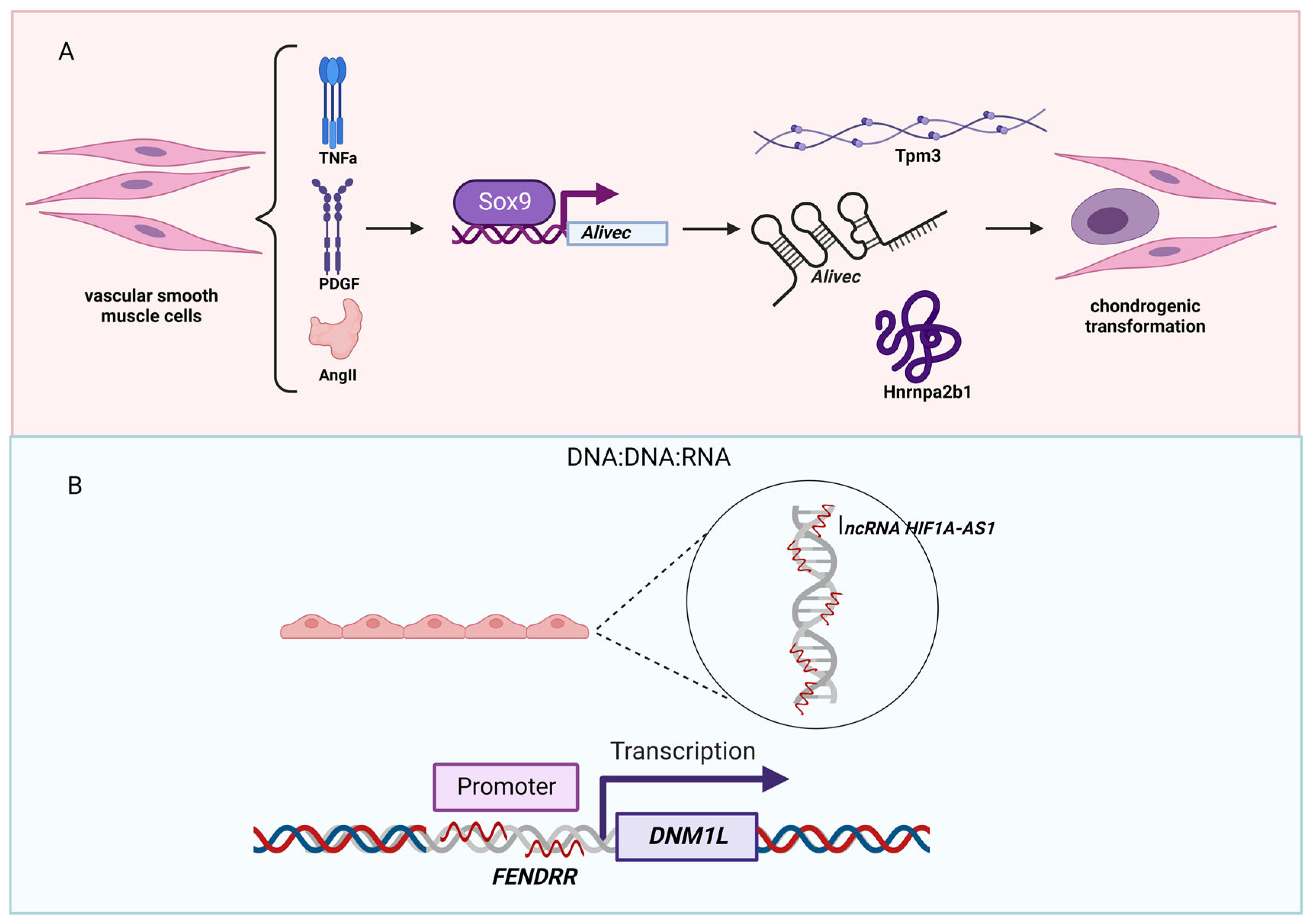
Hypertension (high blood pressure) is a condition in which blood pressure is higher than normal. According to the current guideline by the American College of Cardiology/American Heart Association, hypertension is defined as a cut point of systolic blood pressure (SBP) ≥130 mm Hg and/or diastolic blood pressure (DBP) ≥80 mm Hg [104]. As the risk of hypertension increases due to the underlying conditions (e.g., diabetes and obesity), it is no surprise that several lncRNAs are identified to be linked to hypertension. For example, angiotensin II (AngII)-induced lncRNA Alivec [Angiotensin II-induced lncRNA in vascular smooth muscle cells (VSMCs) eliciting chondrogenic phenotype] is shown to be induced by the growth factors AngII and platelet-derived growth factor (PDGF) and the inflammatory cytokine tumor necrosis factor α (TNF-α) via the action of the transcription factor SRY-box transcription factor 9 (Sox9) in rat VSMCs [105] (Figure 4A). Mechanistically, Alivec binds tropomyosin 3 (Tpm3) and heterogeneous nuclear ribonucleoprotein A2/B1 (Hnrnpa2b1) to regulate chondrogenic transformation of VSMCs implicated in vascular dysfunction and hypertension.
Pulmonary hypertension is caused by high pressure in the blood vessels from the heart to the lungs, which affects both the lungs and the heart [106,107]. It is characterized by pathological changes to signaling pathways (e.g., endothelin, nitric oxide, prostacyclin, transforming growth factor β (TGFB) signaling pathways) [108,109], and a number of lncRNAs have been identified to play key roles in these signaling pathways [110,111,112,113]. Besides binding to miRNAs and RNA-binding proteins, lncRNAs can interact with DNA to regulate the binding of epigenetic and/or transcription factors to the genome, thereby, regulating epitranscriptomic and/or transcriptomic changes (Figure 4B). Increasing evidence suggests that lncRNAs interact with DNA by forming triplex structures [114,115]. Recently, an interesting study was published. In this study, the authors conducted a functional screening of DNA:DNA:RNA triplex-forming lncRNA in human endothelial cells to identify the lncRNA HIF1A antisense RNA 1 (HIF1A-AS1) [116]. HIF1A-AS1 is down-regulated in patients suffering from pulmonary hypertension. Mechanistically, HIF1A-AS1 acts as an adapter for the repressive human silencing hub complex (HUSH). Originally identified as an essential lncRNA gene for the normal development of the heart and body wall in mice [117], FOXF1 adjacent non-coding developmental regulatory RNA (FENDRR) is recently shown to act in a similar mechanism as HIF1A-AS1 by forming triplex structure in the promoter region of dynamin 1 like (DNM1L, also known as DRP1) [118]. These two studies exemplified how the field of lncRNAs has moved from simply identifying the binding partners (e.g., DNA, RNA, and proteins) to detailed mechanistic studies. Although it has not been discussed in these original studies, an open question is whether in such genomic regions that lncRNAs show any disease-specific or -associated mutations or not. It is speculated that bioinformatic analysis by combining genomic mutation data and the binding sites of lncRNAs might reveal further involvement of lncRNAs in disease pathogeneses, including pulmonary hypertension.
6. Conclusions
As summarized above, lncRNAs can be the key to unravel the pathogeneses of CMD, which may lead to the potential cures for CMD [119]. As increasing evidence suggests that genetic factors of CMD are important, understanding how single nucleotide polymorphisms (SNPs) in lncRNAs are linked to various diseases, including CMD, is urgently needed. For this purpose, the readers are advised to explore lncRNA polymorphisms associated with diseases through the NHGRI-EBI Catalog of human genome-wide association studies (https://www.ebi.ac.uk/gwas/search?query=BMI) (accessed on 5 December 2022). Although there are several clinical trials underway to investigate the correlation of lncRNA expression to different diseases as potential diagnostic biomarkers [120,121,122,123], the therapeutic approach to use lncRNAs is still not in any clinical trials [119]. Thus, the further mechanistic investigation of each lncRNA is necessary to advance the lncRNA therapeutics in near future.
Author Contributions
M.I. and S.U. wrote the manuscript, generated figures, and approved the final version of this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yang, T.; Yi, J.; He, Y.; Zhang, J.; Li, X.; Ke, S.; Xia, L.; Liu, L. Associations of Dietary Fats with All-Cause Mortality and Cardiovascular Disease Mortality among Patients with Cardiometabolic Disease. Nutrients 2022, 14, 3608. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.; Menche, J. The regulatory network architecture of cardiometabolic diseases. Nat. Genet. 2022, 54, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Association, T.H. HjerteTal.dk. Available online: www.hjertetal.dk (accessed on 9 December 2022).
- Musunuru, K.; Hershberger, R.E.; Day, S.M.; Klinedinst, N.J.; Landstrom, A.P.; Parikh, V.N.; Prakash, S.; Semsarian, C.; Sturm, A.C.; American Heart Association Council on Genomic and Precision Medicine; et al. Genetic Testing for Inherited Cardiovascular Diseases: A Scientific Statement From the American Heart Association. Circ. Genom. Precis. Med. 2020, 13, e000067. [Google Scholar] [CrossRef]
- Kolber, M.R.; Scrimshaw, C. Family history of cardiovascular disease. Can. Fam. Physician 2014, 60, 1016. [Google Scholar] [PubMed]
- Arvanitis, M.; Tampakakis, E.; Zhang, Y.; Wang, W.; Auton, A.; 23andMe Research Team; Dutta, D.; Glavaris, S.; Keramati, A.; Chatterjee, N.; et al. Genome-wide association and multi-omic analyses reveal ACTN2 as a gene linked to heart failure. Nat. Commun. 2020, 11, 1122. [Google Scholar] [CrossRef] [Green Version]
- Benn, M.; Nordestgaard, B.G. From genome-wide association studies to Mendelian randomization: Novel opportunities for understanding cardiovascular disease causality, pathogenesis, prevention, and treatment. Cardiovasc. Res. 2018, 114, 1192–1208. [Google Scholar] [CrossRef] [PubMed]
- Kessler, T.; Vilne, B.; Schunkert, H. The impact of genome-wide association studies on the pathophysiology and therapy of cardiovascular disease. EMBO Mol. Med. 2016, 8, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Grow, M.A.; Pallaud, C.; Klitz, W.; Erlich, H.A.; Visvikis, S.; Chen, J.J.; Pullinger, C.R.; Malloy, M.J.; Siest, G.; et al. A multilocus genotyping assay for candidate markers of cardiovascular disease risk. Genome Res. 1999, 9, 936–949. [Google Scholar] [CrossRef] [Green Version]
- Aragam, K.G.; Jiang, T.; Goel, A.; Kanoni, S.; Wolford, B.N.; Atri, D.S.; Weeks, E.M.; Wang, M.; Hindy, G.; Zhou, W.; et al. Discovery and systematic characterization of risk variants and genes for coronary artery disease in over a million participants. Nat. Genet. 2022, 54, 1803–1815. [Google Scholar] [CrossRef] [PubMed]
- Uchida, S.; Adams, J.C. Physiological roles of non-coding RNAs. Am. J. Physiol. Cell Physiol. 2019, 317, C1–C2. [Google Scholar] [CrossRef]
- Nurnberg, S.T.; Zhang, H.; Hand, N.J.; Bauer, R.C.; Saleheen, D.; Reilly, M.P.; Rader, D.J. From Loci to Biology: Functional Genomics of Genome-Wide Association for Coronary Disease. Circ. Res. 2016, 118, 586–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirza, A.H.; Kaur, S.; Brorsson, C.A.; Pociot, F. Effects of GWAS-associated genetic variants on lncRNAs within IBD and T1D candidate loci. PLoS ONE 2014, 9, e105723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holdt, L.M.; Teupser, D. Long Noncoding RNA ANRIL: Lnc-ing Genetic Variation at the Chromosome 9p21 Locus to Molecular Mechanisms of Atherosclerosis. Front. Cardiovasc. Med. 2018, 5, 145. [Google Scholar] [CrossRef] [PubMed]
- Holdt, L.M.; Beutner, F.; Scholz, M.; Gielen, S.; Gabel, G.; Bergert, H.; Schuler, G.; Thiery, J.; Teupser, D. ANRIL expression is associated with atherosclerosis risk at chromosome 9p21. Arter. Thromb. Vasc. Biol. 2010, 30, 620–627. [Google Scholar] [CrossRef] [Green Version]
- Matarin, M.; Brown, W.M.; Singleton, A.; Hardy, J.A.; Meschia, J.F. Whole genome analyses suggest ischemic stroke and heart disease share an association with polymorphisms on chromosome 9p21. Stroke 2008, 39, 1586–1589. [Google Scholar] [CrossRef]
- Larson, M.G.; Atwood, L.D.; Benjamin, E.J.; Cupples, L.A.; D’Agostino, R.B., Sr.; Fox, C.S.; Govindaraju, D.R.; Guo, C.Y.; Heard-Costa, N.L.; Hwang, S.J.; et al. Framingham Heart Study 100K project: Genome-wide associations for cardiovascular disease outcomes. BMC Med. Genet. 2007, 8, S5. [Google Scholar] [CrossRef]
- Samani, N.J.; Erdmann, J.; Hall, A.S.; Hengstenberg, C.; Mangino, M.; Mayer, B.; Dixon, R.J.; Meitinger, T.; Braund, P.; Wichmann, H.E.; et al. Genomewide association analysis of coronary artery disease. N. Engl. J. Med. 2007, 357, 443–453. [Google Scholar] [CrossRef] [Green Version]
- The Wellcome Trust Case Control Consortium. Genome-wide association study of 14,000 cases of seven common diseases and 3000 shared controls. Nature 2007, 447, 661–678. [Google Scholar] [CrossRef] [Green Version]
- McPherson, R.; Pertsemlidis, A.; Kavaslar, N.; Stewart, A.; Roberts, R.; Cox, D.R.; Hinds, D.A.; Pennacchio, L.A.; Tybjaerg-Hansen, A.; Folsom, A.R.; et al. A common allele on chromosome 9 associated with coronary heart disease. Science 2007, 316, 1488–1491. [Google Scholar] [CrossRef] [Green Version]
- Helgadottir, A.; Thorleifsson, G.; Manolescu, A.; Gretarsdottir, S.; Blondal, T.; Jonasdottir, A.; Jonasdottir, A.; Sigurdsson, A.; Baker, A.; Palsson, A.; et al. A common variant on chromosome 9p21 affects the risk of myocardial infarction. Science 2007, 316, 1491–1493. [Google Scholar] [CrossRef]
- Fox, A.; Feng, W.; Asal, V. What is driving global obesity trends? Globalization or “modernization”? Glob. Health 2019, 15, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romieu, I.; Dossus, L.; Barquera, S.; Blottiere, H.M.; Franks, P.W.; Gunter, M.; Hwalla, N.; Hursting, S.D.; Leitzmann, M.; Margetts, B.; et al. Energy balance and obesity: What are the main drivers? Cancer Causes Control 2017, 28, 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shook, R.P.; Blair, S.N.; Duperly, J.; Hand, G.A.; Matsudo, S.M.; Slavin, J.L. What is Causing the Worldwide Rise in Body Weight? Eur. Endocrinol. 2014, 10, 136–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, E.; Dhaouadi, I.; Gaziano, I.; Oliverio, M.; Klemm, P.; Awazawa, M.; Mitterer, G.; Fernandez-Rebollo, E.; Pradas-Juni, M.; Wagner, W.; et al. LincRNA H19 protects from dietary obesity by constraining expression of monoallelic genes in brown fat. Nat. Commun. 2018, 9, 3622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viereck, J.; Buhrke, A.; Foinquinos, A.; Chatterjee, S.; Kleeberger, J.A.; Xiao, K.; Janssen-Peters, H.; Batkai, S.; Ramanujam, D.; Kraft, T.; et al. Targeting muscle-enriched long non-coding RNA H19 reverses pathological cardiac hypertrophy. Eur. Heart J. 2020, 41, 3462–3474. [Google Scholar] [CrossRef]
- Hobuss, L.; Foinquinos, A.; Jung, M.; Kenneweg, F.; Xiao, K.; Wang, Y.; Zimmer, K.; Remke, J.; Just, A.; Nowak, J.; et al. Pleiotropic cardiac functions controlled by ischemia-induced lncRNA H19. J. Mol. Cell. Cardiol. 2020, 146, 43–59. [Google Scholar] [CrossRef]
- Liu, L.; An, X.; Li, Z.; Song, Y.; Li, L.; Zuo, S.; Liu, N.; Yang, G.; Wang, H.; Cheng, X.; et al. The H19 long noncoding RNA is a novel negative regulator of cardiomyocyte hypertrophy. Cardiovasc. Res. 2016, 111, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Dong, B.; Chen, M.; Yao, C. LncRNA H19 suppresses pyroptosis of cardiomyocytes to attenuate myocardial infarction in a PBX3/CYP1B1-dependent manner. Mol. Cell. Biochem. 2021, 476, 1387–1400. [Google Scholar] [CrossRef]
- Yengo, L.; Sidorenko, J.; Kemper, K.E.; Zheng, Z.; Wood, A.R.; Weedon, M.N.; Frayling, T.M.; Hirschhorn, J.; Yang, J.; Visscher, P.M.; et al. Meta-analysis of genome-wide association studies for height and body mass index in approximately 700,000 individuals of European ancestry. Hum. Mol. Genet. 2018, 27, 3641–3649. [Google Scholar] [CrossRef]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef]
- Manco, M.; Crudele, A.; Mosca, A.; Caccamo, R.; Braghini, M.R.; De Vito, R.; Alterio, A.; Pizzolante, F.; De Peppo, F.; Alisi, A. LncOb rs10487505 variant is associated with leptin levels in pediatric non-alcoholic fatty liver disease. Pediatr. Res. 2022, 92, 1737–1743. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Nguyen, T.T.L.; Gao, H.; Huang, H.; Kim, D.C.; Sharp, B.; Ye, Z.; Lee, J.H.; Coombes, B.J.; Ordog, T.; et al. TCF7L2 lncRNA: A link between bipolar disorder and body mass index through glucocorticoid signaling. Mol. Psychiatry 2021, 26, 7454–7464. [Google Scholar] [CrossRef] [PubMed]
- Delacretaz, A.; Preisig, M.; Vandenberghe, F.; Saigi Morgui, N.; Quteineh, L.; Choong, E.; Gholam-Rezaee, M.; Kutalik, Z.; Magistretti, P.; Aubry, J.M.; et al. Influence of MCHR2 and MCHR2-AS1 Genetic Polymorphisms on Body Mass Index in Psychiatric Patients and In Population-Based Subjects with Present or Past Atypical Depression. PLoS ONE 2015, 10, e0139155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, N.H.; Cha, M.H.; Kim, M.S. Hypermethylation of the TSPOAP1-AS1 Promoter May Be Associated with Obesity in Overweight/Obese Korean Subjects. Int. J. Mol. Sci. 2020, 21, 3307. [Google Scholar] [CrossRef]
- Sam, S.; Mazzone, T. Adipose tissue changes in obesity and the impact on metabolic function. Transl. Res. 2014, 164, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Attie, A.D.; Scherer, P.E. Adipocyte metabolism and obesity. J. Lipid Res. 2009, 50, S395–S399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Sheng, L.; Miao, H.; Saunders, T.L.; MacDougald, O.A.; Koenig, R.J.; Xu, B. SRA gene knockout protects against diet-induced obesity and improves glucose tolerance. J. Biol. Chem. 2014, 289, 13000–13009. [Google Scholar] [CrossRef] [Green Version]
- Lanz, R.B.; McKenna, N.J.; Onate, S.A.; Albrecht, U.; Wong, J.; Tsai, S.Y.; Tsai, M.J.; O’Malley, B.W. A steroid receptor coactivator, SRA, functions as an RNA and is present in an SRC-1 complex. Cell 1999, 97, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Kerr, A.G.; Wang, Z.; Wang, N.; Kwok, K.H.M.; Jalkanen, J.; Ludzki, A.; Lecoutre, S.; Langin, D.; Bergo, M.O.; Dahlman, I.; et al. The long noncoding RNA ADIPINT regulates human adipocyte metabolism via pyruvate carboxylase. Nat. Commun. 2022, 13, 2958. [Google Scholar] [CrossRef]
- Alvarez-Dominguez, J.R.; Winther, S.; Hansen, J.B.; Lodish, H.F.; Knoll, M. An adipose lncRAP2-Igf2bp2 complex enhances adipogenesis and energy expenditure by stabilizing target mRNAs. iScience 2022, 25, 103680. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, J.; Zhou, X.; Hu, S.; Ge, L.; Sun, J.; Li, P.; Long, K.; Jin, L.; Tang, Q.; et al. Comprehensive Analysis of mRNA and lncRNA Transcriptomes Reveals the Differentially Hypoxic Response of Preadipocytes During Adipogenesis. Front. Genet. 2020, 11, 845. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Lim, Y.C.; Chia, S.Y.; Walet, A.C.E.; Xu, S.; Lo, K.A.; Zhao, Y.; Zhu, D.; Shan, Z.; Chen, Q.; et al. De novo reconstruction of human adipose transcriptome reveals conserved lncRNAs as regulators of brown adipogenesis. Nat. Commun. 2018, 9, 1329. [Google Scholar] [CrossRef] [Green Version]
- Lo, K.A.; Huang, S.; Walet, A.C.E.; Zhang, Z.C.; Leow, M.K.; Liu, M.; Sun, L. Adipocyte Long-Noncoding RNA Transcriptome Analysis of Obese Mice Identified Lnc-Leptin, Which Regulates Leptin. Diabetes 2018, 67, 1045–1056. [Google Scholar] [CrossRef] [Green Version]
- Bai, Z.; Chai, X.R.; Yoon, M.J.; Kim, H.J.; Lo, K.A.; Zhang, Z.C.; Xu, D.; Siang, D.T.C.; Walet, A.C.E.; Xu, S.H.; et al. Dynamic transcriptome changes during adipose tissue energy expenditure reveal critical roles for long noncoding RNA regulators. PLoS Biol. 2017, 15, e2002176. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Goff, L.A.; Trapnell, C.; Alexander, R.; Lo, K.A.; Hacisuleyman, E.; Sauvageau, M.; Tazon-Vega, B.; Kelley, D.R.; Hendrickson, D.G.; et al. Long noncoding RNAs regulate adipogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 3387–3392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saely, C.H.; Geiger, K.; Drexel, H. Brown versus white adipose tissue: A mini-review. Gerontology 2012, 58, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Samuelson, I.; Vidal-Puig, A. Studying Brown Adipose Tissue in a Human in vitro Context. Front. Endocrinol. 2020, 11, 629. [Google Scholar] [CrossRef]
- Wu, C.; Fang, S.; Zhang, H.; Li, X.; Du, Y.; Zhang, Y.; Lin, X.; Wang, L.; Ma, X.; Xue, Y.; et al. Long noncoding RNA XIST regulates brown preadipocytes differentiation and combats high-fat diet induced obesity by targeting C/EBPalpha. Mol. Med. 2022, 28, 6. [Google Scholar] [CrossRef]
- Herz, C.T.; Kiefer, F.W. Adipose tissue browning in mice and humans. J. Endocrinol. 2019, 241, R97–R109. [Google Scholar] [CrossRef]
- Gupta, A.; Shamsi, F.; Altemose, N.; Dorlhiac, G.F.; Cypess, A.M.; White, A.P.; Yosef, N.; Patti, M.E.; Tseng, Y.H.; Streets, A. Characterization of transcript enrichment and detection bias in single-nucleus RNA-seq for mapping of distinct human adipocyte lineages. Genome Res. 2022, 32, 242–257. [Google Scholar] [CrossRef]
- Wang, Y.; Hua, S.; Cui, X.; Cao, Y.; Wen, J.; Chi, X.; Ji, C.; Pang, L.; You, L. The Effect of FOXC2-AS1 on White Adipocyte Browning and the Possible Regulatory Mechanism. Front. Endocrinol. 2020, 11, 565483. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Zhou, Y.; Cui, X.; Wang, X.; Sun, Y.; Gao, Y.; Wang, X.; Wen, J.; Xie, K.; Tang, R.; et al. GM13133 is a negative regulator in mouse white adipocytes differentiation and drives the characteristics of brown adipocytes. J. Cell. Physiol. 2018, 233, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Malakar, A.K.; Choudhury, D.; Halder, B.; Paul, P.; Uddin, A.; Chakraborty, S. A review on coronary artery disease, its risk factors, and therapeutics. J. Cell. Physiol. 2019, 234, 16812–16823. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.C.; Gerhardt, T.E.; Kwon, E. Risk factors for coronary artery disease. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Zareba, L.; Fitas, A.; Wolska, M.; Junger, E.; Eyileten, C.; Wicik, Z.; De Rosa, S.; Siller-Matula, J.M.; Postula, M. MicroRNAs and Long Noncoding RNAs in Coronary Artery Disease: New and Potential Therapeutic Targets. Cardiol. Clin. 2020, 38, 601–617. [Google Scholar] [CrossRef] [PubMed]
- Cardona-Monzonis, A.; Garcia-Gimenez, J.L.; Mena-Molla, S.; Pareja-Galeano, H.; de la Guia-Galipienso, F.; Lippi, G.; Pallardo, F.V.; Sanchis-Gomar, F. Non-coding RNAs and Coronary Artery Disease. Adv. Exp. Med. Biol. 2020, 1229, 273–285. [Google Scholar] [CrossRef]
- Dong, X.H.; Lu, Z.F.; Kang, C.M.; Li, X.H.; Haworth, K.E.; Ma, X.; Lu, J.B.; Liu, X.H.; Fang, F.C.; Wang, C.S.; et al. The Long Noncoding RNA RP11-728F11.4 Promotes Atherosclerosis. Arter. Thromb. Vasc. Biol. 2021, 41, 1191–1204. [Google Scholar] [CrossRef]
- Camargo, F.N.; Matos, S.L.; Araujo, L.C.C.; Carvalho, C.R.O.; Amaral, A.G.; Camporez, J.P. Western Diet-Fed ApoE Knockout Male Mice as an Experimental Model of Non-Alcoholic Steatohepatitis. Curr. Issues Mol. Biol. 2022, 44, 4692–4703. [Google Scholar] [CrossRef]
- Kampschulte, M.; Stockl, C.; Langheinrich, A.C.; Althohn, U.; Bohle, R.M.; Krombach, G.A.; Stieger, P.; Churin, Y.; Kremer, S.; Dierkes, C.; et al. Western diet in ApoE-LDLR double-deficient mouse model of atherosclerosis leads to hepatic steatosis, fibrosis, and tumorigenesis. Lab. Invest. 2014, 94, 1273–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.H.; Deng, W.Y.; Chen, J.J.; Xu, X.D.; Liu, X.X.; Chen, L.; Shi, M.W.; Liu, Q.X.; Tao, M.; Ren, K. LncRNA kcnq1ot1 promotes lipid accumulation and accelerates atherosclerosis via functioning as a ceRNA through the miR-452-3p/HDAC3/ABCA1 axis. Cell Death Dis. 2020, 11, 1043. [Google Scholar] [CrossRef]
- Wang, B.; Su, Z.; Wan, L.; He, T. Relationship between long non-coding RNA polymorphism and the risk of coronary artery disease: A protocol for systematic review and meta-analysis. Medicine 2021, 100, e25146. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Gholipour, M.; Taheri, M. The Emerging Role of Long Non-coding RNAs and Circular RNAs in Coronary Artery Disease. Front. Cardiovasc. Med. 2021, 8, 632393. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, Y.; Huang, J.; Liu, Y.; Zhu, Y. Association of genetic variants in lncRNA GAS5/miR-21/mTOR axis with risk and prognosis of coronary artery disease among a Chinese population. J. Clin. Lab. Anal. 2020, 34, e23430. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, M.; Li, J.; Liang, B.; Chen, Z.; Yang, J.; Guo, X.; Huang, S.; Gu, L.; Su, L. LncRNA H19 rs4929984 Variant is Associated with Coronary Artery Disease Susceptibility in Han Chinese Female Population. Biochem. Genet. 2021, 59, 1359–1380. [Google Scholar] [CrossRef]
- Hu, W.N.; Ding, H.X.; Xu, Q.; Zhang, X.Y.; Yang, D.T.; Jin, Y.Z. Relationship between Long Noncoding RNA H19 Polymorphisms and Risk of Coronary Artery Disease in a Chinese Population: A Case-Control Study. Dis. Markers 2020, 2020, 9839612. [Google Scholar] [CrossRef]
- Gao, W.; Zhu, M.; Wang, H.; Zhao, S.; Zhao, D.; Yang, Y.; Wang, Z.M.; Wang, F.; Yang, Z.J.; Lu, X.; et al. Association of polymorphisms in long non-coding RNA H19 with coronary artery disease risk in a Chinese population. Mutat. Res. 2015, 772, 15–22. [Google Scholar] [CrossRef]
- Kim, I.J.; Lee, J.Y.; Park, H.W.; Park, H.S.; Ko, E.J.; Sung, J.H.; Kim, N.K. Association between HOTAIR lncRNA Polymorphisms and Coronary Artery Disease Susceptibility. J. Pers. Med. 2021, 11, 375. [Google Scholar] [CrossRef]
- Song, N.; Luo, J.Y.; Zhao, Q.; Zhang, J.Y.; Liu, F.; Li, X.M.; Yang, Y.N. MALAT1 gene rs600231 polymorphism positively associated with acute coronary syndrome in Chinese population: A case-control study. Cardiovasc. Diagn. Ther. 2021, 11, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Ding, H.; Ouyang, A.; Zhang, X.; Xu, Q.; Han, Y.; Zhang, X.; Jin, Y. LncRNA MALAT1 gene polymorphisms in coronary artery disease: A case-control study in a Chinese population. Biosci. Rep. 2019, 39, BSR20182213. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Li, Y.; Peng, Y.; Tang, J.; Li, H. Association of polymorphisms in MALAT1 with risk of coronary atherosclerotic heart disease in a Chinese population. Lipids Health Dis. 2018, 17, 75. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.S.; Cheng, J.; Cai, M.Y.; Yang, X.L.; Liu, X.G.; Zheng, B.Y.; Xiong, X.D. Association of lincRNA-p21 Haplotype with Coronary Artery Disease in a Chinese Han Population. Dis. Markers 2016, 2016, 9109743. [Google Scholar] [CrossRef]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef] [PubMed]
- Sherry, S.T.; Ward, M.H.; Kholodov, M.; Baker, J.; Phan, L.; Smigielski, E.M.; Sirotkin, K. dbSNP: The NCBI database of genetic variation. Nucleic Acids Res. 2001, 29, 308–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landrum, M.J.; Lee, J.M.; Benson, M.; Brown, G.R.; Chao, C.; Chitipiralla, S.; Gu, B.; Hart, J.; Hoffman, D.; Jang, W.; et al. ClinVar: Improving access to variant interpretations and supporting evidence. Nucleic Acids Res. 2018, 46, D1062–D1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Prim. 2015, 1, 15019. [Google Scholar] [CrossRef]
- Olokoba, A.B.; Obateru, O.A.; Olokoba, L.B. Type 2 diabetes mellitus: A review of current trends. Oman Med. J. 2012, 27, 269–273. [Google Scholar] [CrossRef]
- Khan, M.A.B.; Hashim, M.J.; King, J.K.; Govender, R.D.; Mustafa, H.; Al Kaabi, J. Epidemiology of Type 2 Diabetes—Global Burden of Disease and Forecasted Trends. J. Epidemiol. Glob. Health 2020, 10, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Schnurr, T.M.; Jakupovic, H.; Carrasquilla, G.D.; Angquist, L.; Grarup, N.; Sorensen, T.I.A.; Tjonneland, A.; Overvad, K.; Pedersen, O.; Hansen, T.; et al. Obesity, unfavourable lifestyle and genetic risk of type 2 diabetes: A case-cohort study. Diabetologia 2020, 63, 1324–1332. [Google Scholar] [CrossRef]
- Kyrou, I.; Tsigos, C.; Mavrogianni, C.; Cardon, G.; Van Stappen, V.; Latomme, J.; Kivela, J.; Wikstrom, K.; Tsochev, K.; Nanasi, A.; et al. Sociodemographic and lifestyle-related risk factors for identifying vulnerable groups for type 2 diabetes: A narrative review with emphasis on data from Europe. BMC Endocr. Disord. 2020, 20, 134. [Google Scholar] [CrossRef] [Green Version]
- Leong, A.; Porneala, B.; Dupuis, J.; Florez, J.C.; Meigs, J.B. Type 2 Diabetes Genetic Predisposition, Obesity, and All-Cause Mortality Risk in the U.S.: A Multiethnic Analysis. Diabetes Care 2016, 39, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Martin-Timon, I.; Sevillano-Collantes, C.; Segura-Galindo, A.; Del Canizo-Gomez, F.J. Type 2 diabetes and cardiovascular disease: Have all risk factors the same strength? World J. Diabetes 2014, 5, 444–470. [Google Scholar] [CrossRef]
- Sun, D.; Zhou, T.; Heianza, Y.; Li, X.; Fan, M.; Fonseca, V.A.; Qi, L. Type 2 Diabetes and Hypertension. Circ. Res. 2019, 124, 930–937. [Google Scholar] [CrossRef]
- Duh, E.J.; Sun, J.K.; Stitt, A.W. Diabetic retinopathy: Current understanding, mechanisms, and treatment strategies. JCI Insight 2017, 2, e93751. [Google Scholar] [CrossRef] [PubMed]
- McGill, J.B.; Haller, H.; Roy-Chaudhury, P.; Cherrington, A.; Wada, T.; Wanner, C.; Ji, L.; Rossing, P. Making an impact on kidney disease in people with type 2 diabetes: The importance of screening for albuminuria. BMJ Open Diabetes Res. Care 2022, 10, e002806. [Google Scholar] [CrossRef] [PubMed]
- Chi, T.; Lin, J.; Wang, M.; Zhao, Y.; Liao, Z.; Wei, P. Non-Coding RNA as Biomarkers for Type 2 Diabetes Development and Clinical Management. Front. Endocrinol. 2021, 12, 630032. [Google Scholar] [CrossRef] [PubMed]
- Formichi, C.; Nigi, L.; Grieco, G.E.; Maccora, C.; Fignani, D.; Brusco, N.; Licata, G.; Sebastiani, G.; Dotta, F. Non-Coding RNAs: Novel Players in Insulin Resistance and Related Diseases. Int. J. Mol. Sci. 2021, 22, 7716. [Google Scholar] [CrossRef]
- Dieter, C.; Lemos, N.E.; Correa, N.R.F.; Assmann, T.S.; Crispim, D. The Impact of lncRNAs in Diabetes Mellitus: A Systematic Review and In Silico Analyses. Front. Endocrinol. 2021, 12, 602597. [Google Scholar] [CrossRef]
- Chen, Y.; He, Y.; Zhou, H. The potential role of lncRNAs in diabetes and diabetic microvascular complications. Endocr. J. 2020, 67, 659–668. [Google Scholar] [CrossRef]
- Hosen, M.R.; Militello, G.; Weirick, T.; Ponomareva, Y.; Dassanayaka, S.; Moore, J.B.; Doring, C.; Wysoczynski, M.; Jones, S.P.; Dimmeler, S.; et al. Airn Regulates Igf2bp2 Translation in Cardiomyocytes. Circ. Res. 2018, 122, 1347–1353. [Google Scholar] [CrossRef]
- Peng, T.; Liu, M.; Hu, L.; Guo, D.; Wang, D.; Qi, B.; Ren, G.; Hu, C.; Zhang, F.; Chun, H.J.; et al. LncRNA Airn alleviates diabetic cardiac fibrosis by inhibiting activation of cardiac fibroblasts via a m6A-IMP2-p53 axis. Biol. Direct 2022, 17, 32. [Google Scholar] [CrossRef]
- Miller, H.E.; Ilieva, M.; Bishop, A.J.R.; Uchida, S. Current Status of Epitranscriptomic Marks Affecting lncRNA Structures and Functions. Noncoding RNA 2022, 8, 23. [Google Scholar] [CrossRef]
- Leon, B.M.; Maddox, T.M. Diabetes and cardiovascular disease: Epidemiology, biological mechanisms, treatment recommendations and future research. World J. Diabetes 2015, 6, 1246–1258. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Xu, W.; Zhang, W.; Wang, W.; Liu, T.; Zhou, X. LncRNA DCRF regulates cardiomyocyte autophagy by targeting miR-551b-5p in diabetic cardiomyopathy. Theranostics 2019, 9, 4558–4566. [Google Scholar] [CrossRef]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2015, 70, 5.47.1–5.47.20. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Lin, Y.; Shen, H.; Yu, S.; Xu, J. LncRNA TUG1 exacerbates myocardial fibrosis in diabetic cardiomyopathy by modulating the microRNA-145a-5p/Cfl2 axis. J. Cardiovasc. Pharmacol. 2022; Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Tian, C.; Gao, Z.; Zhang, B.; Zhao, L. Research status and trends of the diabetic cardiomyopathy in the past 10 years (2012-2021): A bibliometric analysis. Front. Cardiovasc. Med. 2022, 9, 1018841. [Google Scholar] [CrossRef]
- Feng, B.; Liu, J.; Wang, E.; Su, Z.; Chakrabarti, S. Endothelial derived miRNA-9 mediated cardiac fibrosis in diabetes and its regulation by ZFAS1. PLoS ONE 2022, 17, e0276076. [Google Scholar] [CrossRef]
- Xiao, W.; Zheng, D.; Chen, X.; Yu, B.; Deng, K.; Ma, J.; Wen, X.; Hu, Y.; Hou, J. Long non-coding RNA MIAT is involved in the regulation of pyroptosis in diabetic cardiomyopathy via targeting miR-214-3p. iScience 2021, 24, 103518. [Google Scholar] [CrossRef]
- Ni, T.; Huang, X.; Pan, S.; Lu, Z. Inhibition of the long non-coding RNA ZFAS1 attenuates ferroptosis by sponging miR-150-5p and activates CCND2 against diabetic cardiomyopathy. J. Cell. Mol. Med. 2021, 25, 9995–10007. [Google Scholar] [CrossRef]
- Zhou, J.; Ji, X.; Zhang, F.; Yan, K.; Huang, P.; Huang, C. Bioinformatics analysis for identifying micro-RNAs, long noncoding RNAs, transcription factors, and immune genes regulatory networks in diabetic cardiomyopathy using an integrated bioinformatics analysis. Inflamm. Res. 2022, 71, 847–858. [Google Scholar] [CrossRef]
- Rai, A.K.; Lee, B.; Gomez, R.; Rajendran, D.; Khan, M.; Garikipati, V.N.S. Current Status and Potential Therapeutic Strategies for Using Non-coding RNA to Treat Diabetic Cardiomyopathy. Front. Physiol. 2020, 11, 612722. [Google Scholar] [CrossRef]
- Chen, K.; Ma, Y.; Wu, S.; Zhuang, Y.; Liu, X.; Lv, L.; Zhang, G. Construction and analysis of a lncRNA-miRNA-mRNA network based on competitive endogenous RNA reveals functional lncRNAs in diabetic cardiomyopathy. Mol. Med. Rep. 2019, 20, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Whelton, P.K.; Carey, R.M.; Mancia, G.; Kreutz, R.; Bundy, J.D.; Williams, B. Harmonization of the American College of Cardiology/American Heart Association and European Society of Cardiology/European Society of Hypertension Blood Pressure/Hypertension Guidelines: Comparisons, Reflections, and Recommendations. J. Am. Coll. Cardiol. 2022, 80, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Samara, V.A.; Das, S.; Reddy, M.A.; Tanwar, V.S.; Stapleton, K.; Leung, A.; Abdollahi, M.; Ganguly, R.; Lanting, L.; Natarajan, R. Angiotensin II-Induced Long Non-Coding RNA Alivec Regulates Chondrogenesis in Vascular Smooth Muscle Cells. Cells 2021, 10, 2696. [Google Scholar] [CrossRef]
- Hassoun, P.M. Pulmonary Arterial Hypertension. N. Engl. J. Med. 2021, 385, 2361–2376. [Google Scholar] [CrossRef]
- Mathew, R. Pulmonary hypertension and metabolic syndrome: Possible connection, PPARgamma and Caveolin-1. World J. Cardiol. 2014, 6, 692–705. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.J.; Vorla, M.; Kalra, D.K. Molecular Pathways in Pulmonary Arterial Hypertension. Int. J. Mol. Sci. 2022, 23, 10001. [Google Scholar] [CrossRef]
- Maron, B.A.; Loscalzo, J. Pulmonary hypertension: Pathophysiology and signaling pathways. Handb. Exp. Pharmacol. 2013, 218, 31–58. [Google Scholar] [CrossRef]
- Bernardi, N.; Bianconi, E.; Vecchi, A.; Ameri, P. Noncoding RNAs in Pulmonary Arterial Hypertension: Current Knowledge and Translational Perspectives. Heart Fail. Clin. 2023, 19, 137–152. [Google Scholar] [CrossRef]
- Deng, L.; Han, X.; Wang, Z.; Nie, X.; Bian, J. The Landscape of Noncoding RNA in Pulmonary Hypertension. Biomolecules 2022, 12, 796. [Google Scholar] [CrossRef]
- Shirazi-Tehrani, E.; Chamasemani, A.; Firouzabadi, N.; Mousaei, M. ncRNAs and polyphenols: New therapeutic strategies for hypertension. RNA Biol. 2022, 19, 575–587. [Google Scholar] [CrossRef]
- Zang, H.; Zhang, Q.; Li, X. Non-Coding RNA Networks in Pulmonary Hypertension. Front. Genet. 2021, 12, 703860. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Long, Y.; Kwoh, C.K. Deep learning based DNA:RNA triplex forming potential prediction. BMC Bioinform. 2020, 21, 522. [Google Scholar] [CrossRef] [PubMed]
- Antonov, I.; Medvedeva, Y.A. Purine-rich low complexity regions are potential RNA binding hubs in the human genome. F1000Research 2018, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Leisegang, M.S.; Bains, J.K.; Seredinski, S.; Oo, J.A.; Krause, N.M.; Kuo, C.C.; Gunther, S.; Senturk Cetin, N.; Warwick, T.; Cao, C.; et al. HIF1alpha-AS1 is a DNA:DNA:RNA triplex-forming lncRNA interacting with the HUSH complex. Nat. Commun. 2022, 13, 6563. [Google Scholar] [CrossRef]
- Grote, P.; Wittler, L.; Hendrix, D.; Koch, F.; Wahrisch, S.; Beisaw, A.; Macura, K.; Blass, G.; Kellis, M.; Werber, M.; et al. The tissue-specific lncRNA Fendrr is an essential regulator of heart and body wall development in the mouse. Dev. Cell 2013, 24, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, Q.; He, S.; Bai, J.; Ma, C.; Zhang, L.; Guan, X.; Yuan, H.; Li, Y.; Zhu, X.; et al. LncRNA FENDRR with m6A RNA methylation regulates hypoxia-induced pulmonary artery endothelial cell pyroptosis by mediating DRP1 DNA methylation. Mol. Med. 2022, 28, 126. [Google Scholar] [CrossRef]
- Ilieva, M.; Uchida, S. Perspectives of LncRNAs for therapy. Cell Biol. Toxicol. 2022, 38, 915–917. [Google Scholar] [CrossRef]
- Badowski, C.; He, B.; Garmire, L.X. Blood-derived lncRNAs as biomarkers for cancer diagnosis: The Good, the Bad and the Beauty. NPJ Precis. Oncol. 2022, 6, 40. [Google Scholar] [CrossRef]
- Lu, C.; Wei, D.; Zhang, Y.; Wang, P.; Zhang, W. Long Non-Coding RNAs as Potential Diagnostic and Prognostic Biomarkers in Breast Cancer: Progress and Prospects. Front. Oncol. 2021, 11, 710538. [Google Scholar] [CrossRef]
- Pant, T.; Dhanasekaran, A.; Zhao, M.; Thorp, E.B.; Forbess, J.M.; Bosnjak, Z.J.; Benjamin, I.J.; Ge, Z.D. Identification and analysis of circulating long non-coding RNAs with high significance in diabetic cardiomyopathy. Sci. Rep. 2021, 11, 2571. [Google Scholar] [CrossRef]
- Ismail, N.; Abdullah, N.; Abdul Murad, N.A.; Jamal, R.; Sulaiman, S.A. Long Non-Coding RNAs (lncRNAs) in Cardiovascular Disease Complication of Type 2 Diabetes. Diagnostics 2021, 11, 145. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
LncRNAs associated with obesity. (A) The H19–IGF2 locus consists of a maternal unmethylated (white lollipops in the image above) imprinting control region (ICR; yellow box). ICR is bound by the insulator protein CCCTC-binding factor (CTCF), which prevents access of the IGF2 promoter (blue box represents the exon from the paternal chromosome) to downstream enhancers. The overexpression of the lncRNA H19 (pink box represents the exon from the maternal chromosome) in mice has a protective effect against diet-induced obesity and increased insulin sensitivity. In the image above, white boxes represent the repressed genes, while black lollipops represent the methylated CpG sites. (B) The SRA1 gene encodes for protein-coding mRNA and lncRNA Sra1. The lncRNA Sra1 functions as a transcription co-activator of peroxisome proliferator-activated receptor gamma (Pparg), which is a regulator of adipocytes differentiation and lipid accumulation. (C) The lncRNA ADIPINT is upregulated in obesity. This lncRNA is responsible for adipocyte size, insulin resistance, and pyruvate carboxylase activity. (D) Several lncRNAs have been identified to be involved in the process of browning and whitening in the adipose tissue, which could serve as potential therapeutic targets. Figure created with BioRender.com (accessed on 10 January 2022).
Figure 1.
LncRNAs associated with obesity. (A) The H19–IGF2 locus consists of a maternal unmethylated (white lollipops in the image above) imprinting control region (ICR; yellow box). ICR is bound by the insulator protein CCCTC-binding factor (CTCF), which prevents access of the IGF2 promoter (blue box represents the exon from the paternal chromosome) to downstream enhancers. The overexpression of the lncRNA H19 (pink box represents the exon from the maternal chromosome) in mice has a protective effect against diet-induced obesity and increased insulin sensitivity. In the image above, white boxes represent the repressed genes, while black lollipops represent the methylated CpG sites. (B) The SRA1 gene encodes for protein-coding mRNA and lncRNA Sra1. The lncRNA Sra1 functions as a transcription co-activator of peroxisome proliferator-activated receptor gamma (Pparg), which is a regulator of adipocytes differentiation and lipid accumulation. (C) The lncRNA ADIPINT is upregulated in obesity. This lncRNA is responsible for adipocyte size, insulin resistance, and pyruvate carboxylase activity. (D) Several lncRNAs have been identified to be involved in the process of browning and whitening in the adipose tissue, which could serve as potential therapeutic targets. Figure created with BioRender.com (accessed on 10 January 2022).

Figure 2.
LncRNAs associated with atherosclerosis development. (A) The lncRNA FXYD6-AS1 is overexpressed in the patients with atherosclerosis as well as in the apoE deficient mice fed with a Western diet. Together with EWS RNA binding protein 1 (EWSR1), FXYD6-AS1 regulates the expression of the protein-coding gene FXYD6, which is the main modulator of Na+/K+-ATPase activity. (B) The lncRNA KCNQ1OT1 functions as a miRNA sponge. This lncRNA sequesters miR-452-3p, which binds to histone deacetylase 3 (HDAC3) and thus inhibits the expression of ATP binding cassette subfamily A member 1 (ABCA1). Phenotypically, this cascade results in lipid accumulation and proinflammatory cytokine release in macrophages and accelerates the atherosclerotic development. Figure created with BioRender.com (accessed on 9 December 2022).
Figure 2.
LncRNAs associated with atherosclerosis development. (A) The lncRNA FXYD6-AS1 is overexpressed in the patients with atherosclerosis as well as in the apoE deficient mice fed with a Western diet. Together with EWS RNA binding protein 1 (EWSR1), FXYD6-AS1 regulates the expression of the protein-coding gene FXYD6, which is the main modulator of Na+/K+-ATPase activity. (B) The lncRNA KCNQ1OT1 functions as a miRNA sponge. This lncRNA sequesters miR-452-3p, which binds to histone deacetylase 3 (HDAC3) and thus inhibits the expression of ATP binding cassette subfamily A member 1 (ABCA1). Phenotypically, this cascade results in lipid accumulation and proinflammatory cytokine release in macrophages and accelerates the atherosclerotic development. Figure created with BioRender.com (accessed on 9 December 2022).

Figure 3.
LncRNAs associated with diabetes. (A) The lncRNA Airn is paternally imprinted and located in antisense orientation to the maternally imprinted derived protein-coding gene Igf2r. Airn binds to the RNA-binding protein Igf2bp2 and controls the translation of mRNAs (e.g., biglycan and inhibin β-A) to modulate the resistance to stress in cardiomyocytes. Similarly, in diabetic cardiac fibrosis, Airn binds to Igf2bp2 to regulate the degradation of Trp53 mRNA in a m6A manner. (B) Several lncRNAs are involved in diabetic cardiomyopathy (DCM) by functioning as miRNA sponges. The lncRNA DCRF sequester miR551b-5p targets protocadherin 17 mRNA to regulate autophagy in rat cardiomyocytes. The upregulation of the lncRNA Tug1 in the DCM streptozotocin-based mouse model leads to an increasing level of extracellular matrix and myocardial fibrosis development. Figure created with BioRender.com (accessed on 9 December 2022).
Figure 3.
LncRNAs associated with diabetes. (A) The lncRNA Airn is paternally imprinted and located in antisense orientation to the maternally imprinted derived protein-coding gene Igf2r. Airn binds to the RNA-binding protein Igf2bp2 and controls the translation of mRNAs (e.g., biglycan and inhibin β-A) to modulate the resistance to stress in cardiomyocytes. Similarly, in diabetic cardiac fibrosis, Airn binds to Igf2bp2 to regulate the degradation of Trp53 mRNA in a m6A manner. (B) Several lncRNAs are involved in diabetic cardiomyopathy (DCM) by functioning as miRNA sponges. The lncRNA DCRF sequester miR551b-5p targets protocadherin 17 mRNA to regulate autophagy in rat cardiomyocytes. The upregulation of the lncRNA Tug1 in the DCM streptozotocin-based mouse model leads to an increasing level of extracellular matrix and myocardial fibrosis development. Figure created with BioRender.com (accessed on 9 December 2022).

Figure 4.
LncRNAs associated with hypertension. (A) In vascular smooth muscle cells, the expression of the lncRNA Alivec is induced by the growth factors AngII, TNFa, and PDGF via the transcription factor Sox9. Mechanistically, Alivec binds to tropomyosin 3 and Hnrnpa2b1 and regulates chondrogenic transformation of smooth muscles observed in vascular dysfunction and hypertension. (B) A mechanism by which lncRNAs implement a regulatory function through the DNA:DNA:RNA triplex formation. In human endothelial cells, the lncRNA HIF1A-AS1 serves as an adapter for the repressive human silencing hub complex (HUSH). Another lncRNA, FENDRR, forms a triplex structure in the promotor region of dynamin 1 like (DNM1L). Figure created with BioRender.com (accessed on 9 December 2022).
Figure 4.
LncRNAs associated with hypertension. (A) In vascular smooth muscle cells, the expression of the lncRNA Alivec is induced by the growth factors AngII, TNFa, and PDGF via the transcription factor Sox9. Mechanistically, Alivec binds to tropomyosin 3 and Hnrnpa2b1 and regulates chondrogenic transformation of smooth muscles observed in vascular dysfunction and hypertension. (B) A mechanism by which lncRNAs implement a regulatory function through the DNA:DNA:RNA triplex formation. In human endothelial cells, the lncRNA HIF1A-AS1 serves as an adapter for the repressive human silencing hub complex (HUSH). Another lncRNA, FENDRR, forms a triplex structure in the promotor region of dynamin 1 like (DNM1L). Figure created with BioRender.com (accessed on 9 December 2022).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ilieva, M.; Uchida, S. Potential Involvement of LncRNAs in Cardiometabolic Diseases. Genes 2023, 14, 213. https://doi.org/10.3390/genes14010213
AMA Style
Ilieva M, Uchida S. Potential Involvement of LncRNAs in Cardiometabolic Diseases. Genes. 2023; 14(1):213. https://doi.org/10.3390/genes14010213
Chicago/Turabian StyleIlieva, Mirolyuba, and Shizuka Uchida. 2023. "Potential Involvement of LncRNAs in Cardiometabolic Diseases" Genes 14, no. 1: 213. https://doi.org/10.3390/genes14010213
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.