
Germplasm Enhancement and Identification of Loci Conferring Resistance against Plasmodiophora brassicae in Broccoli

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
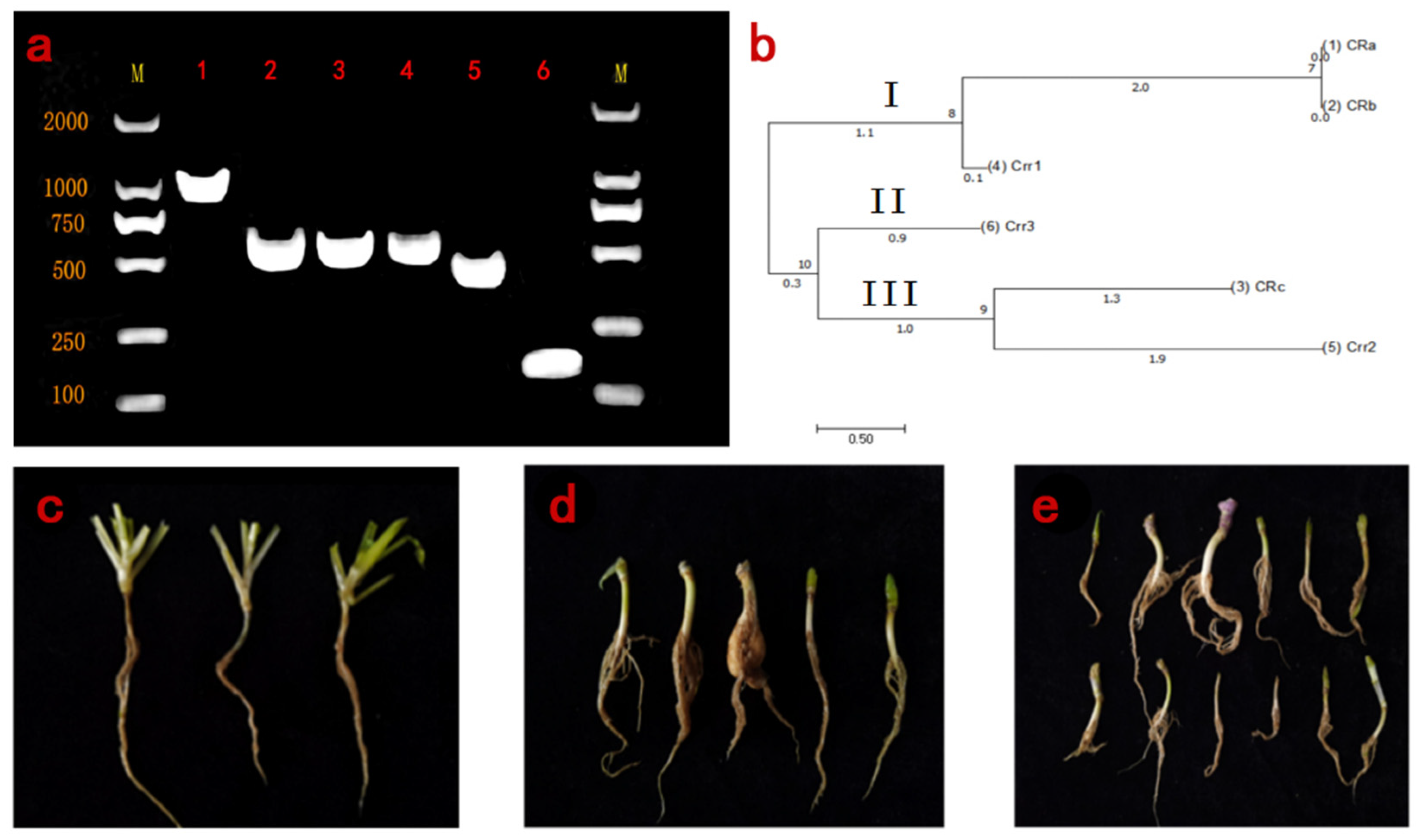
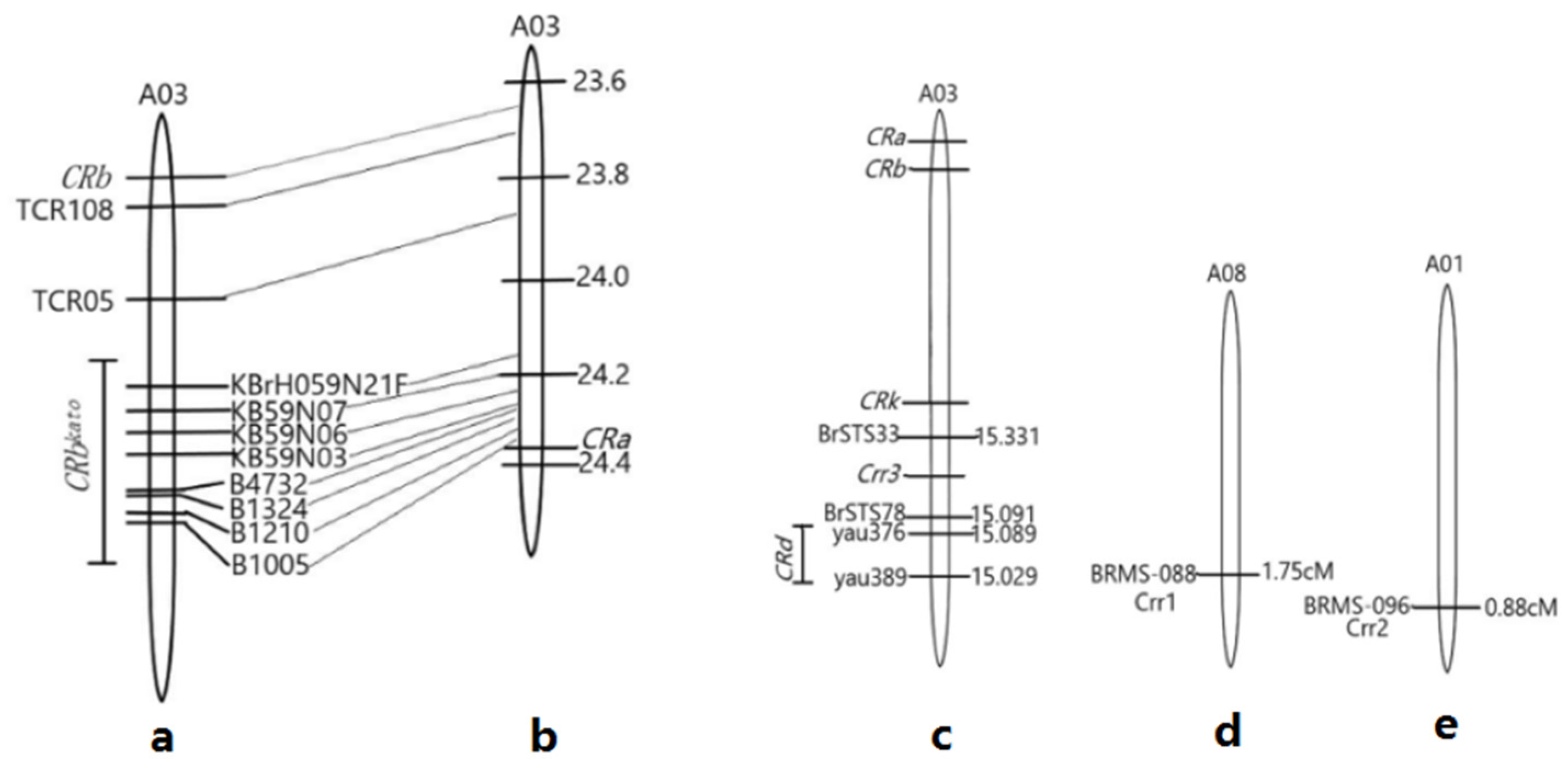
2.2. Molecular Markers and Identification of Clubroot Resistance Genes
2.3. Pathogen Inoculation and Molecular Verification
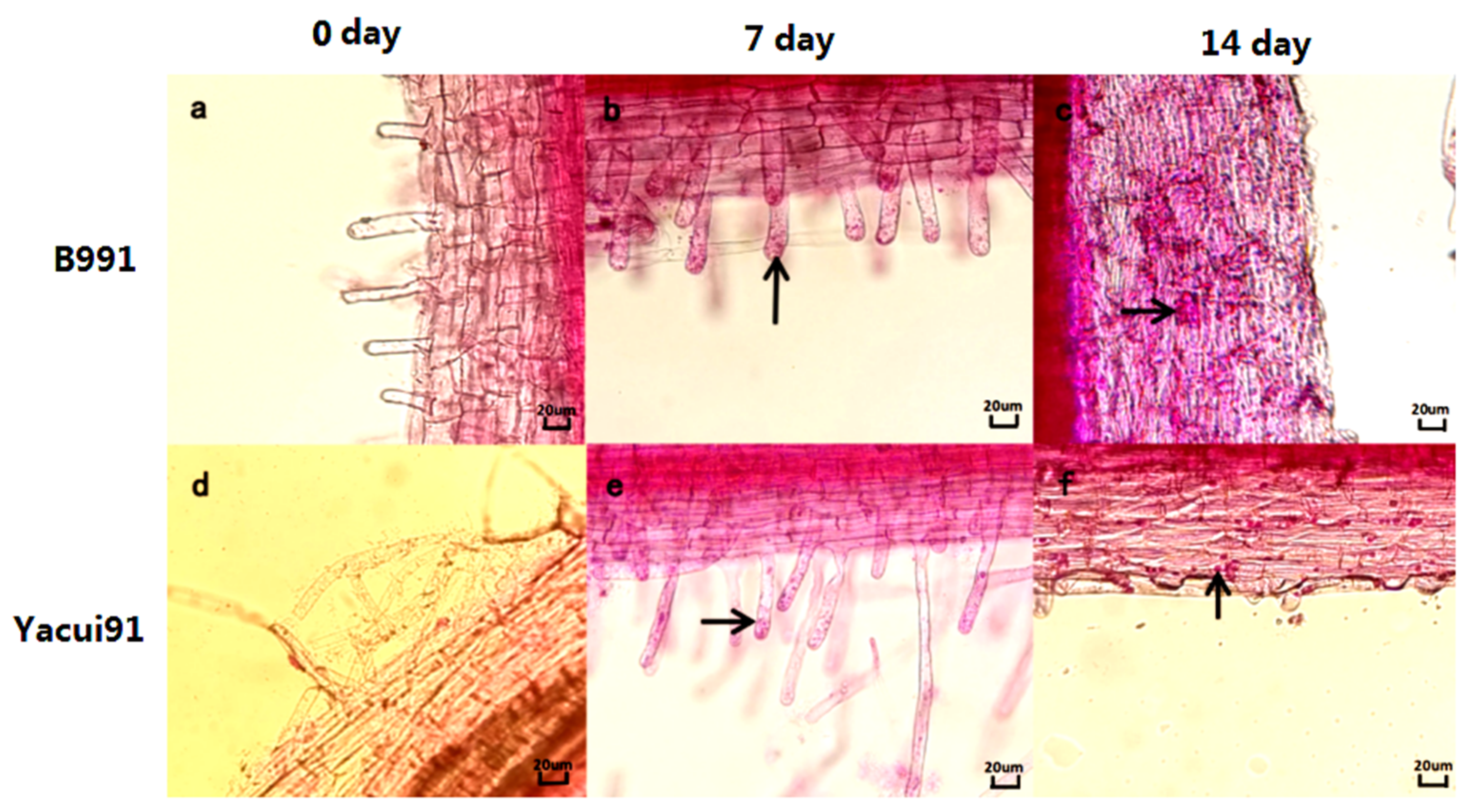
2.4. Staining and Observation
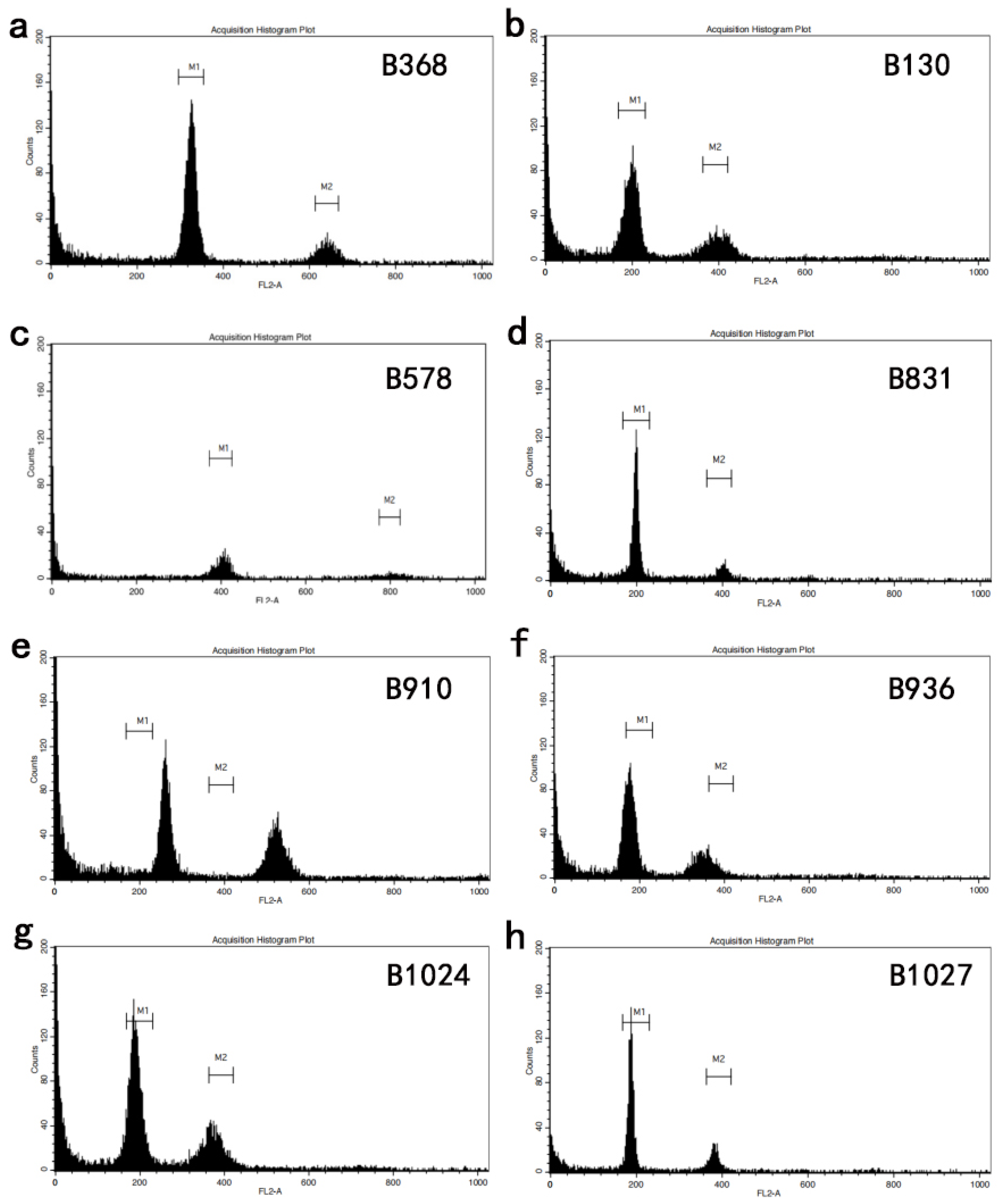
2.5. Ploidy Detection by Flow Cytometry
3. Results
3.1. Diversity of Clubroot Resistance and Plant Ploidy
3.2. Disease Identification and Phylogenetic Analysis of CR Genes
3.3. Comparisons of the Infection Processes of P. brassicae in Resistant and Susceptible Broccoli Cultivars
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hatakeyama, K.; Suwabe, K.; Tomita, R.N.; Kato, T.; Nunome, T.; Fukuoka, H.; Matsumoto, S. Identification and characterization of Crr1a, a gene for resistance to clubroot disease (Plasmodiophora brassicae Woronin) in Brassica rapa L. PLoS ONE 2013, 8, e54745. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Qiu, L.; Zhang, Z.; Liu, K.; Xia, X.; Xiong, S.; Zhao, S.; Zhao, Z.; Hu, Y.; Liang, Y. Control of Streptomyces alfalfae XY25 (T) Over Clubroot Disease and Its Effect on Rhizosphere Microbial Community in Chinese Cabbage Field Trials. Front. Microbiol. 2021, 12, 641556. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.; Fu, P.; Li, X.; Zhan, Z.; Yu, S.; Piao, Z. Identification and Mapping of the Clubroot Resistance Gene CRd in Chinese Cabbage (Brassica rapa ssp. pekinensis). Front. Plant Sci. 2018, 9, 653. [Google Scholar] [CrossRef] [PubMed]
- Mehraj, H.; Akter, A.; Miyaji, N.; Miyazaki, J.; Shea, D.J.; Fujimoto, R.; Doullah, M.A. Genetics of Clubroot and Fusarium Wilt Disease Resistance in Brassica Vegetables: The Application of Marker Assisted Breeding for Disease Resistance. Plants 2020, 9, 726. [Google Scholar] [CrossRef]
- Piao, Z.; Ramchiary, N.; Lim, Y.P. Genetics of Clubroot Resistance in Brassica Species. J. Plant Growth Regul. 2009, 28, 252–264. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Y.; Li, L.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Lv, H. Transcriptome reveals the gene expression patterns of sulforaphane metabolism in broccoli florets. PLoS ONE 2019, 14, e0213902. [Google Scholar] [CrossRef]
- Li, Z.; Mei, Y.; Liu, Y.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Lv, H. The evolution of genetic diversity of broccoli cultivars in China since 1980. Sci. Hortic. 2019, 250, 69–80. [Google Scholar] [CrossRef]
- Li, Z.; Zheng, S.; Liu, Y.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Lv, H.; Wang, Y.; Xu, D. Characterization of glucosinolates in 80 broccoli genotypes and different organs using UHPLC-Triple-TOF-MS method. Food Chem. 2021, 334, 127519. [Google Scholar] [CrossRef]
- Huang, J.; Liu, Y.; Han, F.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Lv, H.; Wang, Y.; Ji, J.; et al. Genetic diversity and population structure analysis of 161 broccoli cultivars based on SNP markers. Hortic. Plant J. 2021, 7, 423–433. [Google Scholar] [CrossRef]
- Hasan, J.; Megha, S.; Rahman, H. Clubroot in Brassica: Recent advances in genomics, breeding, and disease management. Genome 2021, 64, 735–760. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Y.; Yuan, S.; Han, F.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Lv, H.; Wang, Y.; et al. Fine mapping of the major QTLs for biochemical variation of sulforaphane in broccoli florets using a DH population. Sci. Rep. 2021, 11, 9004. [Google Scholar] [CrossRef] [PubMed]
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Matsumoto, S.; Hirai, M. Identification of two loci for resistance to clubroot (Plasmodiophora brassicae Woronin) in Brassica rapa L. Theor. Appl. Genet. 2003, 107, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Hatakeyama, K.; Fukino, N.; Matsumoto, S. Identificaiton of a clubroot resistance locus conferring resistance to a Plasmodiophora brassicae classified into pathotype group 3 in Chinese cabbage (Brassica rapa L.). Breed. Sci. 2012, 62, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Ueno, H.; Matsumoto, E.; Aruga, D.; Kitagawa, S.; Matsumura, H.; Hayashida, N. Molecular characterization of the CRa gene conferring clubroot resistance in Brassica rapa. Plant Mol. Biol. 2012, 80, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Feng, J.; Zhang, S.; Zhang, S.; Li, F.; Strelkov, S.E.; Sun, R.; Hwang, S.F. Resistance to Plasmodiophora brassicae in Brassica rapa and Brassica juncea genotypes From China. Plant Dis. 2015, 99, 776–779. [Google Scholar] [CrossRef]
- Pang, W.; Liang, Y.; Zhan, Z.; Li, X.; Piao, Z. Development of a Sinitic Clubroot Differential Set for the Pathotype Classification of Plasmodiophora brassicae. Front. Plant Sci. 2020, 11, 568771. [Google Scholar] [CrossRef] [PubMed]
- Ce, F.; Mei, J.; He, H.; Zhao, Y.; Hu, W.; Yu, F.; Li, Q.; Ren, X.; Si, J.; Song, H.; et al. Identification of Candidate Genes for Clubroot-Resistance in Brassica oleracea Using Quantitative Trait Loci-Sequencing. Front. Plant Sci. 2021, 12, 703520. [Google Scholar] [CrossRef]
- Matsumoto, E.; Ueno, H.; Aruga, D.; Sakamoto, K.; Hayashida, N. Accumulation of Three Clubroot Resistance Genes through Marker-assisted Selection in Chinese Cabbage (Brassica rapa ssp. pekinensis). J. Jpn. Soc. Hortic. Sci. 2012, 81, 184–190. [Google Scholar] [CrossRef]
- Hirai, M.; Harada, T.; Kubo, N.; Tsukada, M.; Suwabe, K.; Matsumoto, S. A novel locus for clubroot resistance in Brassica rapa and its linkage markers. Theor. Appl. Genet. 2004, 108, 639–643. [Google Scholar] [CrossRef]
- Jia, H.; Wei, X.; Yang, Y.; Yuan, Y.; Wei, F.; Zhao, Y.; Yang, S.; Yao, Q.; Wang, Z.; Tian, B.; et al. Root RNA-seq analysis reveals a distinct transcriptome landscape between clubroot-susceptible and clubroot-resistant Chinese cabbage lines after Plasmodiophora brassicae infection. Plant Soil 2017, 421, 93–105. [Google Scholar] [CrossRef]
- Li, Y.; Yuan, Y.; Yanyan, Z.; Xiaochun, W.; Qiu-ju, Y.; Wusheng, J.; Zhiyong, W.; Shuangjuan, Y.; Xiaowei, Z.; Tian, B. Pathogen Identification of Clubroot Disease in Chinese Cabbage from Yuanyang County, Henan Province. Mol. Plant Breed. 2017, 8. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, Y.; Zhao, Y.; Xie, Z.; Hossain, M.R.; Yang, S.; Shi, G.; Lv, Y.; Wang, Z.; Tian, B.; et al. Root Transcriptome and Metabolome Profiling Reveal Key Phytohormone-Related Genes and Pathways Involved Clubroot Resistance in Brassica rapa L. Front. Plant Sci. 2021, 12, 759623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, Y.; Fang, Z.; Li, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Lv, H. Comparative Transcriptome Analysis between Broccoli (Brassica oleracea var. italica) and Wild Cabbage (Brassica macrocarpa Guss.) in Response to Plasmodiophora brassicae during Different Infection Stages. Front. Plant Sci. 2016, 7, 1929. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.F.; Strelkov, S.E.; Feng, J.; Gossen, B.D.; Howard, R.J. Plasmodiophora brassicae: A review of an emerging pathogen of the Canadian canola (Brassica napus) crop. Mol. Plant Pathol. 2012, 13, 105–113. [Google Scholar] [CrossRef]
- Askarian, H.; Akhavan, A.; González, L.G.; Hwang, S.F.; Strelkov, S.E. Genetic Structure of Plasmodiophora brassicae Populations Virulent on Clubroot Resistant Canola (Brassica napus). Plant Dis. 2021, 105, 3694–3704. [Google Scholar] [CrossRef]
- Zhang, H.; Feng, J.; Manolii, V.P.; Strelkov, S.E.; Hwang, S.F. Characterization of a Gene Identified in Pathotype 5 of the Clubroot Pathogen Plasmodiophora brassicae. Phytopathology 2015, 105, 764–770. [Google Scholar] [CrossRef]
- Chen, J.; Jing, J.; Zhan, Z.; Zhang, T.; Zhang, C.; Piao, Z. Identification of novel QTLs for isolate-specific partial resistance to Plasmodiophora brassicae in Brassica rapa. PLoS ONE 2013, 8, e85307. [Google Scholar] [CrossRef]
- Diederichsen, E.; Beckmann, J.; Schondelmeier, J.; Dreyer, F. Genetics of Clubroot Resistance in Brassica napus ‘Mendel’. Acta Hortic. 2006, 706, 307–312. [Google Scholar] [CrossRef]
- Piao, Z.Y.; Deng, Y.Q.; Choi, S.R.; Park, Y.J.; Lim, Y.P. SCAR and CAPS mapping of CRb, a gene conferring resistance to Plasmodiophora brassicae in Chinese cabbage ( Brassica rapa ssp. pekinensis). Theor. Appl. Genet. 2004, 108, 1458–1465. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Z.; Zhang, C.; Pang, W.; Choi, S.R.; Lim, Y.P.; Piao, Z. Fine genetic and physical mapping of the CRb gene conferring resistance to clubroot disease in Brassica rapa. Mol. Breed. 2014, 34, 1173–1183. [Google Scholar] [CrossRef]
- Vesnina, A.; Prosekov, A.; Kozlova, O.; Atuchin, V. Genes and Eating Preferences, Their Roles in Personalized Nutrition. Genes 2020, 11, 357. [Google Scholar] [CrossRef]
- Saito, M.; Kubo, N.; Matsumoto, S.; Suwabe, K.; Tsukada, M.; Hirai, M. Fine mapping of the clubroot resistance gene, Crr3, in Brassica rapa. Theor. Appl. Genet. 2006, 114, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Saito, A.; Hayashida, N.; Taguchi, G.; Matsumoto, E. Mapping of isolate-specific QTLs for clubroot resistance in Chinese cabbage (Brassica rapa L. ssp. pekinensis). Theor. Appl. Genet. 2008, 117, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Z.; Nwafor, C.C.; Hou, Z.; Gong, J.; Zhu, B.; Jiang, Y.; Zhou, Y.; Wu, J.; Piao, Z.; Tong, Y.; et al. Cytological and morphological analysis of hybrids between Brassicoraphanus, and Brassica napus for introgression of clubroot resistant trait into Brassica napus L. PLoS ONE 2017, 12, e0177470. [Google Scholar] [CrossRef]
- Wang, Y.; Xiang, X.; Huang, F.; Yu, W.; Zhou, X.; Li, B.; Zhang, Y.; Chen, P.; Zhang, C. Fine Mapping of Clubroot Resistance Loci CRA8.1 and Candidate Gene Analysis in Chinese Cabbage (Brassica rapa L.). Front. Plant Sci. 2022, 13, 898108. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accessions | Species | Generations | Source Origin | Disease Resistant | Year |
|---|---|---|---|---|---|
| B891 | Tuscan kale (Brassica oleracea L. var. acephala) | F12 | CAAS-IVF, Beijing, China | S | 2020 |
| B991 | Broccoli (Brassica oleracea L. var. italica) | F6 | CAAS-IVF, Beijing, China | S | 2020 |
| B994 | Broccoli (Brassica oleracea L. var. italica) | F1 | Syngenta, China | S | 2020 |
| B1007 | Chinese black moss (Brassica campestris L.var. purpurea Baileysh) | OP | Hubei, China | S | 2020 |
| B1081 | Black mustard (Brassica nigra) | F12 | CAAS-IVF, Beijing, China | S | 2020 |
| B1082 | Abyssinian mustard (Brassica carinata) | F12 | CAAS-IVF, Beijing, China | S | 2020 |
| B1083 | Black mustard (Brassica nigra) | OP | CAAS-IVF, Beijing, China | S | 2020 |
| B1084 | Black mustard (Brassica nigra) | OP | CAAS-IVF, Beijing, China | S | 2020 |
| B1086 | Black mustard (Brassica nigra) | OP | CAAS-IVF, Beijing, China | S | 2020 |
| B359 | Broccoli (Brassica oleracea L. var. italica) | BC1 | CAAS-IVF, Beijing, China | S | 2021 |
| B366 | Broccoli (Brassica oleracea L. var. italica) Wild cabbage (Brassica macrocarpa Guss.) | BC1 | CAAS-IVF, Beijing, China | S, R | 2021 |
| B368 | Broccoli (Brassica oleracea L. var. italica) Wild cabbage (Brassica macrocarpa Guss.) Rape (Brassica napus L.) | BC1 | CAAS-IVF, Beijing, China | S, R, MR | 2021 |
| B369 | Broccoli (Brassica oleracea L. var. italica) Wild cabbage (Brassica macrocarpa Guss.) Rape (Brassica napus L.) | BC1 | CAAS-IVF, Beijing, China | S, R, MR | 2021 |
| B571 | Broccoli (Brassica oleracea L. var. italica) Turnip (Brassica rapa L. ssp. rapa) | F1 | CAAS-IVF, Beijing, China | HR | 2021 |
| B578 | Rape (Brassica napus L.) (Oil rape) | F1 | CAAS-IVF, Beijing, China | HR | 2021 |
| B581 | Turnip (Brassica rapa L. ssp. rapa) | F11 | CAAS-IVF, Beijing, China | HR | 2021 |
| B582 | Yellow rocket (Barbarea vulgaris R. Br.) | F11 | CAAS-IVF, Beijing, China | I | 2021 |
| B606 | Red cabbage (Brassica oleracea L. var. capitate) Turnip (Brassica rapa L. ssp. rapa) | BC1F3 | CAAS-IVF, Beijing, China | S, R, MR | 2021 |
| B607 | Red cabbage (Brassica oleracea L. var. capitate) Turnip (Brassica rapa L. ssp. rapa) | BC1F3 | CAAS-IVF, Beijing, China | S, R, MR | 2021 |
| B608 | Black mustard (Brassica nigra) | BC1F3 | CAAS-IVF, Beijing, China | S | 2021 |
| B611 | Red cabbage (Brassica oleracea L. var. capitate) Turnip (Brassica rapa L. ssp. rapa) | BC1F3 | CAAS-IVF, Beijing, China | S, R | 2021 |
| B612 | Red cabbage (Brassica oleracea L. var. capitate) Turnip (Brassica rapa L. ssp. rapa) | BC1F3 | CAAS-IVF, Beijing, China | S, R | 2021 |
| B613 | Red cabbage (Brassica oleracea L. var. capitate) Turnip (Brassica rapa L. ssp. rapa) | BC1F3 | CAAS-IVF, Beijing, China | S, R | 2021 |
| B614 | Red cabbage (Brassica oleracea L. var. capitate) Turnip (Brassica rapa L. ssp. rapa) | BC1F3 | CAAS-IVF, Beijing, China | S, R | 2021 |
| B621 | Red cabbage (Brassica oleracea L. var. capitate) Kohlrabi (Brassica oleracea L. var. caulorapa) | BC2 | CAAS-IVF, Beijing, China | S, R | 2021 |
| B831 | Broccoli (Brassica oleracea L. var. italica) Wild cabbage (Brassica macrocarpa Guss.) Rape (Brassica napus L.) | BC1 | CAAS-IVF, Beijing, China | S, R | 2021 |
| B832 | Broccoli (Brassica oleracea L. var. italica) Wild cabbage (Brassica macrocarpa Guss.) Rape (Brassica napus L.) | BC1 | CAAS-IVF, Beijing, China | S, R | 2021 |
| B908 | Broccoli (Brassica oleracea L. var. italica) Turnip (Brassica rapa L. ssp. rapa) Rape (Brassica napus L.) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B909 | Broccoli (Brassica oleracea L. var. italica) Turnip (Brassica rapa L. ssp. rapa) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B910 | Broccoli (Brassica oleracea L. var. italica) Turnip (Brassica rapa L. ssp. rapa) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B932 | Broccoli (Brassica oleracea L. var. italica) Turnip (Brassica rapa L. ssp. rapa) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B933 | Broccoli (Brassica oleracea L. var. italica) Turnip (Brassica rapa L. ssp. rapa) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B934 | Broccoli (Brassica oleracea L. var. italica) Turnip (Brassica rapa L. ssp. rapa) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B935 | Broccoli (Brassica oleracea L. var. italica) Turnip (Brassica rapa L. ssp. rapa) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B936 | Broccoli (Brassica oleracea L. var. italica) Turnip (Brassica rapa L. ssp. rapa) | BC1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B1018 | Broccoli (Brassica oleracea L. var. italica) Cabbage (Brassica oleracea L. var. capitata) Turnip (Brassica rapa L. ssp. rapa) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B1019 | Broccoli (Brassica oleracea L. var. italica) Cabbage (Brassica oleracea L. var. capitata) Turnip (Brassica rapa L. ssp. rapa) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B1024 | Choi Sum (Brassica campestris L. ssp. chinensis var. utilis Tsen et Lee) Chinese cabbage (Brassica pekinensis Rupr.) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B1025 | Choi Sum (Brassica campestris L. ssp. chinensis var. utilis Tsen et Lee) Chinese cabbage Chinese cabbage (Brassica pekinensis Rupr.) Turnip (Brassica rapa L. ssp. rapa) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B1026 | Broccoli (Brassica oleracea L. var. italica) Rape (Brassica napus L.) | F1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| B1027 | Broccoli (Brassica oleracea L. var. italica) Rape (Brassica napus L.) | BC1 | CAAS-IVF, Beijing, China | S, R, HR | 2021 |
| Primer Names | Loci | Primer Sequences (5′-3′) | Product Size (bp) |
|---|---|---|---|
| C2930-T-FW | CRa | TAGACCTTTTTTTTGTCTTTTTTTTTACCT | 800 |
| SC2930-R-FW | CRa | CAGACTAGACTTTTTGTCATTTAGACT | 800 |
| SC2930-RV | CRa | AAGGCCATAGAAATCAGGTC | 800 |
| KBrH129J18R-F | CRb | AGAGCAGAGTGAAACCAGAACT | 254 |
| KBrH129J18R-R | CRb | GTTTCAGTTCAGTCAGGTTTTTGCAG | 194 |
| B50-C9-FW | CRc | GATTCAATGCATTTCTCTCGAT | 800 |
| B50-6R-FW | CRc | AATGCATTTTCGCTCAACC | 800 |
| B50-RV | CRc | CGTATTATATCTCTTTCTCCATCCC | 800 |
| HC688-4-FW | CRk | TCTCTGTATTGCGTTGACTG | 1000 |
| HC688-6-RV | CRk | ATATGTTGAAGCCTATGTCT | 1000 |
| HC688-7-RV | CRk | AAATATATGTGAAGTCTTATGATC | 1000 |
| BRMS-088T | Crr1 | TATCGGTACTGATTCGCTCTTCAAC | 263 |
| BRMS-088R | Crr1 | ATCGGTTGTTATTTGAGAGCAGATT | 233 |
| BRMS-096T | Crr2 | AGTCGAGATCTCGTTCGTGTCTCCC | 220 |
| BRMS-096R | Crr2 | TGAAGAAGGATTGAAGCTGTTGTTG | 189 |
| OPC11-2ST | Crr3 | GTAACTTGGTACAGAACAGCATAG | 1300 |
| OPC11-2SR | Crr3 | ACTTGTCTAATGAATGATGATGG | 1000 |
| Primer Names | Sequences (5′-3′) | Tm (°C) | Products (bp) |
|---|---|---|---|
| Actin1 | F: GGGACATCACCGACTACCTG | 57 | 160 |
| R: ACTGCTCCGAGTTGGACATC | |||
| Novel342-2 | F: CCACGCCTATACCCGGAAAG | 58 | 666 |
| R: CAACAGGACGGCGTTGAAAG | |||
| Novel407-2 | F: GTCGTTGTTCGGGGAGAAGT | 58 | 683 |
| R: GTCCATAGGTGTGGGAACGG | |||
| PBRA_007750-2 | F: ATCTGTTCGATTCGCCTGCT | 58 | 1034 |
| R: GAGTGTACAGGCTCGCTCAG | |||
| PBRA_008439-1 | F: TCGGCGACCTGAGCGAGAA | 58 | 651 |
| R: TCAACATGCGCATAGTAC | |||
| PBRA_009348-1 | F: CACTGCTATCGTCTCCCTGG | 57 | 509 |
| R: CCTGCAATGTTTCGCTGCAA |
| Number | CRa | CRb | CRc | CRk | Crr1 | Crr2 | Crr3 |
|---|---|---|---|---|---|---|---|
| B891 | / | / | / | / | R | R | R |
| B991 | / | / | / | / | R | R | R |
| B1007 | / | / | / | / | R | / | R |
| B1081 | / | / | / | / | R | R | R |
| B1082 | / | / | / | / | R | R | R |
| B1083 | / | / | / | / | / | / | R |
| B1084 | / | / | / | / | / | / | R |
| B1086 | R | / | / | / | R | / | R |
| B359 | S | S | / | / | / | / | S |
| B366 | S | S | / | / | / | S | S |
| B368 | S | S | R | / | / | S | S |
| B369 | S | S | R | / | / | S | S |
| B571 | / | H | / | / | / | / | / |
| B578 | H/S | H/R | H/S | R | H | H/R | S |
| B581 | S | S | / | / | H | H | / |
| B582 | / | R | / | / | / | / | / |
| B606 | H/S | H | S | / | / | H/S | R |
| B607 | H/S | H | S | / | / | H | S |
| B608 | S | H | R | / | / | S | / |
| B611 | H/S | H | S | / | / | S | S |
| B612 | H/R/S | H | S | / | / | H/R/S | / |
| B613 | H/S | H | R | / | S | H/S | S |
| B614 | H/S | H/RS | R | / | S | H/R | S |
| B621 | S | S | / | / | / | S | S |
| B831 | S | S | / | / | S | S | S |
| B832 | S | S | / | / | S | S | S |
| B908 | H/S | H/RS/S | / | / | S | / | S |
| B909 | H/R/S | H/R/S | / | / | S | / | S |
| B910 | H/S | HS | / | / | S | S | R |
| B932 | H/S | S | / | / | S | / | / |
| B933 | S | S | / | / | / | S | R |
| B934 | H/S | H/S | / | / | H/S | R/S | R |
| B935 | H/R/S | H/S | / | / | H/S | H/R/S | R |
| B936 | H/S | H/S | / | / | S | S | R |
| B1018 | H/S | H/S | / | / | S | / | R |
| B1019 | H/S | H/S | / | / | S | H | R |
| B1024 | R/S | H/S | S | S | S | H | S |
| B1025 | H/R/S | H/S | S | R/S | S | S | S |
| B1026 | S | S | S | R | S | S | R/S |
| B1027 | S | S | / | / | S | S | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Q.; Wei, X.; Liu, Y.; Han, F.; Li, Z. Germplasm Enhancement and Identification of Loci Conferring Resistance against Plasmodiophora brassicae in Broccoli. Genes 2022, 13, 1600. https://doi.org/10.3390/genes13091600
Xie Q, Wei X, Liu Y, Han F, Li Z. Germplasm Enhancement and Identification of Loci Conferring Resistance against Plasmodiophora brassicae in Broccoli. Genes. 2022; 13(9):1600. https://doi.org/10.3390/genes13091600
Chicago/Turabian StyleXie, Qi, Xiaochun Wei, Yumei Liu, Fengqing Han, and Zhansheng Li. 2022. "Germplasm Enhancement and Identification of Loci Conferring Resistance against Plasmodiophora brassicae in Broccoli" Genes 13, no. 9: 1600. https://doi.org/10.3390/genes13091600