
Cytogenetic Analysis in the Toad Species Bufo spinosus, Bufotes viridis and Epidalea calamita (Anura, Bufonidae) from the Mediterranean Area


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals Analyzed
2.2. Cell Culture and Chromosome Preparations
2.3. Chromosome Banding
2.4. In Situ Hybridization: FISH and GISH
2.5. Microscopy and Image Capture
2.6. Chromosome Measurements
3. Results
3.1. C-Banding
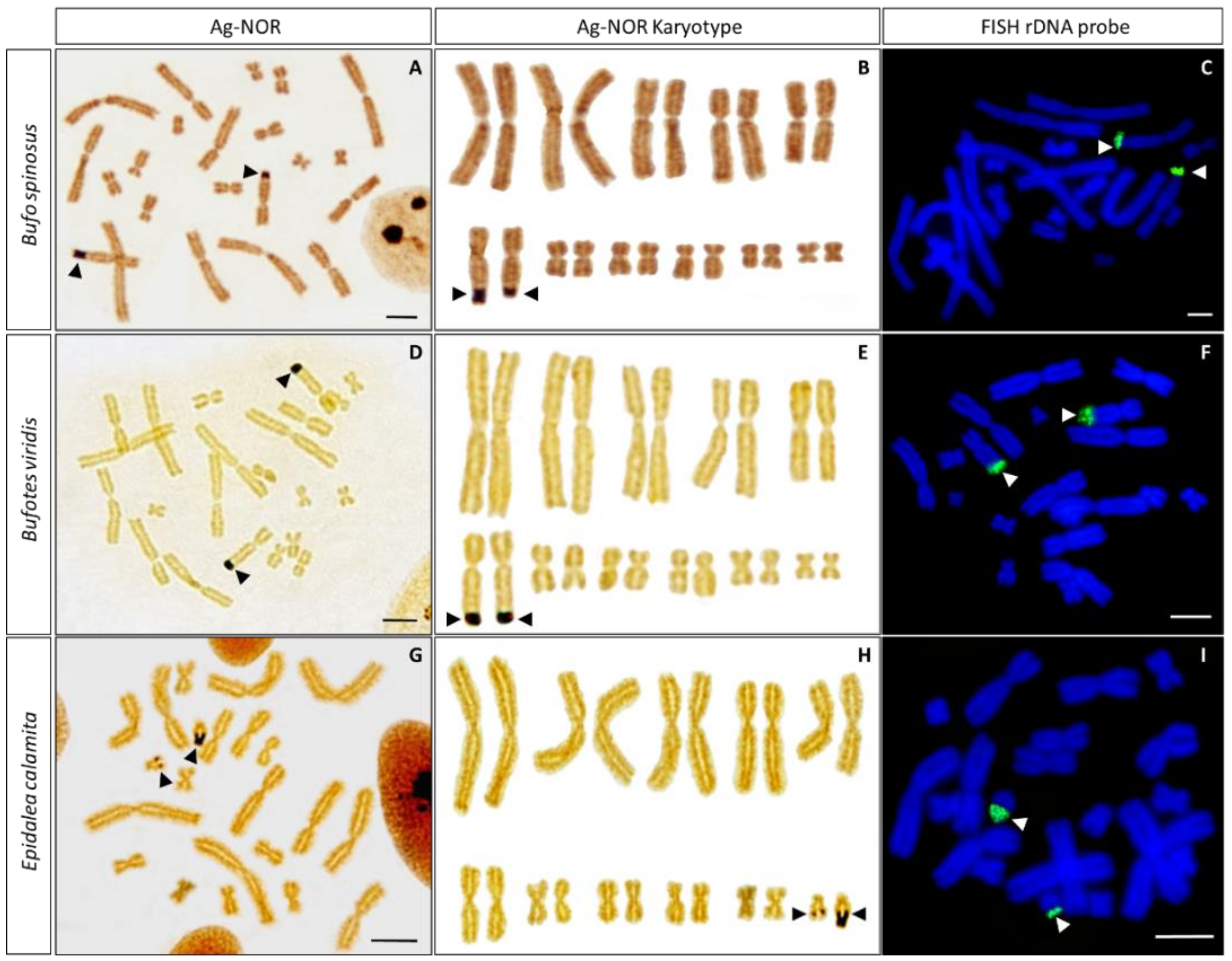
3.2. Ag-NOR Staining
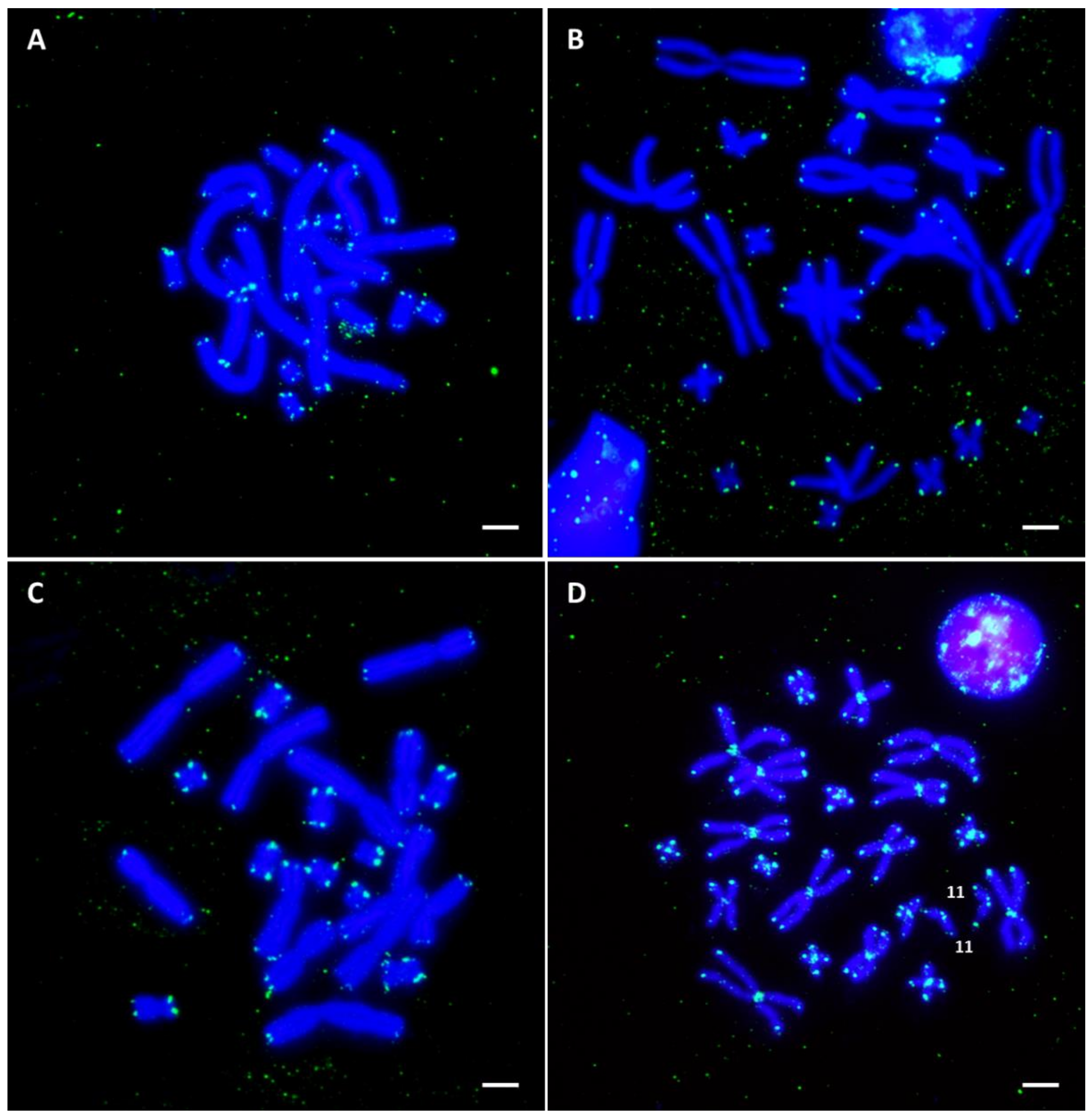
3.3. In Situ Hybridization with Ribosomal DNA
3.4. In Situ Hybridization with Telomeric Repeats
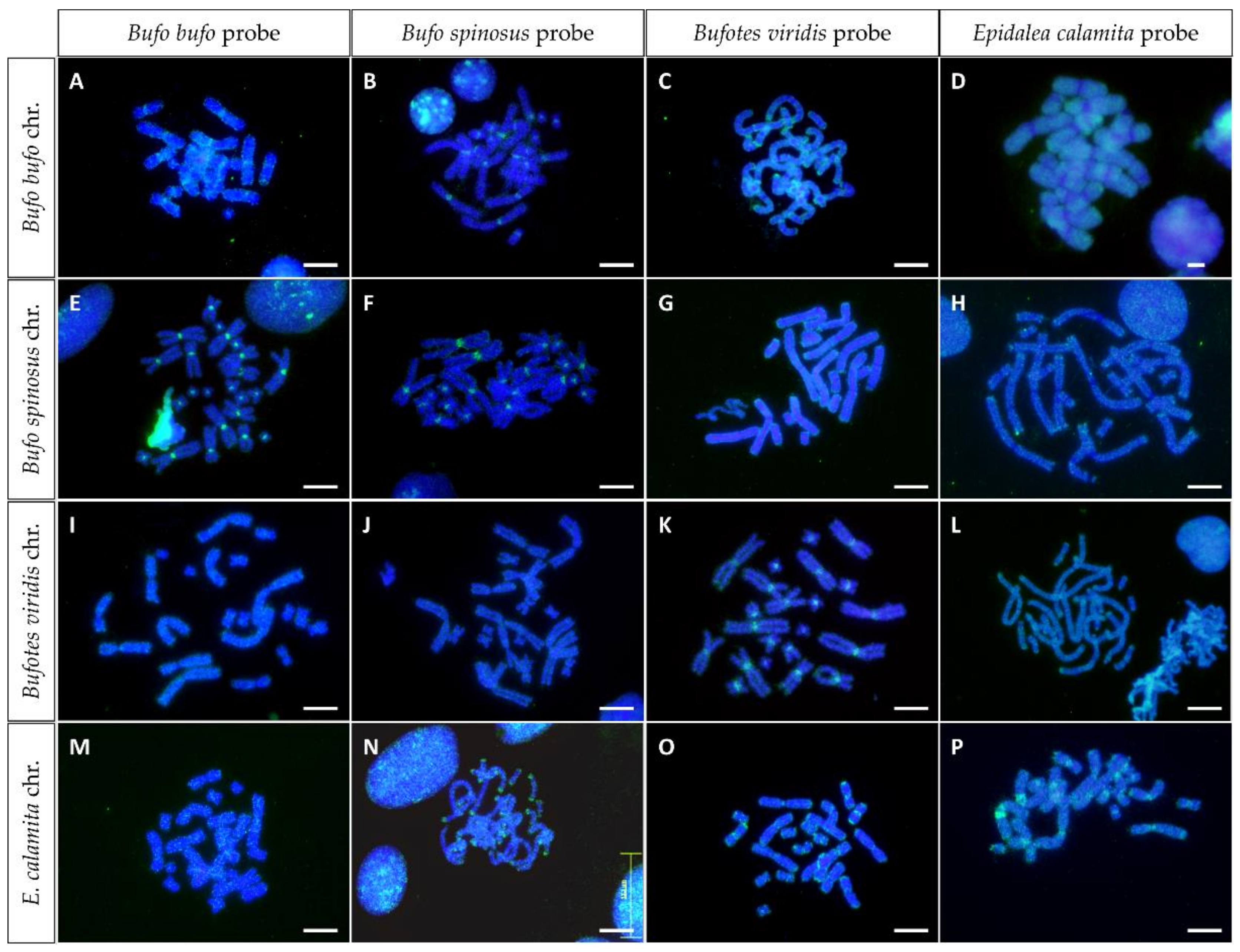
3.5. In Situ Hybridization with Genomic DNA (GISH)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- AmphibiaWeb. Available online: https://amphibiaweb.org (accessed on 14 June 2022).
- Frost, D.R. Amphibian Species of the World: An Online Reference. Version 6.1. Available online: https://amphibiansoftheworld.amnh.org/index.php (accessed on 14 June 2022).
- Integrated Taxonomic Information System (ITIS) On-Line Database. Available online: www.itis.gov (accessed on 1 May 2022).
- Frost, D.R.; Grant, R.D.; Faivovich, J.; Bain, R.H.; Haas, A.; Haddad, C.F.B.; De Sa, R.O.; Channing, A.; Wilkinson, M.; Donnellan, S.C.; et al. The amphibian tree of life. Bull. Am. Museum Nat. Hist. 2006, 371, 1–291. [Google Scholar] [CrossRef]
- Pyron, R.A.; Wiens, J.J. A large-scale phylogeny of Amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians. Mol. Phylogenet. Evol. 2011, 61, 543–583. [Google Scholar] [CrossRef] [PubMed]
- Stöck, M.; Moritz, C.; Hickerson, M.; Frynta, D.; Dujsebayeva, T.; Eremchenko, V.; Macey, J.R.; Papenfuss, T.J.; Wake, D.B. Evolution of mitochondrial relationships and biogeography of Palearctic green toads (Bufo viridis subgroup) with insights in their genomic plasticity. Mol. Phylogenet. Evol. 2006, 41, 663–689. [Google Scholar] [CrossRef]
- Frost, D.R.; Grant, T.; Faivovich, J.; Bain, R.H.; Haas, A.; Haddad, C.F.B.; De Sa, R.O.; Channing, A.; Wilkinson, M.; Donnellan, S.C.; et al. Is The Amphibian Tree of Life really fatally flawed? Cladistics 2008, 24, 385–395. [Google Scholar] [CrossRef]
- Wiens, J. Book review: The amphibian tree of life. Q. Rev. Biol. 2007, 82, 55–56. [Google Scholar] [CrossRef]
- Martínez-Solano, Í.; González, E.G. Patterns of gene flow and source-sink dynamics in high altitude populations of the common toad Bufo bufo (Anura: Bufonidae). Biol. J. Linn. Soc. 2008, 95, 824–839. [Google Scholar] [CrossRef]
- Oromi, N.; Richter-Boix, A.; Sanuy, D.; Fibla, J. Genetic variability in geographic populations of the natterjack toad (Bufo calamita). Ecol. Evol. 2012, 2, 2018–2026. [Google Scholar] [CrossRef]
- Litvinchuk, S.N.; Borkin, L.J.; Skorinov, D.V.; Rosanov, J.M. A new species of common toads from the Talysh Mountains of south-eastern Caucasus: Genome size, allozyme, and morphological evidences. Russ. J. Herpetol. 2008, 15, 19–43. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Recuero, E.; Canestrelli, D.; Martínez-Solano, Í. How complex is the Bufo bufo species group? Mol. Phylogenet. Evol. 2013, 69, 1203–1208. [Google Scholar] [CrossRef]
- Garcia-Porta, J.; Litvinchuk, S.N.; Crochet, P.A.; Romano, A.; Geniez, P.H.; Lo-Valvo, M.; Lymberakis, P.; Carranza, S. Molecular phylogenetics and historical biogeography of the west-palearctic common toads (Bufo bufo species complex). Mol. Phylogenet. Evol. 2012, 63, 113–130. [Google Scholar] [CrossRef]
- Recuero, E.; Canestrelli, D.; Vörös, J.; Szabó, K.; Poyarkov, N.A.; Arntzen, J.W.; Crnobrnja-Isailovic, J.; Kidov, A.A.; Cogălniceanu, D.; Caputo, F.P.; et al. Multilocus species tree analyses resolve the radiation of the widespread Bufo bufo species group (Anura, Bufonidae). Mol. Phylogenet. Evol. 2012, 62, 71–86. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Wilkinson, J.W.; Butôt, R.; Martínez-Solano, Í. A new vertebrate species native to the British Isles: Bufo spinosus Daudin, 1803 in Jersey. Herpetol. J. 2014, 24, 209–216. [Google Scholar]
- Hassine, J.B.; Escoriza, D. Bufo spinosus in Tunisia: New data on occurrence, parasitism and tadpole morphology. Herpetol. Bull. 2014, 127, 22–32. [Google Scholar]
- Spasic-Boskovic, O.; Lazic, T.; Blic, G.; Vujosevic, M. C-banding karyotypes and location of NOR’s in the toad species Bufo bufo and Bufo viridis from Yugoslavia. Genetika 2000, 32, 145–153. [Google Scholar]
- Skorinov, D.V.; Bolshakova, D.S.; Donaire, D.; Pasynkova, R.A.; Litvinchuk, S.N. Karyotypic analysis of the spined toad, Bufo spinosus Daudin, 1803 (Amphibia: Bufonidae). Russ. J. Herpetol. 2018, 25, 253–258. [Google Scholar] [CrossRef]
- Ullerich, F.H. Karyotyp und DNS-Gehalt von Bufo bufo, B. viridis, B. bufo × B. viridis und B. calamita (Amphibia, Anura). Chromosoma 1966, 18, 316–342. [Google Scholar] [CrossRef]
- Schmid, M. Chromosome Banding in Amphibia I. Constitutive Heterochromatin and Nucleolus Organizer Regions in Bufo and Hyla. Chromosoma 1978, 66, 361–388. [Google Scholar] [CrossRef]
- Birstein, V.J.; Mazin, A.L. Chromosomal polymorphism of Bufo bufo: Karyotype and C-banding pattern of B. b. verrucosissima. Genetica 1982, 59, 93–98. [Google Scholar] [CrossRef]
- Odierna, G.; Aprea, G.; Capriglione, T.; Castellano, S.; Balletto, E. Evidence for chromosome and PstI satellite DNA family evolutionary stasis in the Bufo viridis group (Amphibia, Anura). Chromosom. Res. 2004, 12, 671–681. [Google Scholar] [CrossRef]
- Arntzen, J.W.; McAtear, J.; Recuero, E.; Ziermann, J.M.; Ohler, A.; van Alphen, J.; Martínez-Solano, Í. Morphological and genetic differentiation of Bufo toads: Two cryptic species in Western Europe (Anura, Bufonidae). Contrib. Zool. 2013, 82, 147–169. [Google Scholar] [CrossRef]
- Stöck, M.; Sicilia, A.; Belfiore, N.M.; Buckley, D.; Lo Brutto, S.; Lo Valvo, M.; Arculeo, M. Post-Messinian evolutionary relationships across the Sicilian channel: Mitochondrial and nuclear markers link a new green toad from Sicily to African relatives. BMC Evol. Biol. 2008, 8, 56. [Google Scholar] [CrossRef]
- Guzmán, K.; Roco, Á.S.; Stöck, M.; Ruiz-García, A.; García-Muñoz, E.; Bullejos, M. Identification and characterization of a new family of long satellite DNA, specific of true toads (Anura, Amphibia, Bufonidae). Sci. Rep. 2022, 12, 13960. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- Wake, D.B.; Vredenburg, V.T. Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc. Natl. Acad. Sci. USA 2008, 105, 11466–11473. [Google Scholar] [CrossRef]
- Roco, Á.S.; Liehr, T.; Ruiz-García, A.; Guzmán, K.; Bullejos, M. Comparative distribution of repetitive sequences in the karyotypes of Xenopus tropicalis and Xenopus laevis (Anura, pipidae). Genes 2021, 12, 617. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Steinlein, C.; Bogart, J.P.; Feichtinger, W.; León, P.; La Marca, E.; Díaz, L.M.; Sanz, A.; Chen, S.H.; Hedges, S.B. The chromosomes of terraranan frogs: Insights into vertebrate cytogenetics. Cytogenet. Genome Res. 2010, 130–131, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Rufas, J.S.; Giménez-Martín, G.; Esponda, P. Presence of a chromatid core in mitotic and meiotic chromosomes of grasshoppers. Cell Biol. Int. Rep. 1982, 6, 261–267. [Google Scholar] [CrossRef]
- Endow, S.A. Polytenization of the ribosomal genes on the X and Y chromosomes of Drosophila melanogaster. Genetics 1982, 100, 375–385. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ. Available online: imagej.nih.gov/ij/ (accessed on 5 July 2018).
- Matsui, M.; Seto, T.; Kohsaka, Y.; Borkin, L.J. Bearing of chromosome C-banding patterns on the classification of Eurasian toads of Bufo bufo complex. Amphibia-Reptilia 1985, 6, 23–33. [Google Scholar] [CrossRef]
- Odierna, G.; Aprea, G.; Capriglione, T.; Castellano, S.; Balletto, E. Cytological evidence for population-specific sex chromosome heteromorphism in Palaearctic green toads (Amphibia, Anura). J. Biosci. 2007, 32, 763–768. [Google Scholar] [CrossRef]
- Skorinov, D.V.; Berezina, Y.A.; Kidov, A.A.; Matushkina, K.A.; Pasynkova, R.A.; Litvinchuk, S.N. Karyotype of the Eichwald’s Toad, Bufo eichwaldi (Amphibia: Bufonidae). Proc. Zool. Inst. RAS 2014, 318, 424–432. [Google Scholar] [CrossRef]
- Kami, H.G.; Yadollahvand, R. First kartological study of the Talysh Toad (Bufo eichwaldi) in Mazandaran province, Iran (Anura: Bufonidae). Bull. Env. Pharmacol. Life Sci. 2014, 3, 75–78. [Google Scholar]
- Matsui, M. Karyology of the Eurasian toads of the Bufo bufo complex. Annot. Zool. Jpn. Nihon Dbutsugaku Ih. Tokyo 1980, 53, 56–68. [Google Scholar]
- Miura, I. Two differentiated groups of the Japanese toad, Bufo japonicus japonicus, demonstrated by C-banding analysis of chromosomes. Caryologia 1995, 48, 123–136. [Google Scholar] [CrossRef]
- Amaro-Ghilardi, R.C.; Silva, M.J.D.J.; Rodrigues, M.T.; Yonenaga-Yassuda, Y. Chromosomal studies in four species of genus Chaunus (Bufonidae, Anura): Localization of telomeric and ribosomal sequences after fluorescence in situ hybridization (FISH). Genetica 2008, 134, 159–168. [Google Scholar] [CrossRef]
- Stöck, M.; Gunther, R.; Bohme, W. Progress towards a taxonomic revision of the Asian Bufo viridis group: Current status of nominal taxa and unsolved problems (Amphibia: Anura: Bufonidae). Zool. Abhandlungen 2001, 51, 253–319. [Google Scholar]
- Azevedo, M.F.C.; Foresti, F.; Ramos, P.R.R.; Jim, J. Comparative cytogenetic studies of Bufo ictericus, B. paracnemis (Amphibia, Anura) and an intermediate form in sympatry. Genet. Mol. Biol. 2003, 26, 289–294. [Google Scholar] [CrossRef]
- Haaf, T.; Weis, H.; Schindler, D.; Schmid, M. Specific silver staining of experimentally undercondensed chromosome regions. Chromosoma 1984, 90, 149–155. [Google Scholar] [CrossRef]
- Murray, B.G.; Bennett, M.D.; Hammett, K.R.W. Secondary constrictions and NORs of Lathyrus investigated by silver staining and in-situ hybridization. Heredity 1992, 68, 473–478. [Google Scholar] [CrossRef]
- Sudman, P.D.; Greenbaum, I.F. Visualization of kinetochores in mammalian meiotic preparations and observations of argentophilic differences between mitotic and meiotic kinetochores. Genome 1989, 32, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Abramyan, J.; Feng, C.W.; Koopman, P.A. Cloning and expression of candidate sexual development genes in the cane toad (Bufo marinus). Dev. Dyn. 2009, 238, 2430–2441. [Google Scholar] [CrossRef] [PubMed]
- Gregory, T.R.; Nicol, J.A.; Tamm, H.; Kullman, B.; Kullman, K.; Leitch, I.J.; Murray, B.G.; Kapraun, D.F.; Greilhuber, J.; Bennett, M.D. Eukaryotic genome size databases. Nucleic Acids Res. 2007, 35, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Baldari, C.T.; Amaldi, F. DNA reassociation kinetics in relation to genome size in four amphibian species. Chromosoma 1976, 59, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Meyne, J.; Baker, R.J.; Hobart, H.H.; Hsu, T.C.; Ryder, O.A.; Ward, O.G.; Wiley, J.E.; Wurster-Hill, D.H.; Yates, T.L.; Moyzis, R.K. Distribution of non-telomeric sites of the (TTAGGG)n telomeric sequence in vertebrate chromosomes. Chromosoma 1990, 99, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Calixto, M.; de Andrade, I.S.; Cabral-de-Mello, D.C.; Santos, N.; Martins, C.; Loreto, V.; de Souza, M.J. Patterns of rDNA and telomeric sequences diversification: Contribution to repetitive DNA organization in Phyllostomidae bats. Genetica 2014, 142, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Marchal, J.A.; Giagia-Athanasopoulou, E.; Sánchez, A. Molecular composition of heterochromatin and its contribution to chromosome variation in the microtus thomasi/microtus atticus species complex. Genes 2021, 12, 807. [Google Scholar] [CrossRef]
- Clemente, L.; Mazzoleni, S.; Bellavia, E.P.; Augstenová, B.; Auer, M.; Praschag, P.; Protiva, T.; Velenský, P.; Wagner, P.; Fritz, U.; et al. Interstitial telomeric repeats are rare in turtles. Genes 2020, 11, 657. [Google Scholar] [CrossRef]
- Schneider, C.H.; Gross, M.C.; Terencio, M.L.; Artoni, R.F.; Vicari, M.R.; Martins, C.; Feldberg, E. Chromosomal evolution of neotropical cichlids: The role of repetitive DNA sequences in the organization and structure of karyotype. Rev. Fish Biol. Fish. 2013, 23, 201–214. [Google Scholar] [CrossRef]
- Nanda, I.; Fugate, M.; Steinlein, C.; Schmid, M. Distribution of (TTAGGG)n telomeric sequences in karyotypes of the Xenopus species complex. Cytogenet. Genome Res. 2008, 122, 396–400. [Google Scholar] [CrossRef]
- Bolzán, A.D. Interstitial telomeric sequences in vertebrate chromosomes: Origin, function, instability and evolution. Mutat. Res. 2017, 773, 51–65. [Google Scholar] [CrossRef]
- Suárez, P.; Cardozo, D.; Baldo, D.; Pereyra, M.O.; Faivovich, J.; Orrico, V.G.D.; Catroli, G.F.; Grabiele, M.; Bernarde, P.S.; Nagamachi, C.Y.; et al. Chromosome evolution in dendropsophini (amphibia, anura, hylinae). Cytogenet. Genome Res. 2013, 141, 295–308. [Google Scholar] [CrossRef]
- Nergadze, S.G.; Santagostino, M.A.; Salzano, A.; Mondello, C.; Giulotto, E. Contribution of telomerase RNA retrotranscription to DNA double-strand break repair during mammalian genome evolution. Genome Biol. 2007, 8, R260. [Google Scholar] [CrossRef]
- Bruschi, D.P.; Rivera, M.; Lima, A.P.; Zúñiga, A.B.; Recco-Pimentel, S.M. Interstitial Telomeric Sequences (ITS) and major rDNA mapping reveal insights into the karyotypical evolution of Neotropical leaf frogs species (Phyllomedusa, Hylidae, Anura). Mol. Cytogenet. 2014, 7, 22. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Origin | Ploidy | Sex | Dev. St. | Sample ID | Chr. Source | C-Band | Ag-NOR 1 (% HT) | FISH rDNA | TEL |
|---|---|---|---|---|---|---|---|---|---|---|
| B. bufo | Bologna (Italy) | 2n = 22 | F | A | H6 | BM | - | - | - | 7 |
| B. spinosus | Jaén (Spain) | 2n = 22 | M | T | T8 (P5) | CC | - | 14 (78.6%) | - | - |
| F | T | T10 (P4) | CC | 39 | 94 (71.3%) | - | - | |||
| F | T | T12 (P2/P6/P9/P10) | CC | 64 | 80 (88.7%) | 5 | 18 | |||
| B. viridis | Crete (Greece) | 2n = 22 | UK | T | R14 (P5/P6) | CC | 68 | 55 (32.7%) | 1 | 6 |
| UK | T | R15 (P4) | CC | - | - | 4 | 11 | |||
| E. calamita | Jaén (Spain) | 2n = 22 | UK | T | T2 (P21) | CC | 3 | 162 (71.0%) | - | 23 |
| F | T | T3.1 (P17) | CC | 14 | 22 (72.7%) | - | - | |||
| F | T | T3.2 (P10/P15/P16/P19) | CC | 240 | 170 (74.1%) | 4 | 69 | |||
| M | T | T4.4 (P18/P19) | CC | - | 127 (81.9%) | - | - | |||
| M | A | M4 | BM | - | - | - | 28 |
| Sample ID | N | Sex | Lp(6pA-6pB) | Lq(6qA-6qB) | L(6A-6B) |
|---|---|---|---|---|---|
| T8 | 12 | Male | 7.477 ± 4.292 | 6.748 ± 6.325 | 5.462 ± 3.174 |
| T10 | 18 | Female | 4.612 ± 4.105 | 5.452 ± 3.995 | 4.524 ± 2.579 |
| T12 | 67 | Female | 5.883 ± 5.101 | 7.527 ± 5.104 | 5.168 ± 4.509 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzmán-Markevich, K.; Roco, Á.S.; Ruiz-García, A.; Bullejos, M. Cytogenetic Analysis in the Toad Species Bufo spinosus, Bufotes viridis and Epidalea calamita (Anura, Bufonidae) from the Mediterranean Area. Genes 2022, 13, 1475. https://doi.org/10.3390/genes13081475
Guzmán-Markevich K, Roco ÁS, Ruiz-García A, Bullejos M. Cytogenetic Analysis in the Toad Species Bufo spinosus, Bufotes viridis and Epidalea calamita (Anura, Bufonidae) from the Mediterranean Area. Genes. 2022; 13(8):1475. https://doi.org/10.3390/genes13081475
Chicago/Turabian StyleGuzmán-Markevich, Katerina, Álvaro S. Roco, Adrián Ruiz-García, and Mónica Bullejos. 2022. "Cytogenetic Analysis in the Toad Species Bufo spinosus, Bufotes viridis and Epidalea calamita (Anura, Bufonidae) from the Mediterranean Area" Genes 13, no. 8: 1475. https://doi.org/10.3390/genes13081475