
Vacuolal and Peroxisomal Calcium Ion Transporters in Yeasts and Fungi: Key Role in the Translocation of Intermediates in the Biosynthesis of Fungal Metabolites

Abstract
:Highlights
- The intracellular calcium content plays a key role in the expression of genes involved in the biosynthesis and secretion of fungal metabolites.
- The cytosolic calcium concentration in fungi is maintained by influx through the cell membrane and by release from store organelles.
- Some MSF transporters, e.g., PenV of Penicillium chrysogenum and CefP of Acremonium chrysogenum belong to the TRP calcium ion channels.
- A few of the numerous calcium ion transporters existing in organelles of different filamentous fungi have been characterized at the functional and subcellular localization levels.
- The cytosolic calcium signal seems to be transduced by the calcitonin/calcineurin cascade controlling the expression of many fungal genes.
Abstract
1. Introduction
2. Role of Calcium in Subcellular Transport Mechanisms
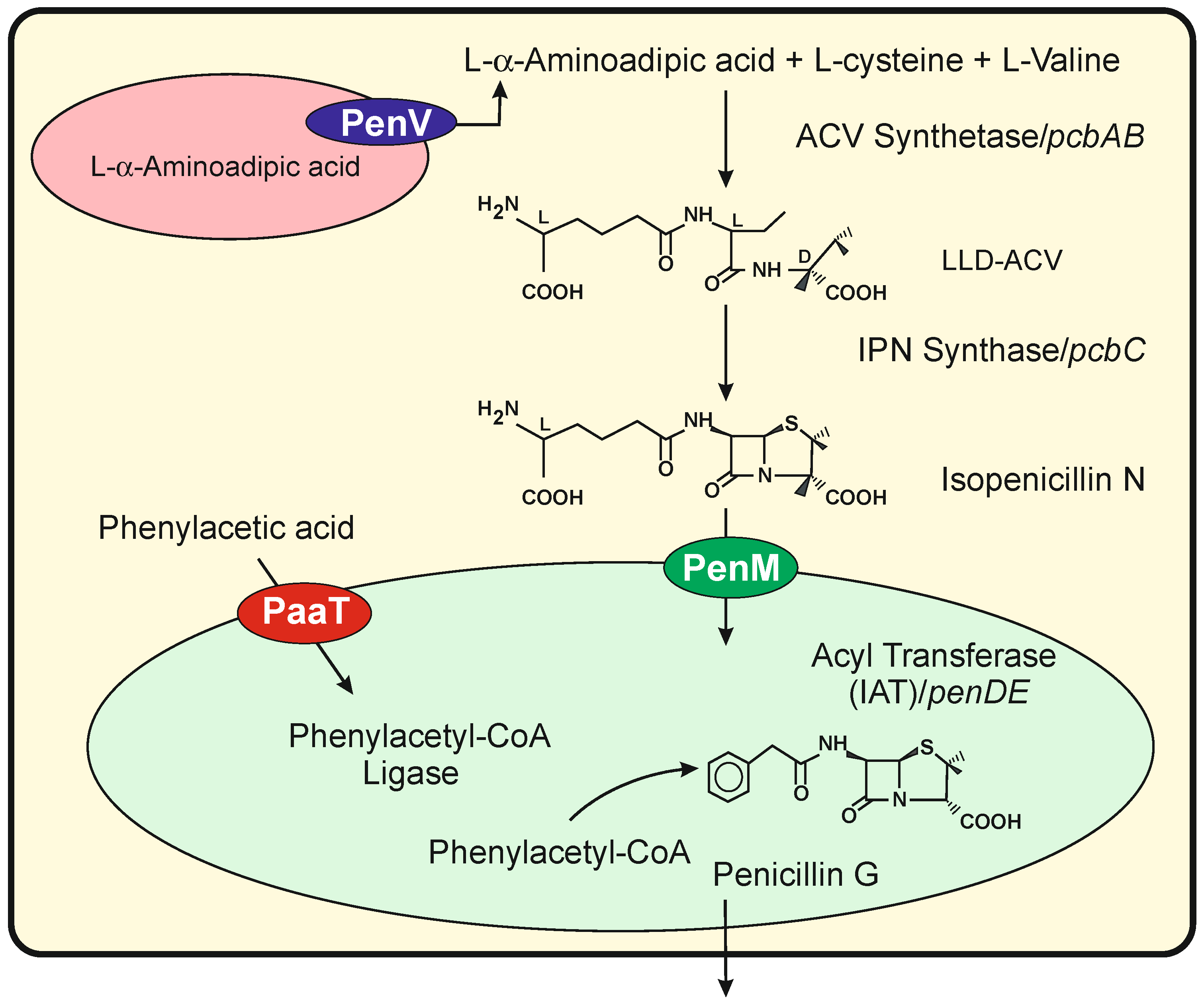
3. Numerous Transporters Are Involved in the Translocation of Fungal Intermediates of Different Specialized Metabolites
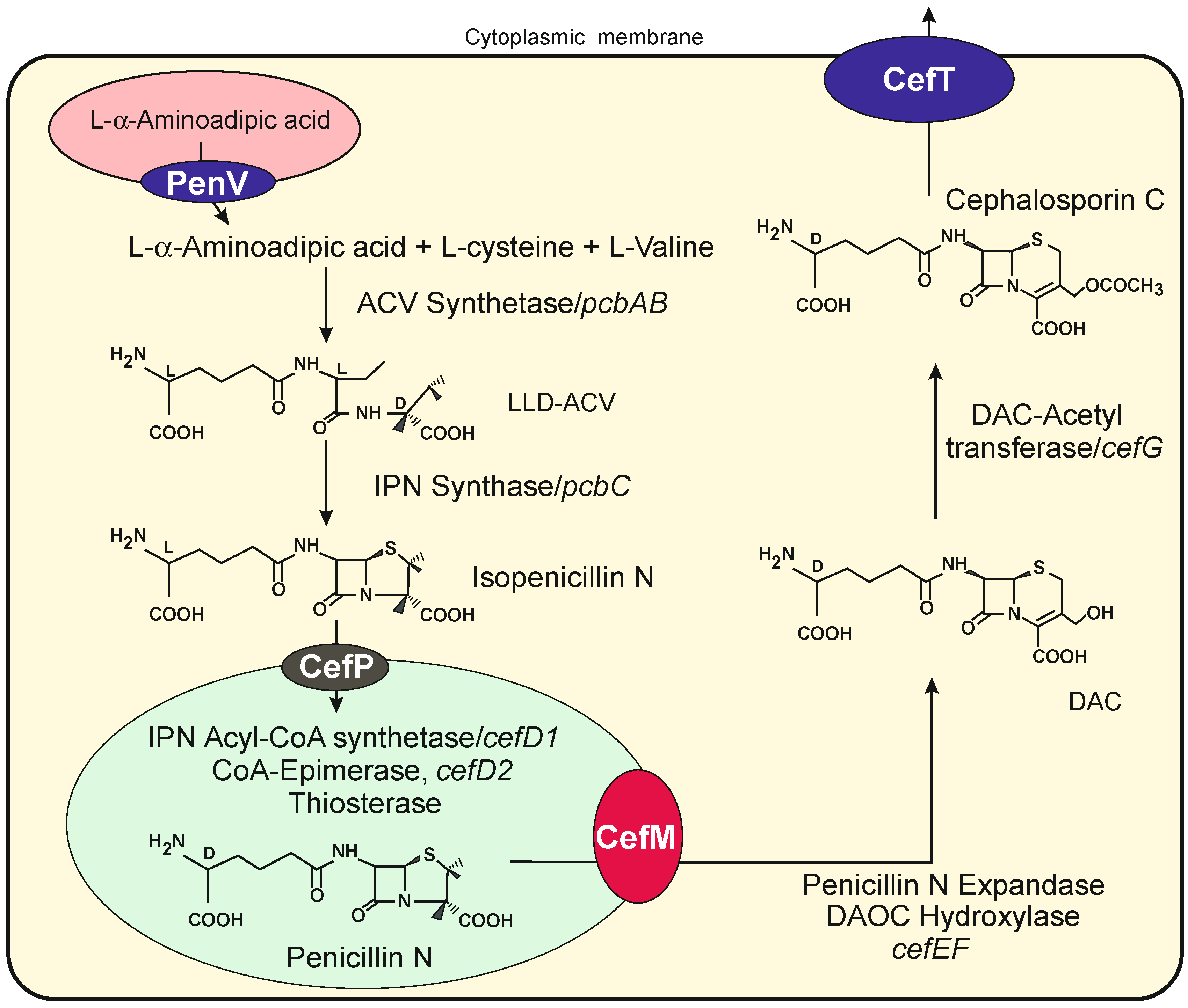
Tranporters Encoded by Genes in the Cephalosporin Gene Cluster
4. Transient Receptor Potential Ion Channels in Fungal Metabolism
5. The Model Vcx1 Ion Exchangers of S. cerevisiae: Comparison with Homologous Proteins in Filamentous Fungi
5.1. A family of Multiple Vcx1-like Calcium Transporters in Filamentous Fungi: Characterization of a Protein of P. chrysogenum Homologous to the Vcx1 of S. cerevisiae
5.2. Characteristics and Localization of the Vcx1-like Transporters of Cryptococcus neoformans and Neurospora crassa
6. Efflux Vacuole Calcium Ion Exchangers That Release Calcium from the Vacuoles to the Cytosol: The Model Yvc1 Transporter in Yeast and Comparison with Filamentous Fungi
7. A Third Class of Calcium Ion Transporters in Filamentous Fungi Belong to the TRP Family
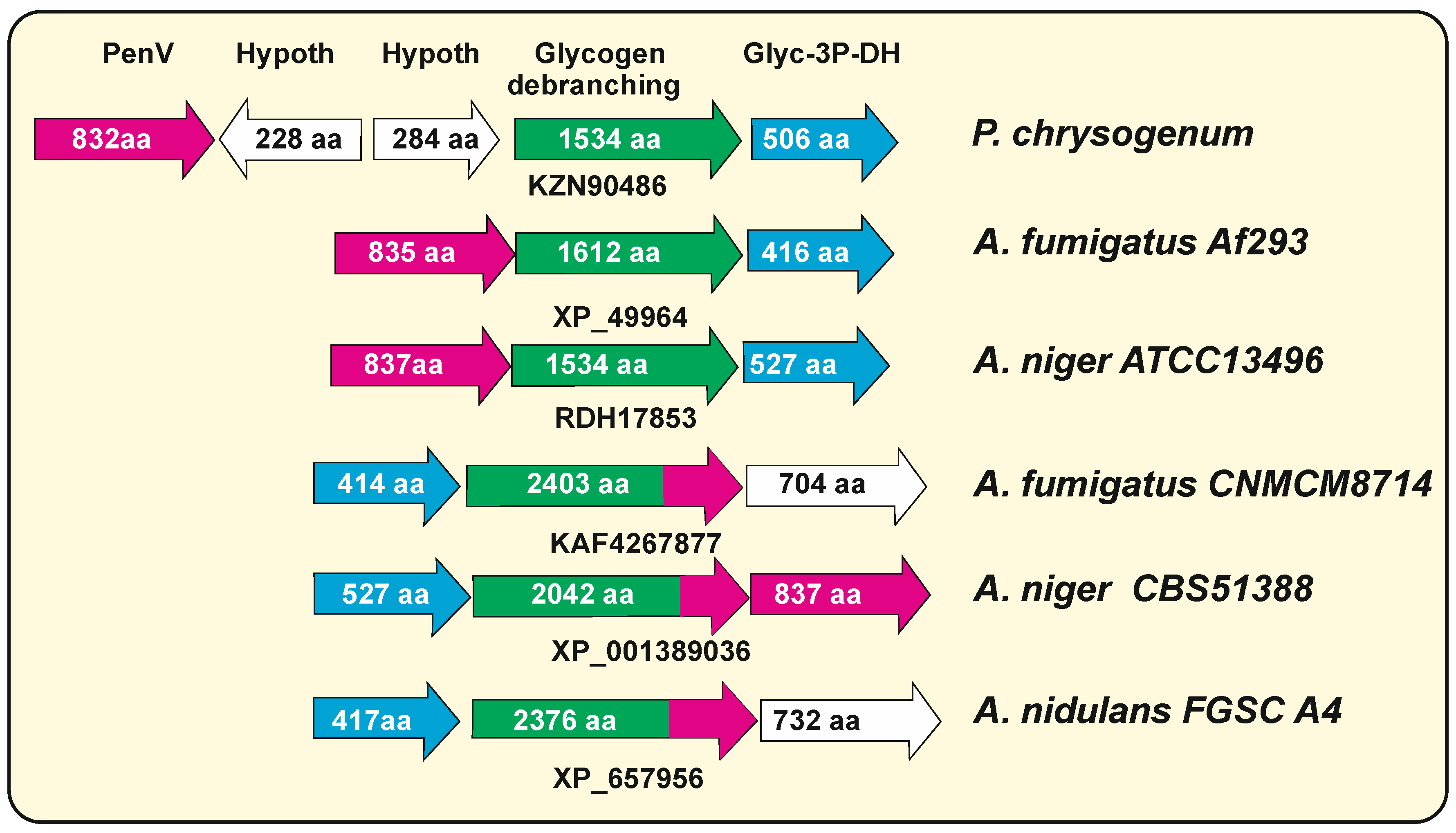
7.1. The P. chrysogenum penV Gene Belong to the TRP-CSC Family
7.2. Hybrid Glycogen Debranching Enzymes—Calcium-Permeable Stress-Gated Transporters in Aspergillus Species
8. Ion Transporters in Peroxisomes of Filamentous Fungi
The Peroxisomal CefP Transporter Also Belongs to the CSC-TRP Ion Channel Family
9. Downstream Transduction of the Calcium Level Signal
10. Conclusions and Future Outlook
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Case, R.M.; Eisner, D.; Gurney, A.; Jones, O.; Muallen, S.; Verkhratsky, A. Evolution of calcium homeostasis: From bith of the first cell to omnipresent signalling system. Cell Calcium. 2007, 42, 345–350. [Google Scholar] [CrossRef]
- Martín, J.F.; Liras, P. The PenV vacuolal membrane protein that controls penicillin biosynthesis is a putative member of a subfamily of stress-gated transient receptor calcium channels. Curr. Res. Biotechnol. 2021, 3, 317–322. [Google Scholar] [CrossRef]
- Zelter, A.; Bencina, M.; Bowman, B.J.; Yarden, O.; Read, N.D. A comparative genomic analysis of the calcium signaling machinery in Neurospora crassa, Magnaporthe grisea and Saccharomyces cerevisiae. Fungal Genet. Biol. 2004, 41, 827–841. [Google Scholar] [CrossRef] [PubMed]
- Silverman-Gavrila, L.; Lew, R.R. Calcium and tip growth in Neurospora crassa. Protoplasma 2000, 213, 203–217. [Google Scholar] [CrossRef]
- Silverman-Gavrila, L.; Lew, R.R. Regulation of the tip-high (Ca2+) gradient in growing hyphae of the fungus Neurospora crassa. Eur. J. Cell Biol. 2001, 80, 379–390. [Google Scholar] [CrossRef]
- Takeshita, N.; Evangelinos, M.; Zhou, L.; Serizawa, T.; Somera-Fajardo, R.A.; Lu, L.; Takaya, N.; Nienhaus, U.; Fischer, R. Pulses of Ca2+ coordinate actin assembly and exocytosis for stepwise cell extension. Proc. Natl. Acad. Sci. USA 2017, 114, 5701–5706. [Google Scholar] [CrossRef] [PubMed]
- Parekh, A.B. Decoding cytosolic Ca2+ oscillations. Trends Biochem. Sci. 2011, 36, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Santos, R.; Kosalkova, K.; García-Estrada, C.; Barreiro, C.; Ibañez, A.; Morales, A.; Martín, J.F. Casein Phosphopeptides and CaCl2 increase penicillin production and cause an increment in microbody/peroxisome protein in Penicillium chrysogenum. J. Proteomics 2017, 156, 52–62. [Google Scholar] [CrossRef]
- Martín, J.F. Calcium-containing phosphopeptides pave the secretory pathway for efficient protein traffic and secretion in fungi. Microb. Cell Fact. 2014, 13, 117. [Google Scholar] [CrossRef]
- Lasorsa, F.M.; Pinton, P.; Palmieri, L.; Scarcia, P.; Rottensteiner, H.; Rizzuto, R.; Palmieri, F. Peroxisomes as novel players in cell calcium homeostasis. J. Biol. Chem. 2008, 283, 15300–15308. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Drago, I.; Zottini, M.; Pizzo, P.; Pozzan, T. Peroxisome Ca(2+) homeostasis in animal and plant cells. Subcell. Biochem. 2013, 69, 111–133. [Google Scholar] [PubMed]
- Ullán, R.V.; Teijeira, F.; Guerra, S.M.; Vaca, I.; Martín, J.F. Characterization of a novel peroxisome membrane protein essential for conversion of isopenicillin N into cephalosporin C. Biochem. J. 2010, 432, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Aguado, M.; Teijeira, F.; Martín, J.F.; Ullán, R.V. A vacuolal membrane protein affects drastically the biosynthesis of the ACV tripeptide and the beta-lactam pathway of Penicillium chrysogenum. Appl. Microbiol. Biotechnol. 2012, 97, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Aguado, M.; Ullán, R.V.; Teijeira, F.; Rodríguez-Castro, R.; Martín, J.F. The transport of phenylacetic acid across the peroxisomal membrane is mediated by the PaaT protein in Penicillium chrysogenum. Appl. Microbiol. Biotechnol. 2013, 97, 3073–3084. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F. Transport systems, intracellular traffic of intermediates and secretion of β-lactam antibiotics in fungi. Fungal Biol. Biotechnol. 2020, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Skellam, E. Subcellular localization of fungal specialized metabolites. Fungal Biol. Biotechnol. 2022, 9, 11. [Google Scholar] [CrossRef]
- Harris, D.M.; Westerlaken, I.; Schipper, D.; van der Krogt, Z.A.; Gombert, A.K.; Sutherland, J.; Raamsdonk, L.M.; van den Berg, M.A.; Bovenberg, R.A.L.; Pronk, J.T.; et al. Engineering of Penicillium chrysogenum for fermentative production of a novel carbamoylated cephem antibiotic precursor. Metab. Eng. 2009, 11, 125–137. [Google Scholar] [CrossRef]
- García-Estrada, C.; Martín, J.F. Penicillins. In Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites; Martín, J.F., García Estrada, C., Zeilinger, S., Eds.; Springer: New York, NY, USA, 2014; pp. 17–42. [Google Scholar]
- Martín, J.F.; Liras, P. Insights into the structure and Molecular Mechanisms of β-lactam synthesizing enzymes in fungi. In Biotechnology of Microbial Enzymes; Brahmachati, G., Demain, A.L., Adrio, J.L., Eds.; Elsevier: New York, NY, USA, 2017; pp. 215–241. [Google Scholar]
- Martín, J.F. Molecular control of expression of penicillin biosynthesis genes in fungi: Regulatory proteins interact with a bidirectional promoter region. J. Bacteriol. 2000, 182, 2355–2362. [Google Scholar] [CrossRef] [PubMed]
- Cepeda-García, C.; Domínguez-Santos, R.; García-Rico, R.O.; García-Estrada, C.; Cajiao, A.; Fierro, F.; Martín, J.F. Direct involvement of the CreA transcription factor in penicillin biosynthesis and expression of the pcbAB gene in Penicillium chrysogenum. Appl. Microbiol. Biotechnol. 2014, 98, 7113–7124. [Google Scholar] [CrossRef] [PubMed]
- Evers, M.E.; Trip, H.; van der Berg, M.A.; Bovenberg, R.A.; Driessen, A.J. Compartmentalization and transport in beta-lactam antibiotics biosynthesis. Adv. Biochem. Eng. Biotechnol. 2004, 88, 111–135. [Google Scholar] [PubMed]
- Martín, J.F.; Ullán, R.V.; García-Estrada, C. Regulation and compartmentalization of β-lactam biosynthesis. Microb. Biotechnol. 2010, 3, 285–299. [Google Scholar] [CrossRef]
- Van Der Lende, T.R.; Breeuwer, P.; Abee, T.; Konings, W.N.; Driessen, A.J.M. Assessment of the microbody luminal pH in the filamentous fungus Penicillium chrysogenum. Biochim. Biophys Acta 2002, 1589, 104–111. [Google Scholar] [PubMed]
- Lendenfeld, T.; Ghali, D.; Wolschek, M.; Kubicek-Pranz, E.M.; Kubicek, C.P. Subcellular compartmentation of penicillin biosynthesis in Penicillium chrysogenum. The amino acid precursors are derived from the vacuole. J. Biol. Chem. 1993, 268, 665–671. [Google Scholar] [CrossRef]
- Lamas-Maceiras, M.; Vaca, I.; Rodríguez, E.; Casqueiro, J.; Martín, J.F. Amplification and disruption of the phenylacetyl-CoA ligase gene of Penicillium chrysogenum encoding an aryl-capping enzyme that supplies phenylacetic acid to the isopenicillin N acyltransferase. Biochem. J. 2006, 395, 147–155. [Google Scholar] [CrossRef] [PubMed]
- García-Estrada, C.; Vaca, I.; Fierro, F.; Sjollema, K.; Veenhuis, M.; Martín, J.F. The unprocessed preprotein form IATC103S of the isopenicillin N acyltransferase is transported inside peroxisomes and regulates its self-processing. Fungal. Genet. Biol. 2008, 45, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Koetsier, M.J.; Jekel, P.A.; van den Berg, M.A.; Bovenberg, R.A.; Janssen, D.B. Characterization of a phenylacetate-CoA ligase from Penicillium chrysogenum. Biochem. J. 2010, 417, 467–476. [Google Scholar] [CrossRef]
- Yu, Z.L.; Liu, J.; Wang, F.Q.; Dai, M.; Zhao, B.H.; He, J.G.; Zhang, H. Cloning and characterization of a novel CoA-ligase gene from Penicillium chrysogenum. Folia Microbiol. 2011, 56, 246–252. [Google Scholar] [CrossRef]
- García-Estrada, C.; Barreiro, C.; Jami, M.S.; Martín-González, J.; Martín, J.F. The inducers 1,3-diaminopropane and spermidine cause the reprogramming of metabolism in Penicillium chrysogenum, leading to multiple vesicles and penicillin overproduction. J. Proteomics 2013, 85, 129–159. [Google Scholar] [CrossRef]
- Fernández-Aguado, M.; Martín, J.F.; Rodríguez-Castro, R.; García-Estrada, C.; Albillos, S.M.; Teijeira, F.; Ullán, R.V. New insights into the isopenicillin N transport in Penicillium chrysogenum. Metabol. Eng. 2014, 22, 89–103. [Google Scholar] [CrossRef]
- Yang, J.; Xu, X.; Liu, G. Amplification of an MFS transporter encoding gene penT significantly stimulates penicillin production and enhances the sensitivity of Penicillium chrysogenum to phenylacetic acid. J. Genet. Genom. 2012, 39, 593–602. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Fierro, F.; Casqueiro, J.; Martín, J.F. Gene organization and plasticity of the β-lactam genes in different filamentous fungi. Antonie Van Leeuwenhoek 1999, 75, 81–94. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Díez, B.; Álvarez, E.; Barredo, J.L.; Martín, J.F. Expression of the penDE gene of Penicillium chrysogenum encoding isopenicillin N acyltransferase in Cephalosporium acremonium: Production of benzylpenicillin by the transformants. Mol. Gen. Genet. 1991, 225, 56–64. [Google Scholar] [CrossRef]
- Ullán, R.V.; Casqueiro, J.; Bañuelos, O.; Fernández, F.J.; Gutiérrez, S.; Martín, J.F. A novel epimerization system in fungal secondary metabolism involved in the conversion of isopenicillin N into penicillin N in Acremonium chrysogenum. J. Biol. Chem. 2002, 277, 46216–46225. [Google Scholar] [CrossRef]
- Martín, J.F.; Ullán, R.V.; Casqueiro, J. Novel genes involved in cephalosporin biosynthesis: The three-component isopenicillin N epimerase system. Adv. Biochem. Eng. Biotechnol. 2004, 88, 91–109. [Google Scholar]
- Kiel, J.A.; van den Berg, M.A.; Fusetti, F.; Poolman, B.; Bovenberg, R.A.; Veenhuis, M.; van der Klei, I.J. Matching the proteome to the genome: The microbody of penicillin-producing Penicillium chrysogenum cells. Funct. Integr. Genom. 2009, 9, 167–184. [Google Scholar] [CrossRef]
- Teijeira, F.; Ullán, R.V.; Guerra, S.M.; García-Estrada, C.; Vaca, I.; Martín, J.F. The transporter CefM involved in translocation of biosynthetic intermediates is essential for cephalosporin production. Biochem. J. 2009, 418, 113–124. [Google Scholar] [CrossRef]
- Rottensteiner, H.; Kramer, A.; Lorenzen, S.; Stein, K.; Landgraf, C.; Volkmer-Engert, R.; Erdmann, R. Peroxisomal membrane proteins contain common Pex19p-binding sites that are an integral part of their targeting signals. Mol. Biol. Cell. 2004, 15, 3406–3417. [Google Scholar] [CrossRef]
- Ullán, R.V.; Liu, G.; Casqueiro, J.; Gutiérrez, S.; Bañuelos, O.; Martín, J.F. The cefT gene of Acremonium chrysogenum C10 encodes a putative multidrug efflux pump protein that significantly increases cephalosporin C production. Mol. Genet. Genom. 2002, 267, 673–683. [Google Scholar] [CrossRef]
- Nijland, J.G.; Kovalchuk, A.; van den Berg, M.; Bovenberg, R.A.L.; Driessen, A.J.M. Expression of the transporter encoded by the cefT gene of Acremonium chrysogenum increases cephalosporin production in Penicillium chrysogenum. Fungal Genet. Biol. 2008, 45, 1415–1421. [Google Scholar] [CrossRef]
- Peiter, E. The Ever-Closer Union of Signals: Propagating Waves of Calcium and ROS Are Inextricably Linked. Plant Physiol. 2016, 172, 3–4. [Google Scholar] [CrossRef]
- Jiang, L.; Yang, Y. The putative transient receptor potential channel protein encoded by the orf19.4805 gene is involved in cation sensitivity, antifungal tolerance, and filamentation in Candida albicans. Can. J. Microbiol. 2018, 64, 727–731. [Google Scholar]
- Venkatachalam, K.; Montell, C. TRP channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef]
- Voolstra, O.; Huber, A. Post-translational modifications of TRP channels. Cells 2014, 3, 258–287. [Google Scholar] [CrossRef]
- Jiang, L.; Pang, H. Functions of CaPhm7 in the regulation of ion homeostasis drug tolerance, filamentation and virulence in Candida albicans. BMC Microbiol. 2018, 18, 49. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, J.E.; Frailey, D.; Nohe, A.; Dunca, R.; Czymmek, K.J.; Kang, S. Roles of three Fusarium oxysporum calcium ion (Ca2+) channels in generating Ca2+ signatures and controlling growth. Fungal Genet. Biol. 2015, 82, 145–157. [Google Scholar] [CrossRef]
- Veses, V.; Richards, A.; Gow, N.A.R. Vacuoles and fungal biology. Curr. Opin. Microbiol. 2008, 11, 503–510. [Google Scholar] [CrossRef]
- Pittman, J.K. Vacuolal Ca(2+) uptake. Cell Calcium 2011, 50, 139–146. [Google Scholar] [CrossRef]
- Tamuli, R.; Kumar, R.; Srivastava, D.A.; Deka, R. Calcium signaling. In Neurospora: Genomics and Molecular Biology; Kasbekar, D.P., McCluskey, K., Eds.; Caister Academic Press: Norfolk, VA, USA, 2013; pp. 209–226. [Google Scholar]
- Brini, M.; Carafoli, E. Calcium pumps in health and disease. Physiol. Rev. 2009, 89, 1341–1378. [Google Scholar] [CrossRef]
- Cunningham, K.W.; Fink, G.R. Calcineurin inhibits VCX1-dependent H+/Ca2+ exchange and induces Ca2+ ATPases in Saccharomyces cerevisiae. Mol. Cell. Biol. 1996, 16, 2226–2237. [Google Scholar] [CrossRef] [PubMed]
- Miseta, A.; Kellermayer, R.; Aiello, D.P.; Fu, L.W.; Bedwell, D.M. The vacuolal Ca2+/H+ exchanger Vcx1/Hum1p tightly controls cytosolic Ca2+ levels in S. cerevisiae. FEBS Lett. 1999, 451, 132–136. [Google Scholar] [CrossRef]
- Cai, X.; Lytton, J. The cation/Ca2+ exchanger superfamily: Phylogenetic analysis and structural implications. Mol. Biol. Evol. 2004, 21, 1692–1703. [Google Scholar] [CrossRef] [PubMed]
- Shigaki, T.; Barkla, B.J.; Miranda-Vergara, M.C.; Zhao, J.; Pantoja, O.; Hirschi, K.D. Identification of a crucial histidine involved in metal transport activity in the Arabidopsis cation/H+ exchanger CAX1. J. Biol. Chem. 2005, 280, 30136–30142. [Google Scholar] [CrossRef] [PubMed]
- Shigaki, T.; Rees, I.; Nakhleh, L.; Hirschi, K.D. Identification of three distinct phylogenetic groups of CAX cation/proton antiporters. J. Mol. Evol. 2006, 63, 815–825. [Google Scholar] [CrossRef]
- Pozos, T.C.; Sekler, I.; Cyert, M.S. The product of HUM1, a novel yeast gene is required for vacuolal Ca2+/H+ exchange and is related to mammalian Na+/Ca2+ exchangers. Mol. Cell Biol. 1996, 16, 3730–3741. [Google Scholar] [CrossRef]
- Segarra, V.A.; Thomas, L. Topology of the vacuolal Vcx1 Ca2+/H+ exchanger from Saccharomyces cerevisiae. Biochem. J. 2008, 414, 133–141. [Google Scholar] [CrossRef]
- Kim, H.; Melen, K.; Osteberg, M.; von Heijne, G. A global topology map of the Saccharomyces cerevisiae membrane proteome. Proc. Natl. Acad. Sci. USA 2006, 103, 11142–11147. [Google Scholar] [CrossRef]
- Chang, Y.; Schlenstedt, G.; Flockerzi, V.; Beck, A. Properties of the intracellular transient receptor potential (TRP) channel in yeast, Yvc1. FEBS Lett. 2010, 584, 2028–2032. [Google Scholar] [CrossRef]
- Hou, C.; Tian, W.; Kleist, T.; He, K.; García, V.; Bai, F.; Hao, Y.; Luan, S.; Li, L. DUF221 proteins are a family of osmosensitive calcium-permeable cation channels conserved across eukaryotes. Cell Res. 2014, 24, 632–635. [Google Scholar] [CrossRef]
- Ihara, M.; Hamamoto, S.; Miyanoiri, Y.; Takeda, M.; Kainosho, M.; Yabe, I.; Uozumi, N.; Yamashita, A. Molecular bases of multimodal regulation of a fungal transient receptor potential (TRP) channel. J. Biol. Chem. 2013, 288, 15303–15317. [Google Scholar] [CrossRef] [PubMed]
- Kmetzsch, L.; Staats, C.C.; Simon, E.; Fonseca, F.I.; de Oliveira, D.L.; Sobrino, L.; Rodrigues, J.; Leal, A.L.; Nimrichter, L.; Rodrigues, M.L.; et al. The vacuolal Ca2+ exchanger Vcx1 is involved in calcineurin-dependent Ca2+ tolerance and virulence in Cryptococcus neoformans. Eukaryotic Cells 2010, 9, 1798–1805. [Google Scholar] [CrossRef] [PubMed]
- Sá-Correia, I.; dos Santos, S.C.; Teixeira, M.C.; Cabrito, T.R.; Mira, N.P. Drug:H+ antiporters in chemical stress response in yeast. Trends Microbiol. 2009, 17, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Bowman, B.J.; Draskovic, M.; Freitag, M.; Bowman, E.J. Structure and distribution of organelles and cellular location of calcium transporters in N. crassa. Eukaryotic. Cell 2009, 8, 1845–1855. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Nimrichter, L.; Oliveira, D.L.; Frases, S.; Miranda, K.; Zaragoza, O.; Alvarez, M.; Nakouzi, A.; Feldmesser, M.; Casadevall, A. Vesicular polysaccharide export in Cryptococcus neoformans is a eukaryotic solution to the problem of fungal trans-cell wall transport. Eukaryot. Cell 2007, 6, 48–59. [Google Scholar] [CrossRef]
- Fox, D.S.; Cruz, M.C.; Sia, R.A.; Ke, H.; Cox, G.M.; Cardenas, M.E.; Heltman, J. Calcineurin regulatory subunit is essential for virulence and mediates interactions with FKB12-FK506 in Cryptococcus neoformans. Mol. Microbiol. 2001, 39, 835–849. [Google Scholar] [CrossRef]
- Kozubowski, L.; Lee, S.C.; Heitman, J. Signalling pathways in the pathogenesis of Cryptococcus. Cell Microbiol. 2009, 11, 370–380. [Google Scholar] [CrossRef]
- Cole, L.; Orlovich, D.A.; Ashford, A.E. Structure, function and motility of vacuoles in filamentous fungi. Fungal Genet. Biol. 1998, 24, 86–100. [Google Scholar] [CrossRef]
- Fisher-Parton, S.; Parton, R.M.; Hickey, P.C.; Dijksterhuis, J.; Atkinson, H.A.; Read, N.D. Confocal microscopy of FM4-64 as a tool for anayzing endocytosis and vesicle trafficking in living fungal hyphae. J. Microscop. 2000, 198, 246–259. [Google Scholar] [CrossRef]
- Hickey, P.C.; Swift, S.R.; Roca, M.G. Live-cell imaging of filamentous fungi using vital fluorescent dyes and confocal microscopy. Methods Microbiol. 2004, 34, 63–87. [Google Scholar]
- Kim, H.S.; Czymmek, K.J.; Patel, A.; Modla, S.; Nohe, A.; Duncan, R.; Gilroy, S.; Kang, S. Expression of the Cameleon calcium biosensor in fungi reveals distinct Ca(2+) signatures associated with polarized growth, development, and pathogenesis. Fungal. Genet. Biol. 2012, 49, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.P.; Zhou, X.L.; Lin, J.; Loukin, S.H.; Kung, C.; Saimi, Y. A TRP homolog in Saccharomyces cerevisiae forms an intracellular Ca(2+)-permeable channel in the yeast vacuolal membrane. Proc. Natl. Acad. Sci. USA 2001, 98, 7801–7805. [Google Scholar] [CrossRef]
- Cunningham, K.W. Calcium Signaling Networks in Yeasts, in Calcium Signaling, 2nd, ed.; Putney, J.W., Ed.; Taylor & Francis Group, CRC Press: Florida, FL, USA, 2005; pp. 107–201. [Google Scholar]
- Denis, V.; Cyert, M.S. Internal Ca2+ release in yeast is triggered by hypertonic shock and mediated by a TRP channel homologue. J. Cell. Biol. 2002, 156, 29–34. [Google Scholar] [CrossRef]
- Zhou, X.-L.; Batiza, A.F.; Loukin, S.H.; Palmer, C.P.; Kung, C.; Saimi, Y. The transient receptor potential channel on the yeast vacuole is mechanosensitive. Proc. Natl. Acad. Sci. USA 2003, 100, 7105–7110. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Ohsumi, Y.; Tanifuji, M.; Kasai, M.; Anraku, Y. Vacuolal ion channel of the yeast, Saccharomyces cerevisiae. J. Biol. Chem. 1987, 262, 17260–17263. [Google Scholar] [CrossRef]
- Bertl, A.; Slayman, C.L. Cation-selective channels in the vacuolal membrane of Saccharomyces: Dependence on calcium, redox state, and voltage. Proc. Natl. Acad. Sci. USA 1990, 87, 7824–7828. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.P.; Shen, D.; Wang, X.; Dawson, T.; Li, X.; Zhang, Q.; Cheng, X.; Zhang, Y.; Weisman, L.S.; Delling, M.; et al. PI(3,5)P(2) controls membrane trafficking by direct activation of mucolipin Ca(2+) release channels in the endolysosome. Nat. Commun. 2010, 1, 38. [Google Scholar] [CrossRef]
- Popa, C.V.; Dumitru, I.; Ruta, L.L.; Danet, A.F.; Farcasanu, I.C. Exogenous oxidative stress induces Ca2+ release in the yeast Saccharomyces cerevisiae. FEBS J. 2010, 277, 4027–4038. [Google Scholar] [CrossRef]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.Y.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, L.; Rottensteiner, H.; Girzalsky, W.; Scarcia, P.; Palmieri, F.; Erdmann, R. Identification and functional reconstitution of the yeast peroxisomal adenine nucleotide transporter. EMBO J. 2001, 20, 5049–5059. [Google Scholar] [CrossRef] [PubMed]
- Visser, W.F.; van Roermund, C.W.; Ijlst, L.; Waterham, H.R.; Wanders, R.J. Metabolite transport across the peroxisomal membrane. Biochem. J. 2007, 401, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Titorenko, V.I.; Rachubinski, R.A. The peroxisome: Orchestrating important developmental decisions from inside the cell. J. Cell Biol. 2004, 164, 641–645. [Google Scholar] [CrossRef]
- Martin, J.F.; Liras, P. Transfer of Secondary Metabolite Gene Clusters: Assembly and Reorganization of the β-Lactam Gene Cluster from Bacteria to Fungi and Arthropods In Horizontal Gene Transfer: Breaking Borders between Living Kingdoms; Villa, T.G., Viñas, M., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 337–361. [Google Scholar]
- Hagestad, O.C.; Hou, L.; Andersen, J.H.; Hansen, E.H.; Altermark, B.; Li, C.; Kuhnert, E.; Cox, R.J.; Crous, P.W.; Spatafora, J.W.; et al. Genomic characterization of three marine fungi, including Emericellopsis atlantica sp. nov. with signatures of a generalist lifestyle and marine biomass degradation. IMA Fungus 2021, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-H.; Choi, Y.-J.; Kwon, B.; Choo, Y.-M.; Yu, K.-Y.; Kim, J. Regulation of secondary metabolism by calmodulin signaling in filamentous fungi. Rev. Iber. Micol. 2018, 36, 167–168. [Google Scholar] [CrossRef] [PubMed]
- Bouché, N.; Yellin, A.; Snedden, W.A.; Fromm, H. Plant specific calmodulin-binding proteins. Annu. Rev. Plant Biol. 2005, 56, 435–466. [Google Scholar] [CrossRef] [PubMed]
- Espeso, E.A. The CRaZy Calcium Cycle. Adv. Exp. Med. Biol. 2016, 892, 169–186. [Google Scholar] [PubMed]
- Chigri, F.; Soll, J.; Vothknecht, U.C. Calcium regulation of chloroplasts protein import. Plant. J. 2005, 42, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Chigri, F.; Hormann, F.; Stamp, A.; Stammers, D.K.; Bolter, B.; Soll, J.; Vothknecht, U.C. Calcium regulation of chloroplasts protein translocation is mediated by calmodulin binding to Tic32. Proc. Natl. Acad. Sci. USA 2006, 103, 16051–16056. [Google Scholar] [CrossRef]
- Chigri, F.; Flosdorff, S.; Pilz, S.; Kölle, E.; Dolze, E.; Gietl, C.; Vothknecht, U.C. The Arabidopsis calmodulin-like proteins AtCML30 and AtCML3 are targeted to mitochondria and peroxisomes, respectively. Plant. Mol. Biol. 2012, 78, 211–222. [Google Scholar] [CrossRef]
- Kuhn, S.; Bussemer, J.; Chigri, F.; Vothknecht, U.C. Calcium depletion and calmodulin inhibition affect the import of nuclear-encoded protein into plant mitochondria. Plant J. 2009, 58, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.H.; Essers, J.; Humbel, B.M.; Verkleij, A.J. Enrichment of Penicillium chrysogenum microbodies by isopycnic centrifugation in nycodenz as visualized with immuno-electron microscopy. Biochim. Biophys. Acta 1995, 1245, 215–220. [Google Scholar] [CrossRef]
- Campos, C.; Lázaro-Rodríguez, T.G.; Fragoso-Soriano, R.; Fernández, F.J. Vesicular transport and secretion of penicillin G in Penicillium rubens P2-32-T. Arch. Microbiol. 2020, 202, 1257–1262. [Google Scholar] [CrossRef] [PubMed]
- Chanda, A.; Roze, L.V.; Kang, S.; Artymovich, K.A.; Hicks, G.R.; Raikhel, N.V.; Calco, A.M.; Linz, J.E. A key role for vesicles in fungal secondary metabolism. Proc. Natl. Acad. Sci. USA 2009, 106, 19533–19538. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Microorganism | Influx Transporters Vcx1 Class | Efflux Transporter TRPs Yvc1 Class | Efflux Transporter TRPs CSC Class |
|---|---|---|---|
| Saccharomyces cerevisiae | NP_010155, 411 aa, 100%. Vcx1 [60] | QHB11688, 675 aa, 100%. Yvc1 [62] | Q06538, 782 aa, 100 %. CSC. [63] |
| Candida albicans | XP_711893, 416 aa, 62.5% | KHC88137, 675 aa, 44.8% | KGQ98295, 866 aa, 46.8% [44] |
| Gibberella zeae | XP_011319968, 456 aa, 48.9% | TRPGz, XP_011321451. 692 aa, 36.3% [64] | EYB29755, 866 aa, 37.7% XP_011328006, 884 aa, 36.5% |
| Penicillium chrysogenum | KZN93851, 449 aa, 52.5%. VcxA. KZN88098, 440 aa, 45.2%. VcxB. KZN85469, 552 aa, 44.9%. VcxC. KZN88096, 270 aa, 35.3%. VcxD. KZN92764, 461 aa, 34.3%. VcxE | KZN84307, 628 aa, 35.3% | KZN90483, 832 aa, 41.8% [15] |
| Cryptococcus neoformans | OWZ58227, 403 aa, 45,3% OWZ62586, 606 aa, 45,7% [65] | OWZ59838, 622 aa, 30.6% | OWZ78565, 1080 aa, 29.7% |
| Neurospora crassa | XP_011394995, 507, 50%. CAX. [4] AAC08353, 443 aa, 49.5% | KHE84196, 685 aa 37.5% | XP_964945, 930 aa, 37.6% |
| Aspergillus fumigatus | X_750174, 462 aa, 53.55% | XP_001481630, 670 aa, 35.1% | XP_749963, 835 aa, 42.2% |
| Acremonium chrysogenum | KFH45419, 448 aa, 49.75% | KFH43541, 671 aa, 39.7% | KFH48720, 866 aa, 37.6% |
| Fungi | Accession Number | Amino Acids | Identity to CefP (%) |
|---|---|---|---|
| A. chrysogenum | KFH48720, CefP | 866 | 100.0 |
| Emericellopsis atlantica | XP_046122791, CefP | 872 | 70.5 |
| Tolypocladium paradoxum | POR38094 | 875 | 68.7 |
| Claviceps lovelessii | KAH0526729 | 868 | 65.2 |
| Metarhizium anisopliae | KFG81683 | 867 | 65.0 |
| Fusarium oxysporum | KAG7415113 | 858 | 64.2 |
| Pochonia chlamydosporia | XP_018145584 | 863 | 63.7 |
| Trichoderma harzianum | KKP01098 | 873 | 63.2 |
| Trichoderma virens | XP_013952278 | 879 | 62.6 |
| G. zeae | EYB29755 | 866 | 61.6 |
| N. crassa | XP_964945 | 930 | 49.9 |
| As. fumigatus | XP_746993 | 835 | 43.5 |
| P. chrysogenum | KZN90483 | 832 | 42.2 |
| S. cerevisiae | Q06538 CSC1p | 782 | 36.9 |
| C. albicans | KGQ92391 | 866 | 34.6 |
| C. neoformans | OWZ67604 | 1080 | 27.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín, J.F. Vacuolal and Peroxisomal Calcium Ion Transporters in Yeasts and Fungi: Key Role in the Translocation of Intermediates in the Biosynthesis of Fungal Metabolites. Genes 2022, 13, 1450. https://doi.org/10.3390/genes13081450
Martín JF. Vacuolal and Peroxisomal Calcium Ion Transporters in Yeasts and Fungi: Key Role in the Translocation of Intermediates in the Biosynthesis of Fungal Metabolites. Genes. 2022; 13(8):1450. https://doi.org/10.3390/genes13081450
Chicago/Turabian StyleMartín, Juan F. 2022. "Vacuolal and Peroxisomal Calcium Ion Transporters in Yeasts and Fungi: Key Role in the Translocation of Intermediates in the Biosynthesis of Fungal Metabolites" Genes 13, no. 8: 1450. https://doi.org/10.3390/genes13081450