
A DNA-Damage Inducible Gene Promotes the Formation of Antibiotic Persisters in Response to the Quorum Sensing Signaling Peptide in Streptococcus mutans


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culturing and Genetic Manipulations
2.2. Preparation of Inoculum for Persister Assays
2.3. Persister Assays
2.4. Antimicrobial Activity Analysis
2.5. Isolation of Persisters-CSP for Gene Expression Analysis
2.6. DNA Damage for Gene Expression Analysis
2.7. RNA Isolation and Gene Expression Analysis
2.8. Statistical Analysis
3. Results
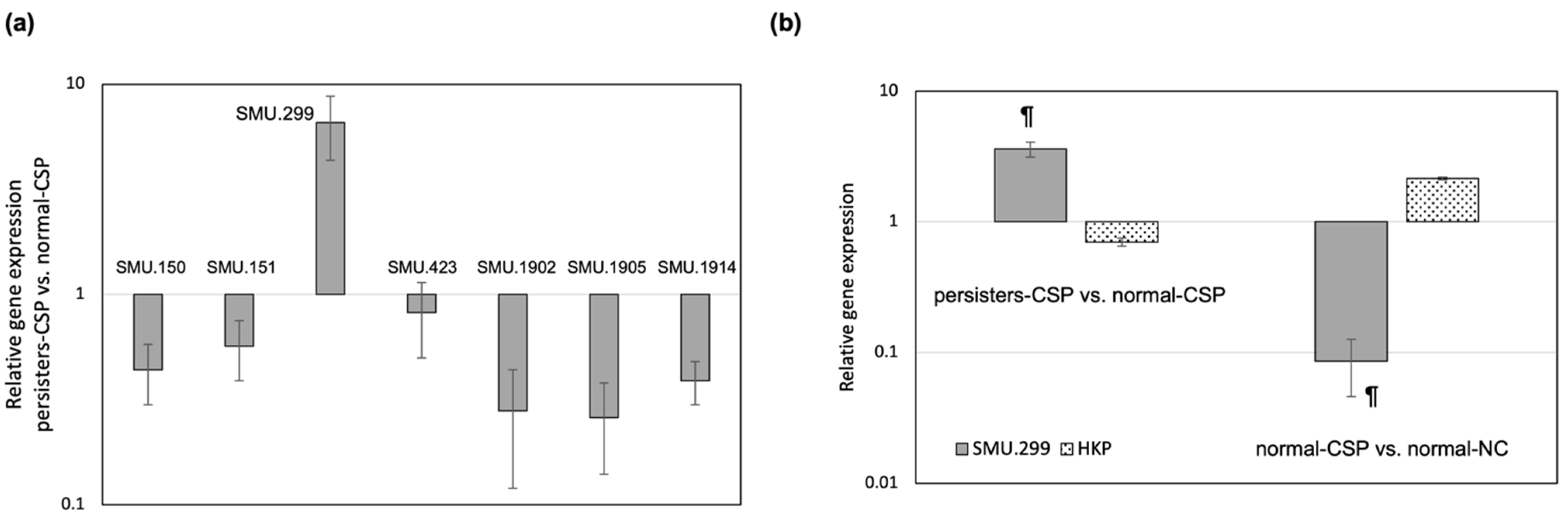
3.1. A Bacteriocin Gene Is Specifically Activated in the Antibiotic Persister Population
3.2. SMU.299 Is a DNA-Damage Inducible Gene
3.3. Pep299 Does Not Possess Bacteriocin Activity
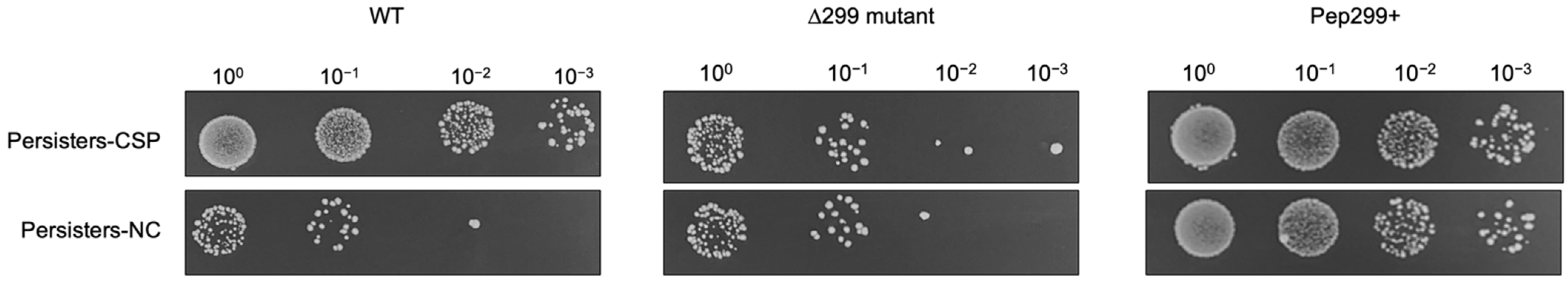
3.4. The CSP-Inducible Persistence Phenotype Is Abolished in a Mutant Deficient in Pep299
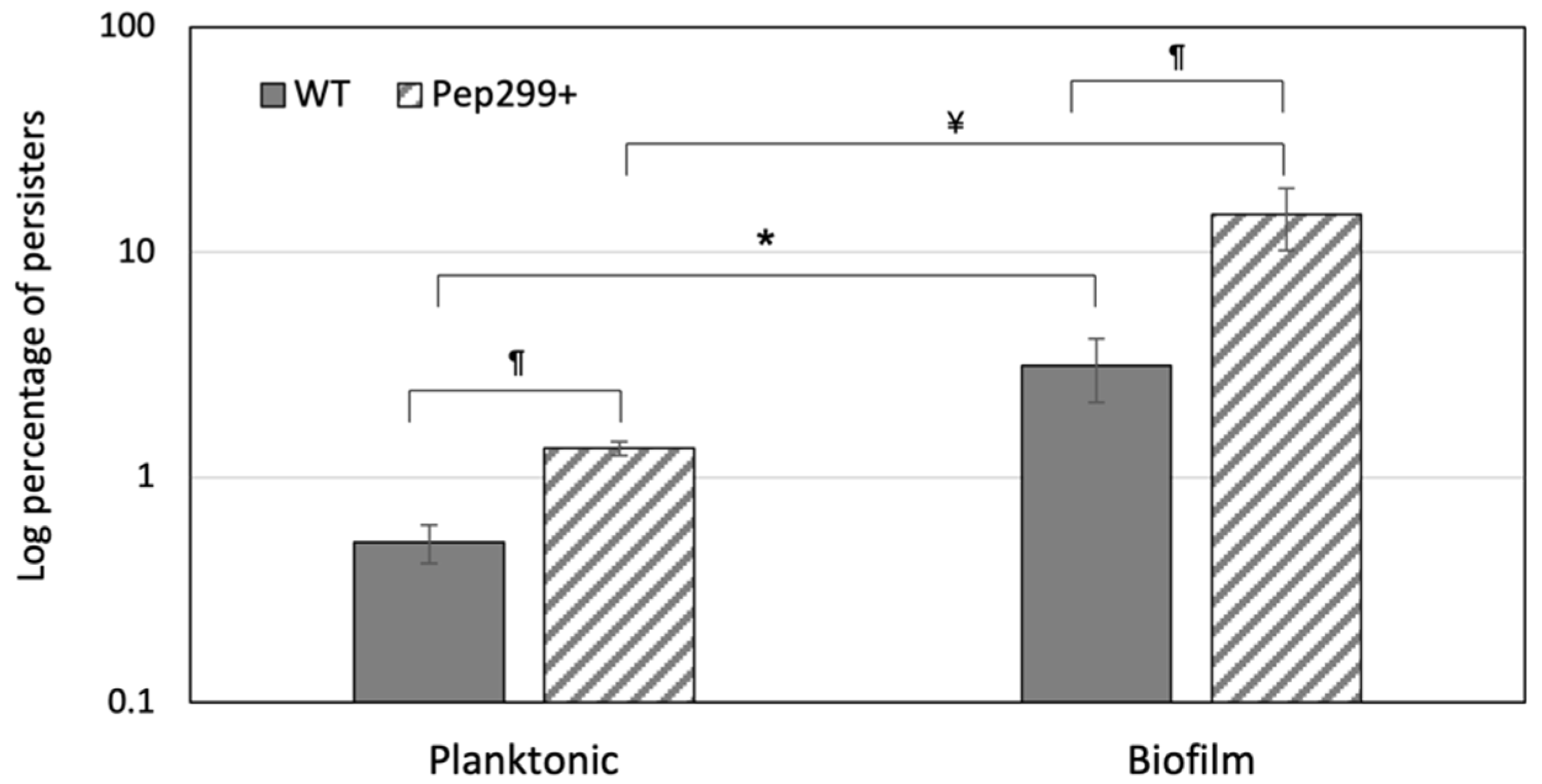
3.5. Pep299 Enhances the Formation of Persisters in Biofilms
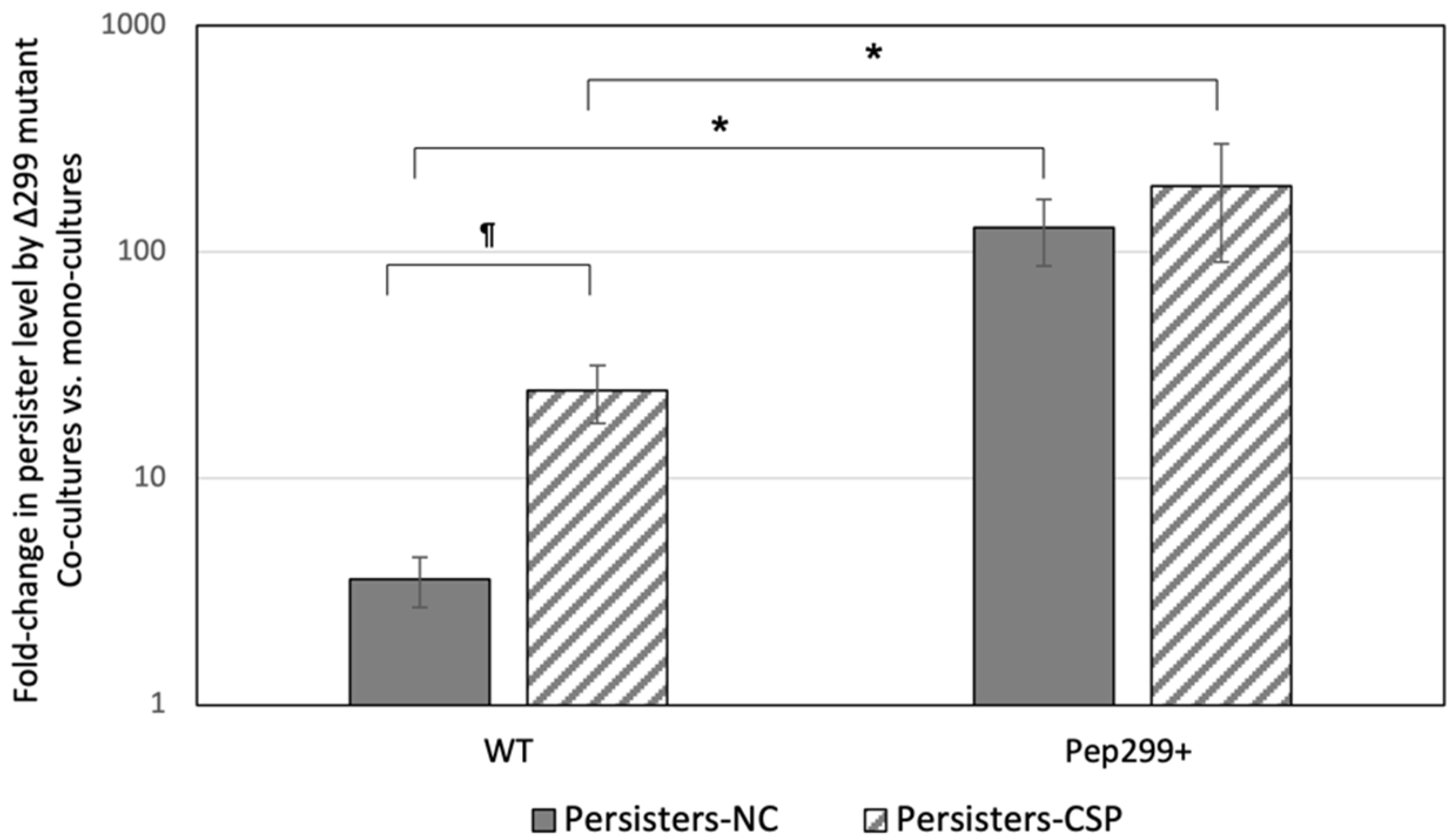
3.6. Secreted Pep299 Promotes the Formation of Antibiotic Persisters in Neighboring Cells
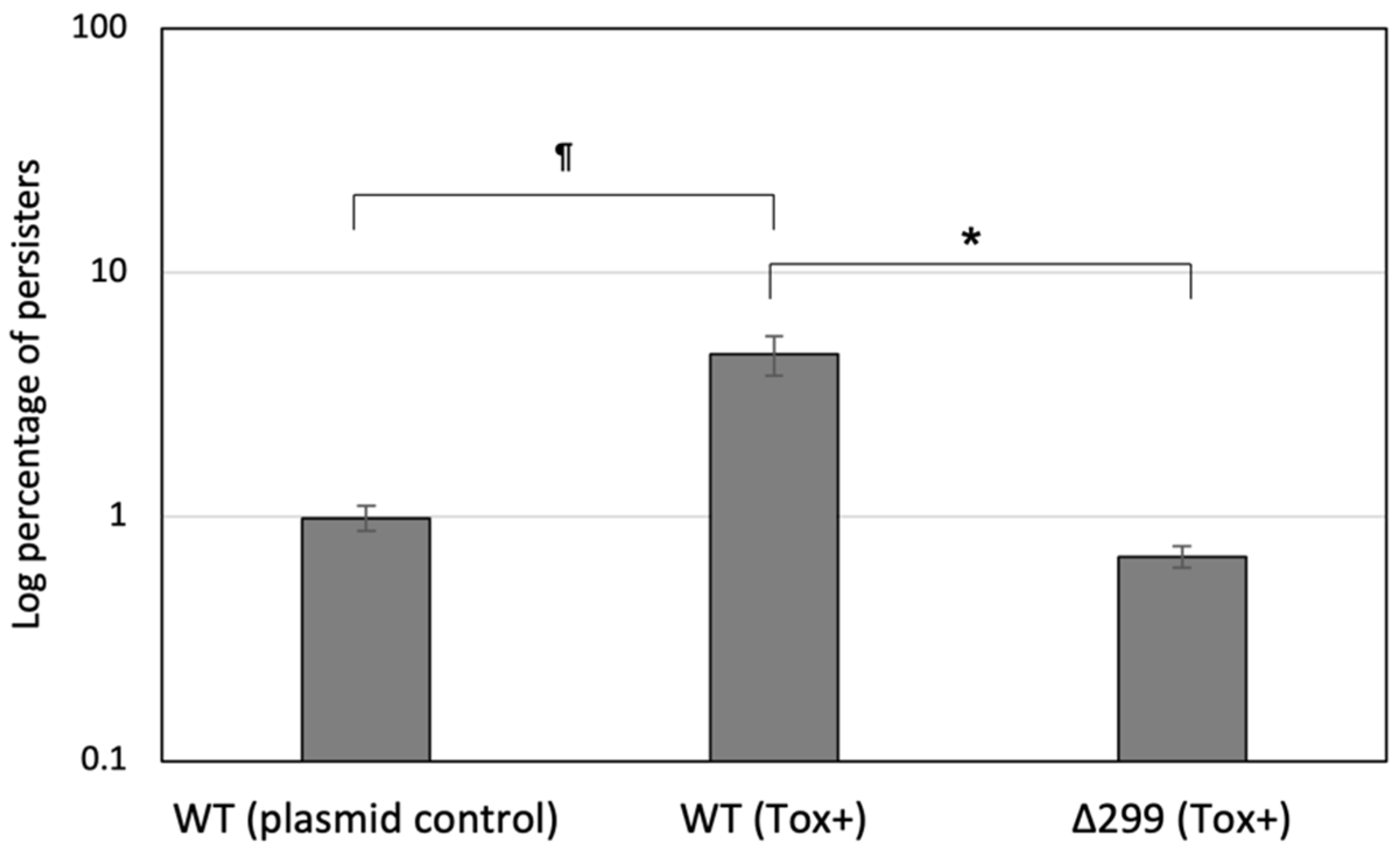
3.7. Pep299 Induces Persister Formation via the Activation of a Specific Toxin-Antitoxin System
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef]
- Li, Y.H.; Lau, P.C.; Lee, J.H.; Ellen, R.P.; Cvitkovitch, D.G. Natural genetic transformation of Streptococcus mutans growing in biofilms. J. Bacteriol. 2001, 183, 897–908. [Google Scholar] [CrossRef]
- Monnet, V.; Juillard, V.; Gardan, R. Peptide conversations in Gram-positive bacteria. Crit. Rev. Microbiol. 2016, 42, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Håvarstein, L.S.; Coomaraswamy, G.; Morrison, D.A. An unmodified heptadecapeptide pheromone induces competence for genetic transformation in Streptococcus pneumoniae. Proc. Natl. Acad. Sci. USA 1995, 92, 11140–11144. [Google Scholar] [CrossRef] [PubMed]
- Kreth, J.; Merritt, J.; Zhu, L.; Shi, W.; Qi, F. Cell density- and ComE-dependent expression of a group of mutacin and mutacin-like genes in Streptococcus mutans. FEMS Microbiol. Lett. 2006, 265, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Shanker, E.; Federle, M.J. Quorum sensing regulation of competence and bacteriocins in Streptococcus pneumoniae and mutans. Genes 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Sieiro, P.; Montalbán-Lopez, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef]
- Wang, C.Y.; Dawid, S. Mobilization of bacteriocins during competence in Streptococci. Trends Microbiol. 2018, 26, 389–391. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Cook, L.C.; Federle, M.J. Peptide pheromone signaling in Streptococcus and Enterococcus. FEMS Microbiol. Rev. 2014, 38, 473–492. [Google Scholar] [CrossRef]
- Dufour, D.; Lévesque, C.M. Cell death of Streptococcus mutans induced by a quorum-sensing peptide occurs via a conserved streptococcal autolysin. J. Bacteriol. 2013, 195, 105–114. [Google Scholar] [CrossRef]
- Merritt, J.; Qi, F. The mutacins of Streptococcus mutans: Regulation and ecology. Mol. Oral Microbiol. 2012, 27, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Kawada-Matsuo, M.; Le, M.N.; Hisatsune, J.; Oogai, Y.; Nakano, Y.; Nakata, M.; Miyawaki, S.; Sugai, M.; Komatsuzawa, H. Comprehensive analysis of bacteriocins in Streptococcus mutans. Sci. Rep. 2021, 11, 12963. [Google Scholar] [CrossRef] [PubMed]
- Dirix, G.; Monsieurs, P.; Dombrecht, B.; Daniels, R.; Marchal, K.; Vanderleyden, J.; Michiels, J. Peptide signal molecules and bacteriocins in gram-negative bacteria: A genome-wide in silico screening for peptides containing a double-glycine leader sequence and their cognate transporters. Peptides 2004, 25, 1425–1440. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, K.; Alegria, A.; Bron, P.A.; de Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C.; et al. Stress physiology of lactic acid bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 837–890. [Google Scholar] [CrossRef] [PubMed]
- Claverys, J.P.; Prudhomme, M.; Martin, B. Induction of competence regulon as a general response to stress in gram-positive bacteria. Annu. Rev. Microbiol. 2006, 60, 451–475. [Google Scholar] [CrossRef]
- Perry, J.A.; Jones, M.B.; Peterson, S.N.; Cvitkovitch, D.G.; Lévesque, C.M. Peptide alarmone signaling triggers an auto-active bacteriocin necessary for genetic competence. Mol. Microbiol. 2009, 72, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Erill, I.; Campoy, S.; Barbé, J. Aeons of distress: An evolutionary perspective on the bacterial SOS response. FEMS Microbiol. Rev. 2007, 31, 637–656. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, X.; Polard, P.; Claverys, J.P. Induction of competence for genetic transformation by antibiotics: Convergent evolution of stress responses in distant bacterial species lacking SOS? Curr. Opin. Microbiol. 2012, 15, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Dufour, D.; Lévesque, C.M. Bacterial behaviors associated with the quorum-sensing peptide pheromone (‘alarmone’) in streptococci. Future Microbiol. 2013, 8, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Leung, V.; Lévesque, C.M. A stress-inducible quorum-sensing peptide mediates the formation of persister cells with noninherited multidrug tolerance. J. Bacteriol. 2012, 194, 2265–2274. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, B.; Fauvart, M.; Michiels, J. Formation, physiology, ecology, evolution and clinical importance of bacterial persisters. FEMS Microbiol. Rev. 2017, 41, 219–251. [Google Scholar] [CrossRef]
- Ronneau, S.; Hill, P.W.; Helaine, S. Antibiotic persistence and tolerance: Not just one and the same. Curr. Opin. Microbiol. 2021, 64, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.K.; Knabel, S.J.; Kwan, B.W. Bacterial persister cell formation and dormancy. Appl. Environ. Microbiol. 2013, 79, 7119–7121. [Google Scholar] [CrossRef] [PubMed]
- Moldoveanu, A.L.; Rycroft, J.A.; Helaine, S. Impact of bacterial persisters on their host. Curr. Opin. Microbiol. 2021, 59, 65–71. [Google Scholar] [CrossRef]
- Zou, J.; Peng, B.; Qu, J.; Zheng, J. Are bacterial persisters dormant cells only? Front. Microbiol. 2022, 12, 708580. [Google Scholar] [CrossRef]
- Verstraete, L.; Van den Bergh, B.; Verstraeten, N.; Michiels, J. Ecology and evolution of antibiotic persistence. Trends Microbiol. 2022, 30, 466–479. [Google Scholar] [CrossRef]
- Leung, V.; Ajdic, D.; Koyanagi, S.; Lévesque, C.M. The formation of Streptococcus mutans persisters induced by the quorum-sensing peptide pheromone is affected by the LexA regulator. J. Bacteriol. 2015, 197, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, C.M.; Mair, R.W.; Perry, J.A.; Lau, P.C.Y.; Li, Y.H.; Cvitkovitch, D.G. Systemic inactivation and phenotypic characterization of two-component systems in expression of Streptococcus mutans virulence properties. Lett. Appl. Microbiol. 2007, 45, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Biswas, I.; Jha, J.K.; Fromm, N. Shuttle expression plasmids for genetic studies in Streptococcus mutans. Microbiology 2008, 154, 2275–2282. [Google Scholar] [CrossRef] [PubMed]
- Dufour, D.; Villemin, C.; Perry, J.A.; Lévesque, C.M. Escape from the competence state in Streptococcus mutans is governed by the bacterial population density. Mol. Oral Microbiol. 2016, 31, 501–514. [Google Scholar] [CrossRef]
- Kaldalu, N.; Hauryliuk, V.; Tenson, T. Persisters-as elusive as ever. Appl. Microbiol. Biotechnol. 2016, 100, 6545–6553. [Google Scholar] [CrossRef]
- Dufour, D.; Barbour, A.; Chan, Y.; Cheng, M.; Rahman, T.; Thorburn, M.; Stewart, C.; Finer, Y.; Gong, S.G.; Lévesque, C.M. Genetic analysis of mutacin B-Ny266, a lantibiotic active against caries pathogens. J. Bacteriol. 2020, 202, e00762-19. [Google Scholar] [CrossRef] [PubMed]
- Tagg, J.R.; Bannister, L.V. “Fingerprinting” β-haemolytic streptococci by their production of and sensitivity to bacteriocin-like inhibitors. J. Med Microbiol. 1979, 12, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Rukke, H.V.; Høvik, H.; Åmdal, H.A.; Chen, T.; Morrison, D.A.; Peterson, F.C. Comprehensive transcriptome profiles of Streptococcus mutans UA159 map core streptococcal competence genes. mSystems 2016, 1, e00038-15. [Google Scholar] [CrossRef] [PubMed]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Tymoszewska, A.; Diep, D.B.; Wirtek, P.; Aleksandrzak-Piekarczyk, T. The non-lantibiotic bacteriocin garvicin Q targets Man-PTS in a broad spectrum of sensitive bacterial genera. Sci. Rep. 2017, 7, 8359. [Google Scholar] [CrossRef] [PubMed]
- Hale, J.D.; Ting, Y.T.; Jack, R.W.; Tagg, J.R.; Heng, N.C. Bacteriocin (mutacin) production by Streptococcus mutans genome sequence reference strain UA159: Elucidation of the antimicrobial repertoire by genetic dissection. Appl. Environ. Microbiol. 2005, 71, 7613–7617. [Google Scholar] [CrossRef] [PubMed]
- Gollan, B.; Grabe, G.; Michaux, C.; Helaine, S. Bacterial persisters and infection: Past, present, and progressing. Annu. Rev. Microbiol. 2019, 73, 359–385. [Google Scholar] [CrossRef] [PubMed]
- Koyanagi, S.; Lévesque, C.M. Characterization of a Streptococcus mutans intergenic region containing a small toxic peptide and its cis-encoded antisense small RNA antitoxin. PLoS ONE 2013, 8, e54291. [Google Scholar] [CrossRef] [PubMed]
- Dufour, D.; Mankovskaia, A.; Chan, Y.; Motavaze, K.; Gong, S.G.; Lévesque, C.M. A tripartite toxin-antitoxin module induced by quorum-sensing is associated with the persistence phenotype in Streptococcus mutans. Mol. Oral Microbiol. 2018, 33, 420–429. [Google Scholar] [CrossRef]
- Xie, Y.; Wei, Y.; Shen, Y.; Li, X.; Zhou, H.; Tai, C.; Deng, Z.; Ou, H.Y. TADB 20: An updated database of bacterial type II toxin-antitoxin loci. Nucleic Acids Res. 2018, 46, D749–D753. [Google Scholar] [CrossRef]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, targets, and triggers: An overview of toxin-antitoxin biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.; Sahu, B.R.; Suar, M. Plausible role of bacterial toxin-antitoxin system in persister cell formation and elimination. Mol. Oral Microbiol. 2019, 34, 97–107. [Google Scholar] [CrossRef]
- Meng, P.; Lu, C.; Zhang, Q.; Lin, J.; Chen, F. Exploring the genomic diversity and cariogenic differences of Streptococcus mutans strains through pan-genome and comparative genome analysis. Curr. Microbiol. 2017, 74, 1200–1209. [Google Scholar] [CrossRef]
- Jack, R.W.; Tagg, J.R.; Ray, B. Bacteriocins of gram-positive bacteria. Microbiol. Rev. 1995, 59, 171–200. [Google Scholar] [CrossRef] [PubMed]
- Nes, I.F.; Diep, D.B.; Holo, H. Bacteriocin diversity in Streptococcus and Enterococcus. J. Bacteriol. 2007, 189, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Hallez, R.; Geeraerts, D.; Sterckx, Y.; Mine, N.; Loris, R.; Van Melderen, L. New toxins homologous to pare belonging to three-component toxin-antitoxin systems in Escherichia coli O157:H7. Mol. Microbiol. 2010, 76, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Mignolet, J.; Fontaine, L.; Sass, A.; Nannan, C.; Mahillon, J.; Coenye, T.; Hols, P. Circuitry rewiring directly couples competence to predation in the gut dweller Streptococcus salivarius. Cell Rep. 2018, 22, 1627–1638. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Resistance a | Description | Reference |
|---|---|---|---|---|
| UA159 | wild-type | ATCC | ||
| Δ299 | ΔSMU.299 | Ery or Spc | In-frame insertion-deletion | This study |
| Pep299+ | ΔSMU.299(pS921) | Ery Cml | Ectopic expression of pep299 | This study |
| WT(plasmid control) | UA159(pIB166) | Cml | Empty plasmid | This study |
| WT(Tox+) | UA159(pS1109) | Cml | Ectopic expression of tox40 | This study |
| Δ299(plasmid control) | ΔSMU.299(pIB166) | Spc Cml | Empty plasmid | This study |
| Δ299(Tox+) | ΔSMU.299(pS1109) | Spc Cml | Ectopic expression of tox40 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dufour, D.; Zhao, H.; Gong, S.-G.; Lévesque, C.M. A DNA-Damage Inducible Gene Promotes the Formation of Antibiotic Persisters in Response to the Quorum Sensing Signaling Peptide in Streptococcus mutans. Genes 2022, 13, 1434. https://doi.org/10.3390/genes13081434
Dufour D, Zhao H, Gong S-G, Lévesque CM. A DNA-Damage Inducible Gene Promotes the Formation of Antibiotic Persisters in Response to the Quorum Sensing Signaling Peptide in Streptococcus mutans. Genes. 2022; 13(8):1434. https://doi.org/10.3390/genes13081434
Chicago/Turabian StyleDufour, Delphine, Haowei Zhao, Siew-Ging Gong, and Céline M. Lévesque. 2022. "A DNA-Damage Inducible Gene Promotes the Formation of Antibiotic Persisters in Response to the Quorum Sensing Signaling Peptide in Streptococcus mutans" Genes 13, no. 8: 1434. https://doi.org/10.3390/genes13081434