
Complete Mitochondrial Genome of Two Ectoparasitic Capsalids (Platyhelminthes: Monogenea: Monopisthocotylea): Gene Content, Composition, and Rearrangement

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. Mitochondrial Genome Sequencing and Assembling
2.3. Sequence Annotation and Analysis
2.4. Gene Rearrangement Analysis and Phylogenomic Reconstruction
3. Results and Discussion
3.1. Genome Organization
3.2. Protein Coding Genes and Codon Usage
3.3. Transfer RNA Genes and Ribosomal RNA
3.4. Gene Rearrangement
3.5. Phylogenetic Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rohde, K. (Ed.) Marine Parasitology; Csiro Publishing: Melbourne, Australia, 2005. [Google Scholar]
- Bauer, O.N. Spread of parasites and diseases of aquatic organisms by acclimatization: A short review. J. Fish Biol. 1991, 39, 679–686. [Google Scholar] [CrossRef]
- Hutson, K.S.; Ernst, I.; Whittington, I.D. Risk assessment for metazoan parasites of yellowtail kingfish Seriola lalandi (Perciformes: Carangidae) in South Australian sea-cage aquaculture. Aquaculture 2007, 271, 85–99. [Google Scholar] [CrossRef]
- Whittington, I.D. The Capsalidae (Monogenea: Monopisthocotylea): A review of diversity, classification and phylogeny with a note about species complexes. Folia Parasit. 2004, 51, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittington, I.D.; Deveney, M.R.; Morgan, J.A.T.; Chisholm, L.A.; Adlard, R.D. A preliminary phylogenetic analysis of the Capsalidae (Platyhelminthes: Monogenea: Monopisthocotylea) inferred from large subunit rDNA sequences. Parasitology 2004, 128, 511–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chisholm, L.A.; Whittington, I.D. Review of the Capsalinae (Monogenea: Capsalidae). Zootaxa 2007, 1559, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Boeger, W.A.; Kritsky, D.C. Phylogeny and a revised classification of the Monogenoidea Bychowsky, 1937 (Platyhelminthes). Syst. Parasitol. 1993, 26, 1–32. [Google Scholar] [CrossRef]
- Mollaret, I.; Jamieson, B.G.; Justine, J.L. Phylogeny of the Monopisthocotylea and Polyopisthocotylea (Platyhelminthes) inferred from 28S rDNA sequences. Inter. J. Parasitol. 2000, 30, 171–185. [Google Scholar] [CrossRef]
- Olson, P.D.; Littlewood, D.T.J. Phylogenetics of the Monogenea–evidence from a medley of molecules. Inter. J. Parasitol. 2002, 32, 233–244. [Google Scholar] [CrossRef]
- Zhang, D.; Li, W.X.; Zou, H.; Wu, S.G.; Li, M.; Jakovlić, I.; Zhang, J.; Chen, R.; Wang, G.T. Mitochondrial genomes of two diplectanids (Platyhelminthes: Monogenea) expose paraphyly of the order Dactylogyridea and extensive tRNA gene rearrangements. Parasite. Vector. 2018, 11, 601. [Google Scholar] [CrossRef]
- Kearn, G.C. The egg, oncomiracidium and larval development of Entobdella soleae, a monogenean skin parasite of the common sole. Parasitology 1963, 53, 435–447. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, Z.H.; Yuan, K.; Ding, X.J. Three new recorded species in Capsalidae (monogenea) from fishes in the South China Sea. Acta Parasitol. Medica Entomol. Sin. 2019, 26, 251–255. [Google Scholar]
- Cameron, S. How to sequence and annotate insect mitochondrial genomes for systematic and comparative genomics research. Syst. Entomol. 2014, 39, 400–411. [Google Scholar] [CrossRef] [Green Version]
- Park, J.K.; Kim, K.H.; Kang, S.; Kim, W.; Eom, K.S.; Littlewood, D.T.J. A common origin of complex life cycles in parasitic flatworms: Evidence from the complete mitochondrial genome of Microcotyle sebastis (Monogenea: Platyhelminthes). BMC Evol. Biol. 2007, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, R.; Barker, S.C. Mitochondrial genomes of parasitic arthropods: Implications for studies of population genetics and evolution. Parasitology 2007, 134, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Huyse, T.; Buchmann, K.; Littlewood, D.T.J. The mitochondrial genome of Gyrodactylus derjavinoides (Platyhelminthes: Monogenea)—a mitogenomic approach for Gyrodactylus species and strain identification. Gene 2008, 417, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liang, D.; Mao, R.L.; Hillis, D.M.; Wake, D.B.; Cannatella, D.C. Efficient sequencing of anuran mtDNAs and a mitogenomic exploration of the phylogeny and evolution of frogs. Mol. Biol. Evol. 2013, 30, 1899–1915. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.P.; Shan, B.B.; Liu, Y.; Zhao, Y.; Sun, D.R. Next-generation sequencing yields the complete mitochondrial genome of the Capsala pricei Hidalgo, 1959 (Platyhelminthes: Monogenea) from South China Sea. Mitochondrial DNA Part B 2020, 5, 1964–1966. [Google Scholar] [CrossRef]
- Baeza, J.A.; Sepúlveda, F.A.; González, M.T. The complete mitochondrial genome and description of a new cryptic species of Benedenia Diesing, 1858 (Monogenea: Capsalidae), a major pathogen infecting the yellowtail kingfish Seriola lalandi Valenciennes in the South-East Pacific. Parasite Vector 2019, 12, 490. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, X.Y.; Li, Y.W.; Zhao, M.W.; Xie, M.Q.; Li, A.X. The complete mitochondrial genome of Neobenedenia melleni (Platyhelminthes: Monogenea): Mitochondrial gene content, arrangement and composition compared with two Benedenia species. Mol. Biol. Rep. 2014, 41, 6583–6589. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.P.; Liu, Y.; Shan, B.B.; Zhang, G.J.; Zhao, Y.; Sun, D.R.; Yu, W. The complete mitochondrial genome of the Capsaloides cristatus (Platyhelminthes, Monogenea), a pathogen of the sailfish (Istiophorus platypterus). Mitochondrial DNA Part B 2021, 6, 1053–1055. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Kuma, K.I.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Kim, J.; Lee, J.; Kim, S.; Min, G.S.; Park, J.K. The complete mitochondrial genome of an ectoparasitic monopisthocotylean fluke Benedenia hoshinai (Monogenea: Platyhelminthes). Mitochondrial DNA 2012, 23, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zou, H.; Jakovlić, I.; Wu, S.G.; Li, M.; Zhang, J.; Chen, R.; Li, W.X.; Wang, G.T. Mitochondrial genomes of two thaparocleidus species (Platyhelminthes: Monogenea) reveal the first rRNA gene rearrangement among the neodermata. Inter. J. Mol. Sci. 2019, 20, 4214. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Li, W.X.; Zou, H.; Wu, S.G.; Li, M.; Jakovlić, I.; Zhang, J.; Chen, R.; Wang, G.T. Mitochondrial genomes and 28S rDNA contradict the proposed obsoletion of the order Tetraonchidea (Platyhelminthes: Monogenea). Inter. J. Biol. Macromol. 2020, 143, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zou, H.; Wu, S.G.; Li, M.; Jakovlić, I.; Zhang, J.; Chen, R.; Wang, G.T.; Li, W.X. Sequencing of the complete mitochondrial genome of a fish-parasitic flatworm Paratetraonchoides inermis (Platyhelminthes: Monogenea): tRNA gene arrangement reshuffling and implications for phylogeny. Parasite Vector 2017, 10, 462. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Ruhlman, T.A.; Weng, M.L.; Hajrah, N.H.; Sabir, J.S.; Jansen, R.K. Contrasting patterns of nucleotide substitution rates provide insight into dynamic evolution of plastid and mitochondrial genomes of Geranium. Genome Boil. Evol. 2017, 9, 1766–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moritz, C.; Brown, W.M. Tandem duplications in animal mitochondrial DNAs: Variation in incidence and gene content among lizards. Proc. Natl. Acad. Sci. USA 1987, 84, 7183–7187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavrov, D.V.; Boore, J.L.; Brown, W.M. Complete mtDNA sequences of two millipedes suggest a new model for mitochondrial gene rearrangements: Duplication and nonrandom loss. Mol. Biol. Evol. 2002, 19, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rokas, A.; Ladoukakis, E.; Zouros, E. Animal mitochondrial DNA recombination revisited. Trends Ecol. Evol. 2003, 18, 411–417. [Google Scholar] [CrossRef]
- Bychowsky, B.E. Monogenetic Trematodes, Their Classification and Phylogeny; Izdatel’suo Akademiya Nauk SSSR: Moscow, Russia, 1957. [Google Scholar]
- Zhang, D.; Zou, H.; Wu, S.G.; Li, M.; Jakovlić, I.; Zhang, J.; Chen, R.; Wang, G.T.; Li, W.X. Sequencing, characterization and phylogenomics of the complete mitochondrial genome of Dactylogyrus lamellatus (Monogenea: Dactylogyridae). J. Helminthol. 2018, 92, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Justine, J.L.; Lambert, A.; Mattei, X. Spermatozoon ultrastructure and phylogenetic relationships in the monogeneans (Platyhelminthes). Inter. J. Parasitol. 1985, 15, 601–608. [Google Scholar] [CrossRef]
- Šimková, A.; Plaisance, L.; Matějusová, I.; Morand, S.; Verneau, O. Phylogenetic relationships of the Dactylogyridae Bychowsky, 1933 (Monogenea: Dactylogyridea): The need for the systematic revision of the Ancyrocephalinae Bychowsky, 1937. Syst. Parasitol. 2003, 54, 1–11. [Google Scholar] [CrossRef]





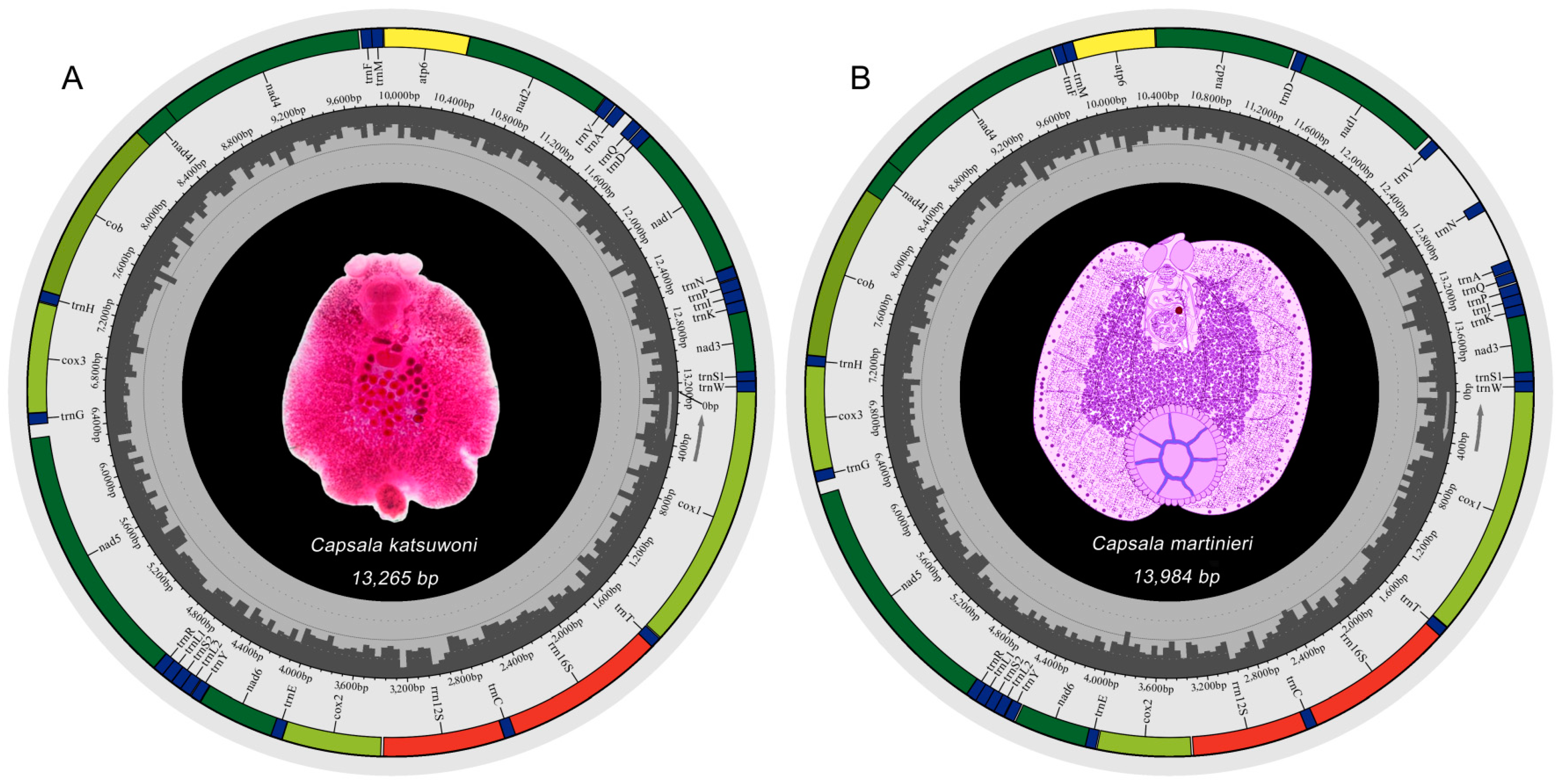{kind=link}
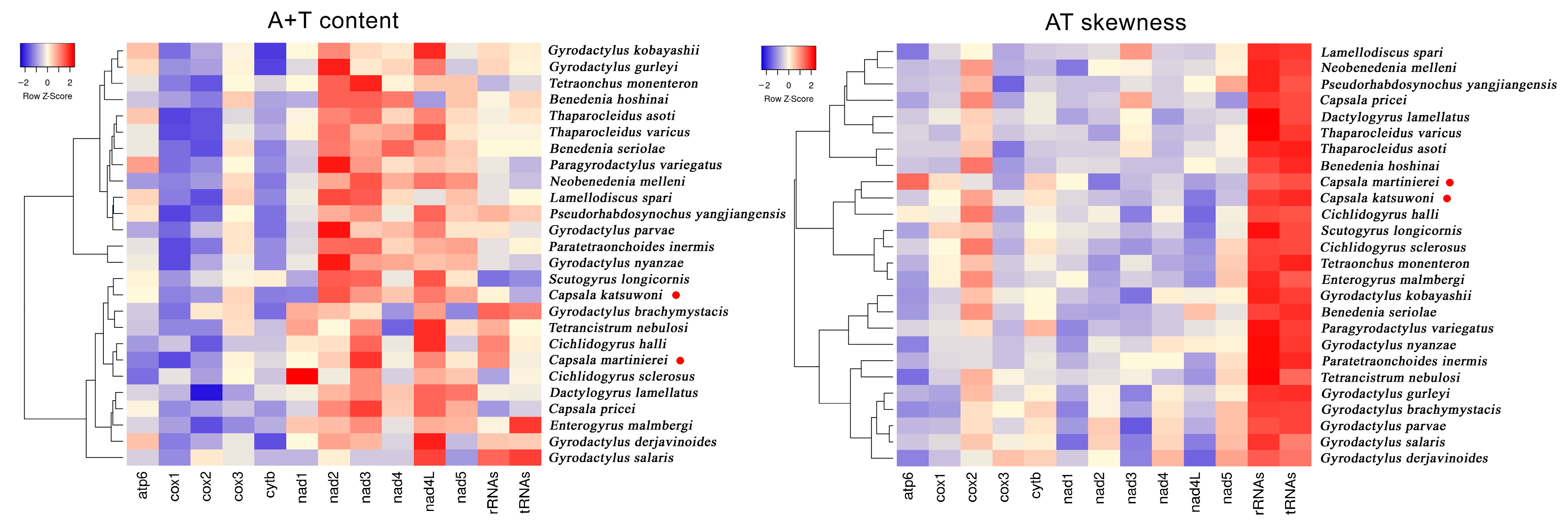{kind=link}
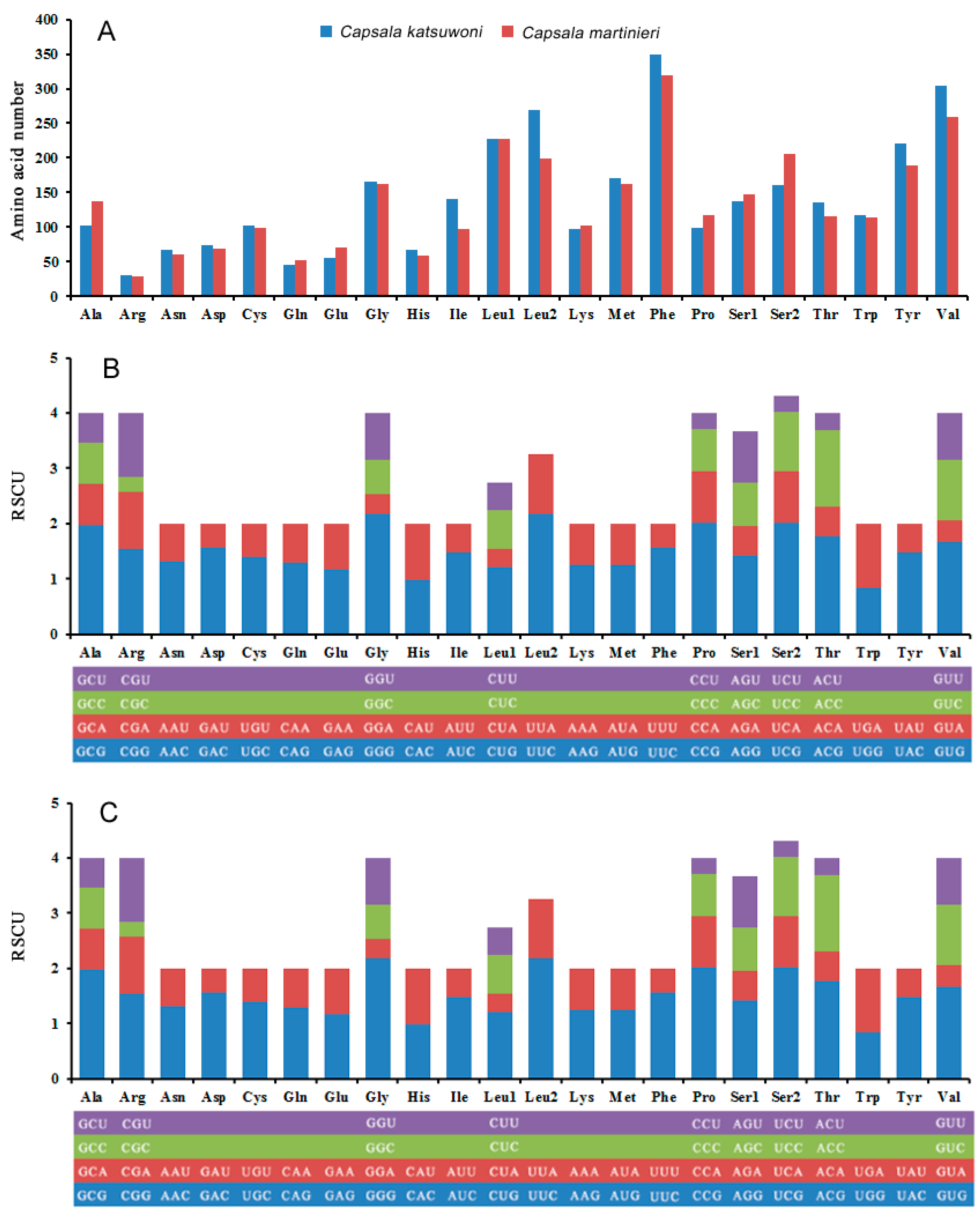{kind=link}
{kind=link}
| Order | Family | Species | Full Length | A% | T% | G% | C% | GenBank No. |
|---|---|---|---|---|---|---|---|---|
| Capsalidea | Capsalidae | Capsala katsuwoni | 13,265 | 25.4 | 41.1 | 18.9 | 14.6 | OL884727 |
| Capsalidea | Capsalidae | Capsala martinierei | 13,984 | 25.8 | 38.2 | 19.1 | 17.0 | OL790148 |
| Capsalidea | Capsalidae | Capsala pricei | 13,851 | 26.1 | 43.2 | 17.3 | 13.4 | MN746360 |
| Capsalidea | Capsalidae | Benedenia hoshinai | 13,554 | 28.7 | 45.4 | 14.9 | 11.0 | NC_014591 |
| Capsalidea | Capsalidae | Benedenia seriolae | 13,498 | 28.9 | 46.7 | 14.6 | 9.8 | AP019637 |
| Capsalidea | Capsalidae | Neobenedenia melleni | 13,270 | 30.7 | 45.2 | 14.7 | 9.4 | JQ038228 |
| Dactylogyridea | Ancyrocephalidae | Cichlidogyrus halli | 15,047 | 25.4 | 38.0 | 13.2 | 23.2 | MG970255 |
| Dactylogyridea | Ancyrocephalidae | Cichlidogyrus sclerosus | 15,052 | 24.9 | 40.5 | 11.7 | 22.9 | JQ038226 |
| Dactylogyridea | Dactylogyridae | Dactylogyrus lamellatus | 15,187 | 27.5 | 43.1 | 9.9 | 19.5 | KR871673 |
| Dactylogyridea | Ancyrocephalidae | Enterogyrus malmbergi | 14,107 | 27.0 | 42.0 | 10.2 | 20.8 | NC_048529 |
| Gyrodactylidea | Gyrodactylidae | Gyrodactylus brachymystacis | 14,767 | 30.6 | 35.2 | 15.1 | 19.1 | NC_031337 |
| Gyrodactylidea | Gyrodactylidae | Gyrodactylus derjavinoides | 14,741 | 33.1 | 35.1 | 14.2 | 17.6 | NC_010976 |
| Gyrodactylidea | Gyrodactylidae | Gyrodactylus gurleyi | 14,771 | 29.2 | 42.9 | 11.1 | 16.9 | KU659806 |
| Gyrodactylidea | Gyrodactylidae | Gyrodactylus kobayashii | 14,786 | 29.7 | 41.9 | 11.1 | 17.3 | NC_030050 |
| Gyrodactylidea | Gyrodactylidae | Gyrodactylus nyanzae | 14,885 | 32.2 | 47.9 | 6.7 | 13.2 | MG970256 |
| Gyrodactylidea | Gyrodactylidae | Gyrodactylus parvae | 14,702 | 32.2 | 41.3 | 10.7 | 15.8 | NC_031438 |
| Gyrodactylidea | Gyrodactylidae | Gyrodactylus salaris | 14,790 | 29.8 | 32.7 | 17.1 | 20.4 | EF527269 |
| Dactylogyridea | Diplectanidae | Lamellodiscus spari | 14,614 | 31.5 | 45.3 | 7.9 | 15.4 | MH328204 |
| Gyrodactylidea | Gyrodactylidae | Paragyrodactylus variegatus | 14,517 | 30.4 | 45.8 | 9.5 | 14.2 | NC_024754 |
| Tetraonchidea | Tetraonchoididae | Paratetraonchoides inermis | 14,654 | 36.1 | 46.5 | 6.0 | 11.4 | KY856918 |
| Dactylogyridea | Diplectanidae | Pseudorhabdosynochus yangjiangensis | 12,458 | 29.9 | 44.8 | 8.0 | 17.4 | JQ038231 |
| Dactylogyridea | Ancyrocephalidae | Scutogyrus longicornis | 14,241 | 22.4 | 43.4 | 22.9 | 11.3 | MT447060 |
| Dactylogyridea | Dactylogyridae | Tetrancistrum nebulosi | 13,392 | 27.2 | 38.2 | 13.1 | 21.5 | NC_018031 |
| Tetraonchidea | Tetraonchidae | Tetraonchus monenteron | 14,791 | 26.2 | 47.2 | 11.2 | 15.4 | NC_046757 |
| Dactylogyridea | Ancylodiscoididae | Thaparocleidus asoti | 17,493 | 32.1 | 46.6 | 7.3 | 14.1 | NC_053548 |
| Dactylogyridea | Ancylodiscoididae | Thaparocleidus varicus | 14,088 | 30.1 | 46.8 | 7.6 | 15.5 | NC_053547 |
| Region | Size (bp) | A (%) | T (%) | G (%) | C (%) | A + T (%) | AT-Skew | GC-Skew |
|---|---|---|---|---|---|---|---|---|
| Mitogenome | 13,265/13,984 | 25.4/25.8 | 41.1/38.2 | 18.9/19.1 | 14.6/17.0 | 66.5/63.9 | −0.24/−0.19 | 0.13/0.06 |
| cox1 | 1563/1563 | 23.8/23.0 | 39.9/36.5 | 20.0/21.7 | 16.3/18.8 | 63.7/59.6 | −0.25/−0.23 | 0.10/0.07 |
| cox2 | 582/630 | 26.5/27.3 | 37.8/34.0 | 20.6/22.2 | 15.1/16.5 | 64.3/61.3 | −0.18/−0.11 | 0.15/0.15 |
| atp6 | 546/510 | 23.6/21.8 | 43.0/39.0 | 17.6/20.8 | 15.8/18.4 | 66.7/60.8 | −0.29/−0.28 | 0.05/0.06 |
| cox3 | 651/651 | 24.7/23.7 | 43.0/40.3 | 18.7/19.4 | 13.5/16.7 | 67.7/63.9 | −0.27/−0.26 | 0.16/0.07 |
| nad3 | 354/354 | 24.0/26.8 | 45.2/41.2 | 18.6/17.8 | 12.2/14.1 | 69.2/68.1 | −0.31/−0.21 | 0.21/0.12 |
| nad1 | 900/900 | 22.9/25.2 | 40.8/38.4 | 21.9/19.0 | 14.4/17.3 | 63.7/63.7 | −0.28/−0.21 | 0.20/0.05 |
| nad5 | 1542/1542 | 24.1/23.8 | 44.8/38.4 | 16.5/18.6 | 14.7/17.3 | 68.8/64.2 | −0.30/−0.26 | 0.06/0.04 |
| nad4 | 1221/1167 | 24.6/24.1 | 43.5/39.4 | 17.0/18.3 | 15.0/18.2 | 68.1/63.5 | −0.28/−0.24 | 0.06/0.01 |
| nad4L | 249/249 | 22.5/23.7 | 47.4/42.6 | 16.9/16.9 | 13.3/16.9 | 69.9/66.3 | −0.36/−0.28 | 0.12/0.00 |
| nad6 | 453/453 | 21.9/22.5 | 45.9/43.9 | 19.2/19.9 | 13.0/13.7 | 67.8/66.5 | −0.36/−0.32 | 0.19/0.18 |
| cob | 1104/1083 | 24.5/25.9 | 39.1/37.0 | 20.6/19.4 | 15.9/17.7 | 63.6/62.9 | −0.23/−0.18 | 0.13/0.04 |
| nad2 | 861/864 | 24.6/22.3 | 46.1/42.5 | 17.5/18.9 | 11.7/16.3 | 70.7/64.8 | −0.30/−0.31 | 0.20/0.07 |
| tRNAs | 1429/1436 | 29.4/28.8 | 35.4/34.7 | 20.2/21.0 | 15.0/15.5 | 64.8/63.5 | −0.09/−0.09 | 0.15/0.15 |
| rRNAs | 1662/1655 | 30.0/29.8 | 36.9/36.4 | 18.6/18.0 | 14.4/15.8 | 66.9/66.2 | −0.10/−0.10 | 0.13/0.06 |
| PCGs | 10,026/9966 | 24.1/24.1 | 42.5/39.1 | 18.8/19.6 | 14.6/17.3 | 66.6/63.2 | −0.28/−0.24 | 0.12/0.06 |
| Gene | Position | Size | Initiation | Termination | Anticodon | Overlapping | Intergenic | Strand | |
|---|---|---|---|---|---|---|---|---|---|
| cox1 | 1/1 | 1563/1563 | 1563/1563 | ATG/ATG | TAA/TAA | 7/6 | +/+ | ||
| trnT | 1571/1568 | 1635/1633 | 65/66 | tgt/tgt | +/+ | ||||
| rrnL | 1636/1634 | 2588/2576 | 953/943 | 6/10 | +/+ | ||||
| trnC | 2583/2567 | 2648/2634 | 66/68 | gca/gca | 1/- | +/+ | |||
| rrnS | 2650/2635 | 3364/3348 | 715/714 | -/35 | 15/- | +/+ | |||
| cox2 | 3380/3314 | 3961/3943 | 582/630 | ATG/ATG | TAG/TAG | 2/8 | +/+ | ||
| trnE | 3964/3952 | 4030/4016 | 67/65 | ttc/ttc | +/+ | ||||
| nad6 | 4031/4017 | 4483/4469 | 453/453 | ATG/ATG | TAG/TAA | 4/10 | +/+ | ||
| trnY | 4488/4480 | 4548/4543 | 61/64 | gta/gta | 6/5 | +/+ | |||
| trnL1 | 4555/4549 | 4621/4615 | 67/67 | tag/tag | +/+ | ||||
| trnS2 | 4622/4616 | 4686/4679 | 65/64 | tga/tga | 1/- | +/+ | |||
| trnL2 | 4688/4680 | 4753/4745 | 66/66 | taa/taa | -/1 | +/+ | |||
| trnR | 4754/4745 | 4816/4810 | 63/66 | tcg/tcg | 2/1 | +/+ | |||
| nad5 | 4819/4812 | 6360/6353 | 1542/1542 | ATG/ATG | TAA/TAG | 80/80 | +/+ | ||
| trnG | 6441/6434 | 6506/6499 | 66/66 | tcc/tcc | 2/2 | +/+ | |||
| cox3 | 6509/6502 | 7159/7152 | 651/651 | ATG/ATG | TAA/TAA | 6/1 | +/+ | ||
| trnH | 7166/7154 | 7228/7216 | 63/63 | gtg/gtg | 20/- | -/2 | +/+ | ||
| cob | 7209/7219 | 8312/8301 | 1104/1083 | ATG/ATG | TAG/TAA | 1/1 | +/+ | ||
| nad4L | 8312/8301 | 8560/8549 | 249/249 | GTG/ATG | TAG/TAG | 28/- | -/29 | +/+ | |
| nad4 | 8533/8579 | 9753/9745 | 1221/1167 | GTG/ATA | TAA/TAG | 14/15 | +/+ | ||
| trnF | 9768/9761 | 9835/9823 | 68/63 | gaa/gaa | 5/3 | +/+ | |||
| trnM | 9831/9821 | 9897/9887 | 67/67 | cat/cat | 36/- | +/+ | |||
| atp6 | 9862/9888 | 10,407/10,397 | 546/510 | ATG/ATG | TAG/TAG | 2/6 | +/+ | ||
| nad2 | 10,410/10,404 | 11,270/11,267 | 861/864 | GTG/ATG | TAA/TAG | 6/18 | +/+ | ||
| trnD | 11,551/11,286 | 11,616/11,351 | 66/66 | gtc/gtc | +/+ | ||||
| nad1 | 11,617/11,352 | 12,516/12,251 | 900/900 | ATG/ATG | TAA/TGA | 4/29 | +/+ | ||
| trnV | 11,277/12,281 | 11,342/12,344 | 66/64 | tac/tac | 11/413 | +/+ | |||
| trnN | 12,521/12,758 | 12,586/12,823 | 66/66 | gtt/gtt | 6/335 | +/+ | |||
| trnA | 11,354/13,159 | 11,418/13,225 | 65/67 | tgc/tgc | 60/8 | +/+ | |||
| trnQ | 11,479/13,234 | 11,541/13,297 | 63/64 | ttg/ttg | 9/11 | +/+ | |||
| trnP | 12,593/13,309 | 12,658/13,375 | 66/67 | tgg/tgg | 1/1 | +/+ | |||
| trnI | 12,658/13,375 | 12,723/13,440 | 66/66 | gat/gat | 1/- | +/+ | |||
| trnK | 12,725/13,441 | 12,790/13,507 | 66/67 | ctt/ctt | +/+ | ||||
| nad3 | 12,791/13,508 | 13,144/13,861 | 354/354 | ATG/GTG | TAG/TAG | -/1 | +/+ | ||
| trnS1 | 13,145/13,861 | 13,201/13,918 | 57/42 | gct/agc | +/+ | ||||
| trnW | 13,202/13,919 | 13,265/13,984 | 64/66 | tca/tca | +/+ | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Shan, B.; Liu, Y.; Wang, L.; Wu, Q.; Luo, Z.; Sun, D. Complete Mitochondrial Genome of Two Ectoparasitic Capsalids (Platyhelminthes: Monogenea: Monopisthocotylea): Gene Content, Composition, and Rearrangement. Genes 2022, 13, 1376. https://doi.org/10.3390/genes13081376
Yang C, Shan B, Liu Y, Wang L, Wu Q, Luo Z, Sun D. Complete Mitochondrial Genome of Two Ectoparasitic Capsalids (Platyhelminthes: Monogenea: Monopisthocotylea): Gene Content, Composition, and Rearrangement. Genes. 2022; 13(8):1376. https://doi.org/10.3390/genes13081376
Chicago/Turabian StyleYang, Changping, Binbin Shan, Yan Liu, Liangming Wang, Qiaer Wu, Zhengli Luo, and Dianrong Sun. 2022. "Complete Mitochondrial Genome of Two Ectoparasitic Capsalids (Platyhelminthes: Monogenea: Monopisthocotylea): Gene Content, Composition, and Rearrangement" Genes 13, no. 8: 1376. https://doi.org/10.3390/genes13081376