
Transcriptional Interference Regulates the Evolutionary Development of Speech


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Radiology and MRI Analyses
2.2. Primate Tissues
2.3. RT-PCR Characterisation of TOSPEAK Transcripts
2.4. Comparative Genome Analyses
2.5. Luciferase Assays
2.6. BAC Transgenic Analyses
3. Results
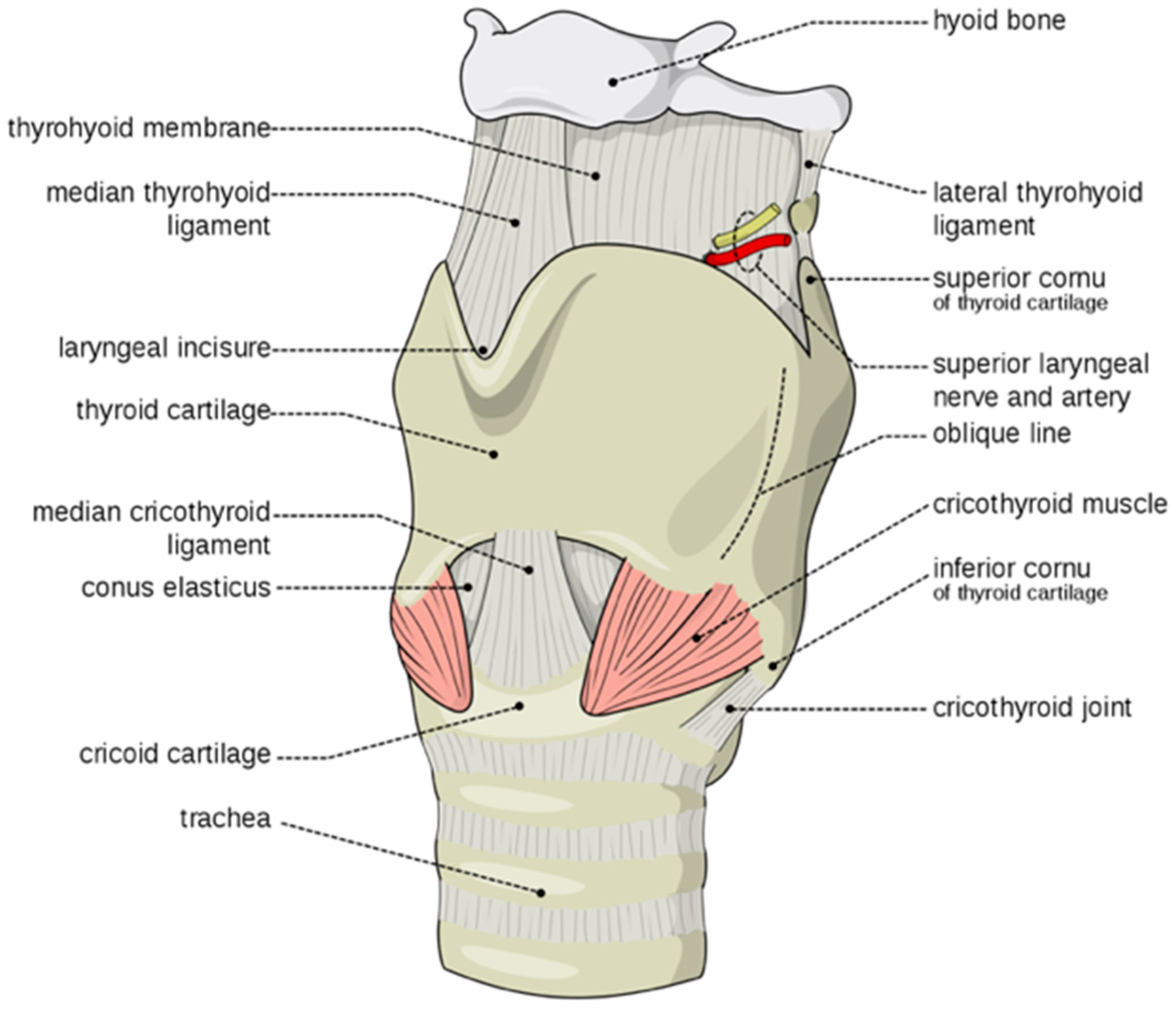
3.1. Speech Impairment Associated with Evolutionary Retrograde Laryngeal Configuration
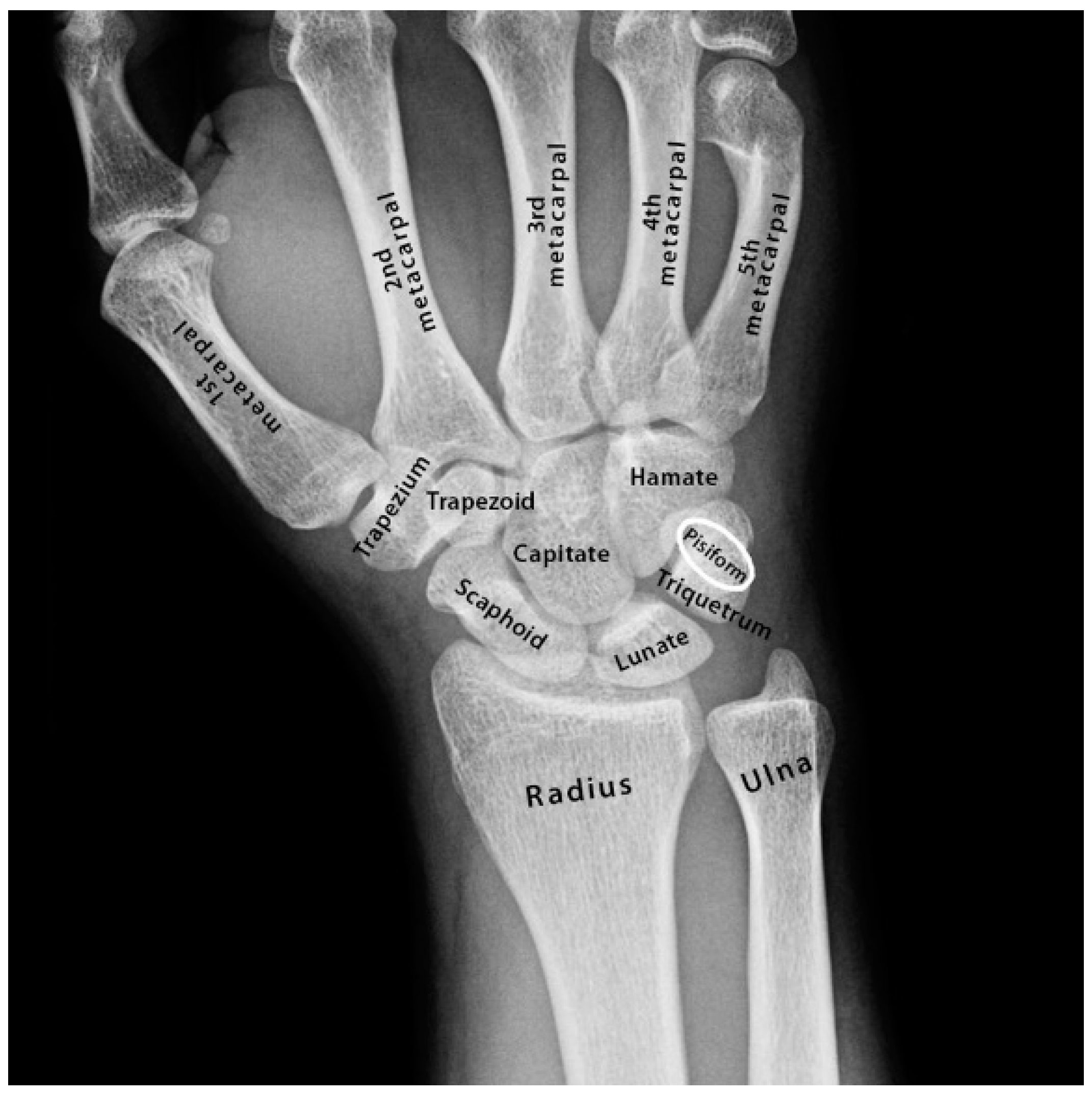
3.2. Restricted Wrist Rotation Associated with Evolutionary Retrograde Wrist Morphology
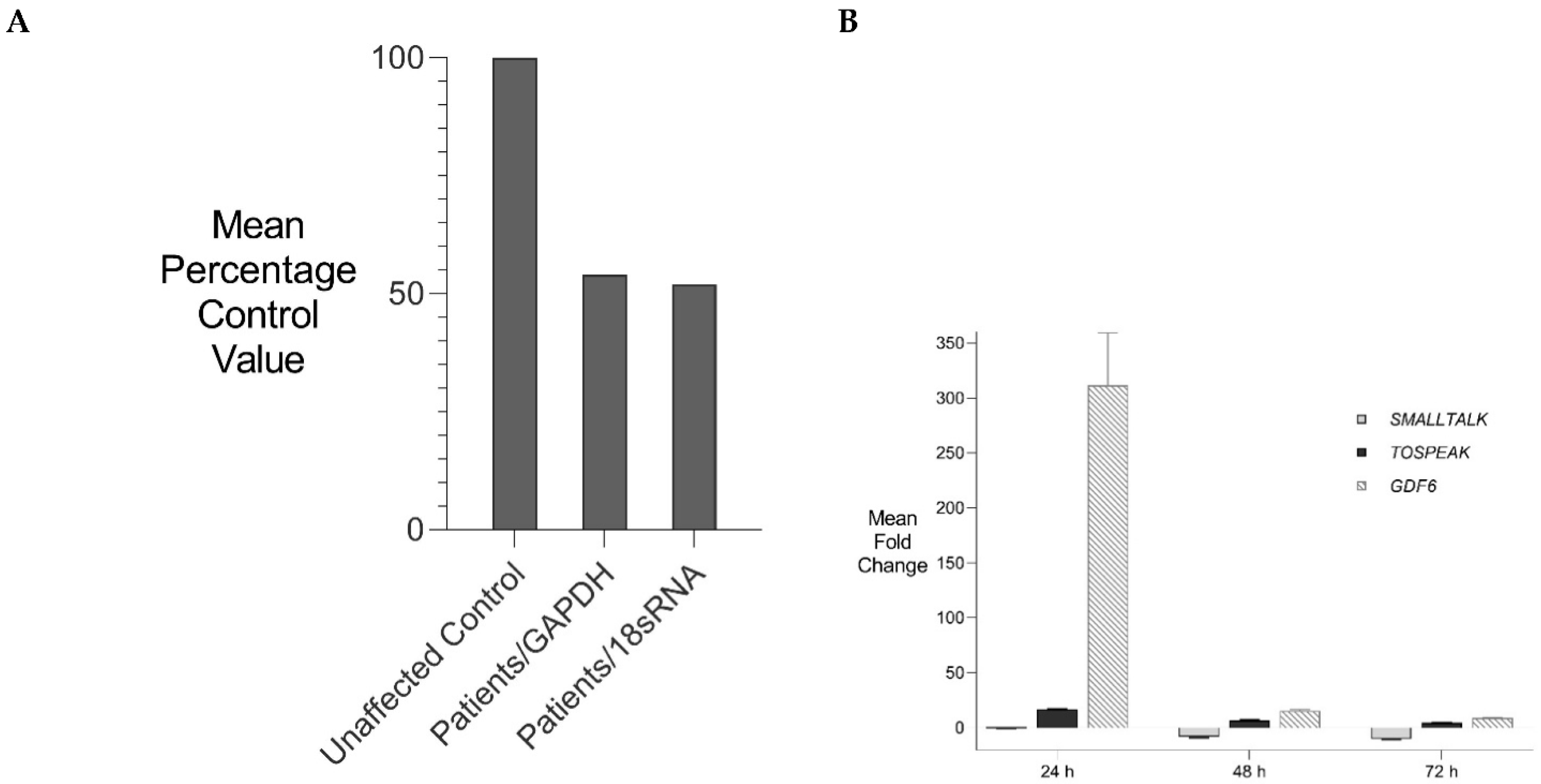
3.3. Primate-Specific Gene TOSPEAK Disrupted in Speech Impaired Family
3.4. TOSPEAK Proximal Core Promoter and Start Site Perfectly Conserved
3.5. Incremental Increase in TOSPEAK Promoter Strength in Higher Hominids with Human Having the Strongest Promoter
3.6. TOSPEAK Is Transcribed across GDF6 Long-Range Enhancers
4. Discussion
5. Conclusions
- Pisiform and its elongation (retrogression) and extension into the radio-ulnar joint that restricted wrist rotation.
- Hyoid-thyroid ligament of larynx retarded (retrogressed) laryngeal descent causing speech impairment.
- Vocal cords that reduced their elasticity and retarded their pubertal elongation and the progressive protrusion/prominence of the thyroid cartilage that increased the severity of speech impairment in the affected family (Figure 4C,D).
- Emergence of TOSPEAK transcription in primates across these GDF6 enhancers
- Incremental increases in TOSPEAK promoter strength in hominoids
- Increased descent and expansion of the larynx and the incremental retraction of the pisiform.
- Conserved long-range tissue-specific enhancers regulate the expression of the ancient GDF6 gene in its role in regulating the morphology, elasticity and structure of the ancient larynx and wrist joints.
- The de novo birth of the TOSPEAK gene in primates across pre-existing GDF6 enhancers (DJE1, ECR5 and DJE2) for laryngeal and wrist development (Figure 6).
- Perfect conservation in primates of the
- Incremental increases in TOSPEAK promoter strength in hominoids and hominids with the strongest TOSPEAK promoter in human (Figure 7B) drives incremental increases in the transcription of TOSPEAK across GDF6 enhancers in higher hominoids.
- Phylogenetic increases in transcriptional interference of site-specific GDF6 enhancers positively upregulates the transcription of GDF6 at those sites during development [12].
- Increased flexibility of the hyoid thyroid ligament permitted phylogenetic descent of the larynx thus removing hyoid cartilage constraints on the flexibility and utility of the larynx and the tongue in hominids (Figure 4).
- Increased flexibility of the larynx and tongue increased their utility and the capacity to speak.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fitch, W.T. Genes, language, cognition, and culture: Towards productive inquiry. Hum. Biol. 2011, 83, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holden, C. The origin of speech. Science 2004, 303, 1316–1319. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Oishi, T.; Suzuki, J.; Matsuda, K.; Takahashi, T. Development of the supralaryngeal vocal tract in Japanese macaques: Implications for the evolution of the descent of the larynx. Am. J. Phys. Anthropol. 2008, 135, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Fitch, W.T.; Reby, D. The descended larynx is not uniquely human. Proc. Biol. Sci. 2001, 268, 1669–1675. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, D.E.; McCarthy, R.C.; Hiiemae, K.M.; Palmer, J.B. Ontogeny of postnatal hyoid and larynx descent in humans. Arch. Oral Biol. 2001, 46, 117–128. [Google Scholar] [CrossRef]
- Lieberman, P. The evolution of human speech—Its anatomical and neural bases. Curr. Anthropol. 2007, 48, 39–66. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, F.A. Primate Locomotion; Academic Press: New York, NY, USA, 1974. [Google Scholar]
- Tuttle, R. The Functional and Evolutionary Biology of Primates; New Brunswick (USA) Aldine Transaction; Routledge: New York, NY, USA, 2007. [Google Scholar]
- Kjosness, K.M.; Reno, P.L. Identifying the homology of the short human pisiform and its lost ossification center. Evodevo 2019, 10, 32. [Google Scholar] [CrossRef]
- Settle, S.H.; Rountree, R.B., Jr.; Sinha, A.; Thacker, A.; Higgins, K.; Kingsley, D.M. Multiple joint and skeletal patterning defects caused by single and double mutations in the mouse Gdf6 and Gdf5 genes. Dev. Biol. 2003, 254, 116–130. [Google Scholar] [CrossRef] [Green Version]
- Mortlock, D.P.; Guenther, C.; Kingsley, D.M. A general approach for identifying distant regulatory elements applied to the Gdf6 gene. Genome Res. 2003, 13, 2069–2081. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.; Zhao, Z.; Eapen, V.; Clarke, R.A. siRNA Mediate RNA Interference Concordant with Early On-Target Transient Transcriptional Interference. Genes 2021, 12, 1290. [Google Scholar] [CrossRef]
- Clarke, R.A.; Fang, Z.; Murrell, D.; Sheriff, T.; Eapen, V. GDF6 Knockdown in a Family with Multiple Synostosis Syndrome and Speech Impairment. Genes 2021, 12, 1354. [Google Scholar] [CrossRef] [PubMed]
- Tassabehji, M.; Fang, Z.M.; Hilton, E.N.; McGaughran, J.; Zhao, Z.; de Bock, C.E.; Howard, E.; Malass, M.; Donnai, D.; Diwan, A.; et al. Mutations in GDF6 are associated with vertebral segmentation defects in Klippel-Feil syndrome. Hum. Mutat. 2008, 29, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic. Acids. Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- De Bock, C.E.; Lin, Z.; Itoh, T.; Morris, D.; Murrell, G.; Wang, Y. Inhibition of urokinase receptor gene expression and cell invasion by anti-uPAR DNAzymes in osteosarcoma cells. FEBS J. 2005, 272, 3572–3582. [Google Scholar] [CrossRef] [PubMed]
- Portnoy, M.E.; McDermott, K.J.; Antonellis, A.; Margulies, E.H.; Prasad, A.B.; Kingsley, D.M.; Green, E.D.; Mortlock, D.P. Detection of potential GDF6 regulatory elements by multispecies sequence comparisons and identification of a skeletal joint enhancer. Genomics 2005, 86, 295–305. [Google Scholar] [CrossRef]
- DiLeone, R.J.; Marcus, G.A.; Johnson, M.D.; Kingsley, D.M. Efficient studies of long-distance Bmp5 gene regulation using bacterial artificial chromosomes. Proc. Natl. Acad. Sci. USA 2000, 97, 1612–1617. [Google Scholar] [CrossRef] [Green Version]
- Chandler, R.L.; Chandler, K.J.; McFarland, K.A.; Mortlock, D.P. Bmp2 transcription in osteoblast progenitors is regulated by a distant 3′ enhancer located 156.3 kilobases from the promoter. Mol. Cell. Biol. 2007, 27, 2934–2951. [Google Scholar] [CrossRef] [Green Version]
- Clarke, R.A.; Davis, P.J.; Tonkin, J. Klippel-Feil syndrome associated with malformed larynx. Case report. Ann. Otol. Rhinol. Laryngol. 1994, 103, 201–207. [Google Scholar] [CrossRef]
- Clarke, R.A.; Singh, S.; McKenzie, H.; Kearsley, J.H.; Yip, M.Y. Familial Klippel-Feil syndrome and paracentric inversion inv(8)(q22.2q23.3). Am. J. Hum. Genet. 1995, 57, 1364–1370. [Google Scholar]
- Mattick, J.S. The genetic signatures of noncoding RNAs. PLoS Genet. 2009, 5, e1000459. [Google Scholar] [CrossRef]
- Reed, N.P.; Mortlock, D.P. Identification of a distant cis-regulatory element controlling pharyngeal arch-specific expression of zebrafish gdf6a/radar. Dev. Dyn. 2010, 239, 1047–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bushehri, A.; Barez, M.R.; Mansouri, S.K.; Biglarian, A.; Ohadi, M. Genome-wide identification of human- and primate-specific core promoter short tandem repeats. Gene 2016, 587, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Ohadi, M.; Valipour, E.; Ghadimi-Haddadan, S.; Namdar-Aligoodarzi, P.; Bagheri, A.; Kowsari, A.; Rezazadeh, M.; Darvish, H.; Kazeminasab, S. Core promoter short tandem repeats as evolutionary switch codes for primate speciation. Am. J. Primatol. 2015, 77, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Sawaya, S.M.; Lennon, D.; Buschiazzo, E.; Gemmell, N.; Minin, V.N. Measuring microsatellite conservation in mammalian evolution with a phylogenetic birth-death model. Genome Biol. Evol. 2012, 4, 636–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomberk, G.; Urrutia, R. The family feud: Turning off Sp1 by Sp1-like KLF proteins. Biochem. J. 2005, 392 Pt 1, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pregizer, S.; Mortlock, D.P. Control of BMP gene expression by long-range regulatory elements. Cytokine Growth Factor Rev. 2009, 20, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Bhargav, D.; Wei, A.; Williams, L.A.; Tao, H.; Ma, D.D.; Diwan, A.D. BMP-13 emerges as a potential inhibitor of bone formation. Int. J. Biol. Sci. 2009, 5, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Miles, J.; Mitchell, J.A.; Chakalova, L.; Goyenechea, B.; Osborne, C.S.; O’Neill, L.; Tanimoto, K.; Engel, J.D.; Fraser, P. Intergenic transcription, cell-cycle and the developmentally regulated epigenetic profile of the human beta-globin locus. PLoS ONE 2007, 2, e630. [Google Scholar] [CrossRef] [Green Version]
- Debrand, E.; Chakalova, L.; Miles, J.; Dai, Y.F.; Goyenechea, B.; Dye, S.; Osborne, C.S.; Horton, A.; Harju-Baker, S.; Pink, R.C.; et al. An intergenic non-coding RNA promoter required for histone modifications in the human beta-globin chromatin domain. PLoS ONE 2019, 14, e0217532. [Google Scholar] [CrossRef] [Green Version]
- Kaessmann, H. Origins, evolution, and phenotypic impact of new genes. Genome Res. 2010, 20, 1313–1326. [Google Scholar] [CrossRef] [Green Version]
- Clarke, R.A.; Lee, S.; Eapen, V. Pathogenetic model for Tourette syndrome delineates overlap with related neurodevelopmental disorders including Autism. Transl. Psychiatry 2012, 2, e158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eapen, V.; Clarke, R.A. Autism spectrum disorders: From genotypes to phenotypes. Front. Hum. Neurosci. 2014, 8, 914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, R.A.; Eapen, V. Balance within the Neurexin Trans-Synaptic Connexus Stabilizes Behavioral Control. Front. Hum. Neurosci. 2014, 8, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Z.; Eapen, V.; Clarke, R.A. CTNNA3 discordant regulation of nested LRRTM3, implications for autism spectrum disorder and Tourette syndrome. Meta Gene 2017, 5, 43–48. [Google Scholar] [CrossRef]
- Clarke, R.A.; Eapen, V. LRRTM4 terminal exon duplicated in family with Tourette syndrome, autism and ADHD. Genes 2022, 13, 66. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human | Exon1 | 1A | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 8A | 9 | 9A | GenBank |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variants | 55 bp | 83 bp | 71 bp | 163 bp | 155 bp | 46 bp | 34 bp | 110 bp | 160 bp | 59 bp | 125 bp | 95 bp | |
| 1 | * | * | * | * | * | * | GU295153 | ||||||
| 2 | * | * | * | * | * | GU295154 | |||||||
| 3 | * | * | * | * | * | GU295155 | |||||||
| 4 | * | * | * | * | GU295156 | ||||||||
| 5 | * | * | * | * | GU295157 | ||||||||
| 6 | * | +5 | * | GU295158 | |||||||||
| 7 | * | +5 | −101 | * | GU295159 | ||||||||
| 8 | * | * | * | * | * | * | GU295160 | ||||||
| 9 | * | GU295161 | |||||||||||
| 10 | * | * | * | * | GU295162 | ||||||||
| 11 | * | * | * | * | GU295163 | ||||||||
| 12 | * | * | GU295164 | ||||||||||
| Other Primate Variants | |||||||||||||
| Chimp | * | * | * | * | GU295165 | ||||||||
| Gorilla | * | * | * | −101 | * | GU295166 | |||||||
| Orangutan | * | +185 | GU295167 | ||||||||||
| OWM1 | 28 | * | * | +5 | −101 | * | +192 | GU295168 | |||||
| OWM2 | 28 | * | * | +5 | −101 | * | −44 | * | GU295169 | ||||
| NWM | * | * | +192 | GU295170 |
| BAC | Method | Total Embryos | Transgenics | Expressed in Interdigits | Expressed in Proximal Wrist/Ankle | Expressed in Distal Wrist/Ankle |
|---|---|---|---|---|---|---|
| RP23-11F19 | Co-injection | 71 | 18 | 7 | 0 | 0 |
| RP23-444O1 | Co-injection | 46 | 15 | 0 | 0 | 0 |
| RP23-56O6 | Co-injection | 84 | 14 | 0 | 6 | 0 |
| “ | Modified BAC | 17 | 2 | 0 | 2 | 0 |
| RP23-333E9 | Co-injection | 14 | 4 | 0 | 0 | 3 |
| “ | Modified BAC | 16 | 2 | 0 | 0 | 1 |
| RP23-354G12 | Co-injection | 24 | 2 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mortlock, D.P.; Fang, Z.-M.; Chandler, K.J.; Hou, Y.; Bickford, L.R.; de Bock, C.E.; Eapen, V.; Clarke, R.A. Transcriptional Interference Regulates the Evolutionary Development of Speech. Genes 2022, 13, 1195. https://doi.org/10.3390/genes13071195
Mortlock DP, Fang Z-M, Chandler KJ, Hou Y, Bickford LR, de Bock CE, Eapen V, Clarke RA. Transcriptional Interference Regulates the Evolutionary Development of Speech. Genes. 2022; 13(7):1195. https://doi.org/10.3390/genes13071195
Chicago/Turabian StyleMortlock, Douglas P., Zhi-Ming Fang, Kelly J. Chandler, Yue Hou, Lissett R. Bickford, Charles E. de Bock, Valsamma Eapen, and Raymond A. Clarke. 2022. "Transcriptional Interference Regulates the Evolutionary Development of Speech" Genes 13, no. 7: 1195. https://doi.org/10.3390/genes13071195