
Insight from OPN1LW Gene Haplotypes into the Cause and Prevention of Myopia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Human Subjects
2.3. Opsin Gene Sequencing
2.4. Minigenes
2.5. Splicing Assays
2.6. The MassArray Assay
2.7. Statistics
3. Results
3.1. The Relationship between Xq28 Opsin Gene Haplotypes and Refractive Error
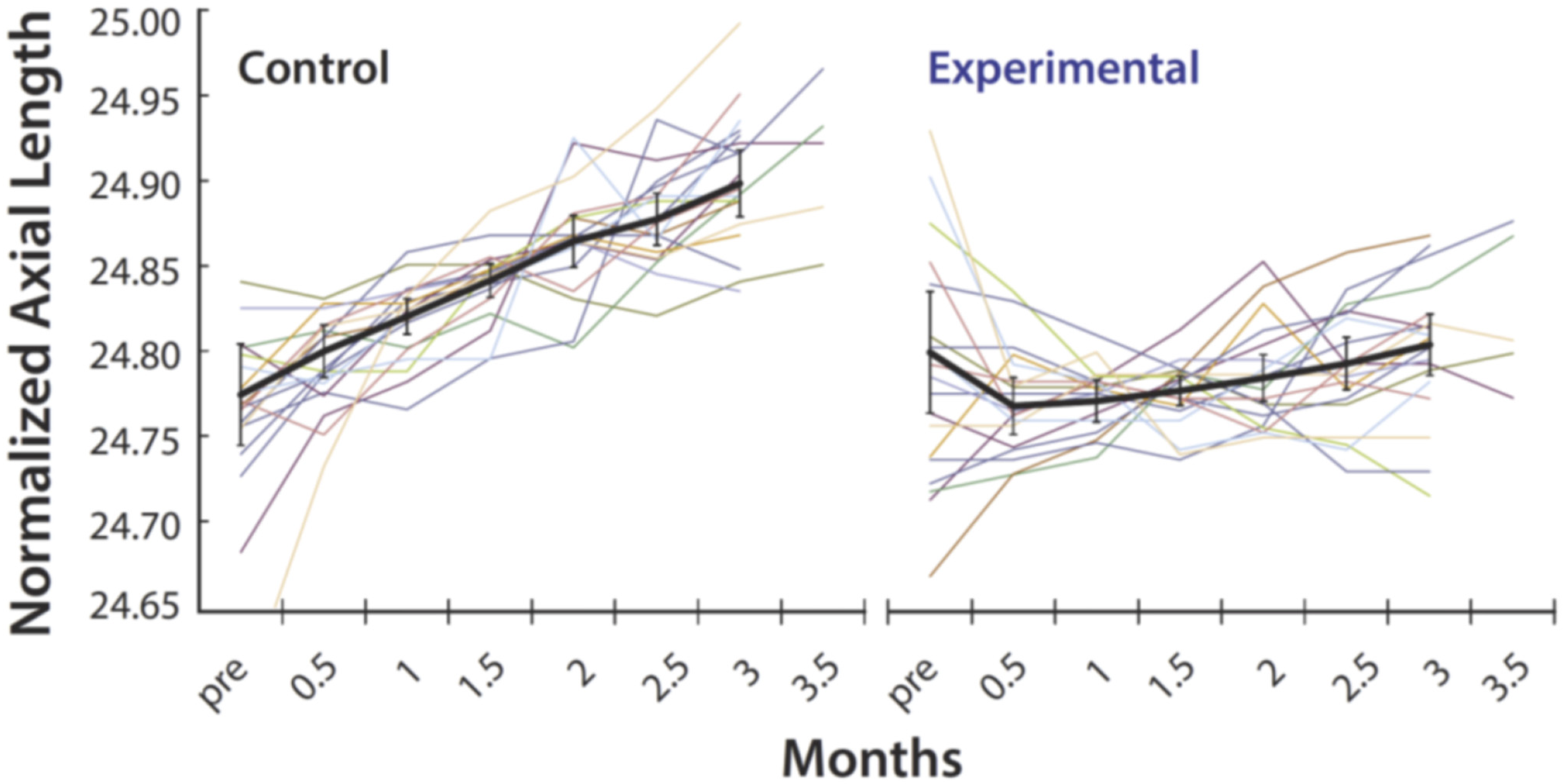
3.2. Myopia Prevention Using Spectacles That Control the Spatial Distribution of Light Reaching the Retina
4. Discussion
4.1. Implications of Xq28 Opsin Gene Haplotypes for Females
4.2. Why Was OPN1LW Not Identified in GWAS Searches for Myopia Genes?
4.3. An Optimized Version of the Myopia Reducing Spectacles
4.4. An Unorthodox Theory of Myopia
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dolgin, E. The myopia boom. Nature 2015, 519, 276–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holden, B.A.; Fricke, T.R.; Wilson, D.A.; Jong, M.; Naidoo, K.S.; Sankaridurg, P.; Wong, T.Y.; Naduvilath, T.J.; Resnikoff, S. Global Prevalence of Myopia and High Myopia and Temporal Trends from 2000 through 2050. Ophthalmology 2016, 123, 1036–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karthikeyan, S.K.; Ashwini, D.L.; Priyanka, M.; Nayak, A.; Biswas, S. Physical activity, time spent outdoors, and near work in relation to myopia prevalence, incidence, and progression: An overview of systematic reviews and meta-analyses. Indian J. Ophthalmol. 2022, 70, 728–739. [Google Scholar] [CrossRef]
- Holden, B.; Sankaridurg, P.; Smith, E.; Aller, T.; Jong, M.; He, M. Myopia, an underrated global challenge to vision: Where the current data takes us on myopia control. Eye (Lond. Engl.) 2014, 28, 142–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitale, S.; Sperduto, R.D.; Ferris, F.L., III. Increased prevalence of myopia in the United States between 1971–1972 and 1999–2004. Arch. Ophthalmol. 2009, 127, 1632–1639. [Google Scholar] [CrossRef] [Green Version]
- Flitcroft, D.I. The complex interactions of retinal, optical and environmental factors in myopia aetiology. Prog. Retin. Eye Res. 2012, 31, 622–660. [Google Scholar] [CrossRef]
- Hysi, P.G.; Choquet, H.; Khawaja, A.P.; Wojciechowski, R.; Tedja, M.S.; Yin, J.; Simcoe, M.J.; Patasova, K.; Mahroo, O.A.; Thai, K.K.; et al. Meta-analysis of 542,934 subjects of European ancestry identifies new genes and mechanisms predisposing to refractive error and myopia. Nat. Genet. 2020, 52, 401–407. [Google Scholar] [CrossRef]
- Hysi, P.G.; Wojciechowski, R.; Rahi, J.S.; Hammond, C.J. Genome-wide association studies of refractive error and myopia, lessons learned, and implications for the future. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3344–3351. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowski, R.; Hysi, P.G. Focusing in on the complex genetics of myopia. PLoS Genet. 2013, 9, e1003442. [Google Scholar] [CrossRef] [Green Version]
- Kiefer, A.K.; Tung, J.Y.; Do, C.B.; Hinds, D.A.; Mountain, J.L.; Francke, U.; Eriksson, N. Genome-wide analysis points to roles for extracellular matrix remodeling, the visual cycle, and neuronal development in myopia. PLoS Genet. 2013, 9, e1003299. [Google Scholar] [CrossRef] [Green Version]
- McClements, M.; Davies, W.I.; Michaelides, M.; Young, T.; Neitz, M.; Maclaren, R.E.; Moore, A.T.; Hunt, D.M. Variations in opsin coding sequences cause X-linked cone dysfunction syndrome with myopia and dichromacy. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1361–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gao, B.; Guan, L.; Xiao, X.; Zhang, J.; Li, S.; Jiang, H.; Jia, X.; Yang, J.; Guo, X.; et al. Unique Variants in OPN1LW Cause Both Syndromic and Nonsyndromic X-Linked High Myopia Mapped to MYP1. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4150–4155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buena-Atienza, E.; Ruther, K.; Baumann, B.; Bergholz, R.; Birch, D.; De Baere, E.; Dollfus, H.; Greally, M.T.; Gustavsson, P.; Hamel, C.P.; et al. De novo intrachromosomal gene conversion from OPN1MW to OPN1LW in the male germline results in Blue Cone Monochromacy. Sci. Rep. 2016, 6, 28253. [Google Scholar] [CrossRef] [Green Version]
- Greenwald, S.; Kuchenbecker, J.A.; Rowlan, J.S.; Neitz, J.; Neitz, M. Role of a dual splicing and amino acid code in myopia, cone dysfunction and cone dystrophy associated with L/M opsin interchange mutations. TVST 2017, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orosz, O.; Rajta, I.; Vajas, A.; Takacs, L.; Csutak, A.; Fodor, M.; Kolozsvari, B.; Resch, M.; Senyi, K.; Lesch, B.; et al. Myopia and Late-Onset Progressive Cone Dystrophy Associate to LVAVA/MVAVA Exon 3 Interchange Haplotypes of Opsin Genes on Chromosome X. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1834–1842. [Google Scholar] [CrossRef] [Green Version]
- Neitz, M.; Neitz, J. Intermixing the OPN1LW and OPN1MW Genes Disrupts the Exonic Splicing Code Causing an Array of Vision Disorders. Genes 2021, 12, 1180. [Google Scholar] [CrossRef]
- Winderickx, J.; Battisti, L.; Hibibya, Y.; Motulsky, A.G.; Deeb, S.S. Haplotype diversity in the human red and green opsin genes: Evidence for frequent sequence exchange in exon 3. Hum. Mol. Genet. 1993, 2, 1413–1421. [Google Scholar] [CrossRef]
- Carroll, J.; Neitz, M.; Neitz, J. Estimates of L:M cone ratio from ERG flicker photometry and genetics. J. Vis. 2002, 2, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Verrelli, B.C.; Tishkoff, S.A. Signatures of selection and gene conversion associated with human color vision variation. Am. J. Hum. Genet. 2004, 75, 363–375. [Google Scholar] [CrossRef] [Green Version]
- Ueyama, H.; Muraki-Oda, S.; Yamade, S.; Tanabe, S.; Yamashita, T.; Shichida, Y.; Ogita, H. Unique haplotype in exon 3 of cone opsin mRNA affects splicing of its precursor, leading to congenital color vision defect. Biochem. Biophys. Res. Commun. 2012, 424, 152–157. [Google Scholar] [CrossRef]
- Gardner, J.C.; Liew, G.; Quan, Y.H.; Ermetal, B.; Ueyama, H.; Davidson, A.E.; Schwarz, N.; Kanuga, N.; Chana, R.; Maher, E.R.; et al. Three different cone opsin gene array mutational mechanisms with genotype-phenotype correlation and functional investigation of cone opsin variants. Hum. Mutat. 2014, 35, 1354–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, E.J.; Wilk, M.; Langlo, C.S.; Kasilian, M.; Ring, M.; Hufnagel, R.B.; Dubis, A.M.; Tee, J.J.; Kalitzeos, A.; Gardner, J.C.; et al. Cone Photoreceptor Structure in Patients With X-Linked Cone Dysfunction and Red-Green Color Vision Deficiency. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3853–3863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, E.J.; Kalitzeos, A.; Kasilian, M.; Gardner, J.C.; Neitz, J.; Hardcastle, A.J.; Neitz, M.; Carroll, J.; Michaelides, M. Residual Cone Structure in Patients with X-Linked Cone Opsin Mutations. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4238–4248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathans, J.; Thomas, D.; Hogness, D.S. Molecular genetics of human color vision: The genes encoding blue, green, and red pigments. Science 1986, 232, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollrath, D.; Nathans, J.; Davis, R.W. Tandem array of human visual pigment genes at Xq28. Science 1988, 240, 1669–1672. [Google Scholar] [CrossRef] [PubMed]
- Neitz, M.; Neitz, J. Numbers and ratios of visual pigment genes for normal red-green color vision. Science 1995, 267, 1013–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidoff, C.; Neitz, M.; Neitz, J. Genetic Testing as a New Standard for Clinical Diagnosis of Color Vision Deficiencies. Transl. Vis. Sci. Technol. 2016, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Winderickx, J.; Battisti, L.; Motulsky, A.G.; Deeb, S.S. Selective expression of human X chromosome-linked green opsin genes. Proc. Natl. Acad. Sci. USA 1992, 89, 9710–9714. [Google Scholar] [CrossRef] [Green Version]
- Rappon, J.; Chung, C.; Young, G.; Hunt, C.; Neitz, J.; Neitz, M.; Chalberg, T.W. Control of Myopia Using Diffusion Optics Spectacle Lenses: 12-Month Results of a Randomised Controlled, Efficacy and Safety Study (CYPRESS). Br. J. Ophthalmol. 2022; in revision. [Google Scholar]
- Neitz, M.; Carroll, J.; Renner, A.; Knau, H.; Werner, J.S.; Neitz, J. Variety of genotypes in males diagnosed as dichromatic on a conventional clinical anomaloscope. Vis. Neurosci. 2004, 21, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Neitz, M.; Neitz, J.; Jacobs, G.H. Spectral tuning of pigments underlying red-green color vision. Science 1991, 252, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Neitz, J.; Neitz, M.; Kainz, P.M. Visual pigment gene structure and the severity of human color vision defects. Science 1996, 274, 801–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; Abecasis, G.R. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, K.M.; Verhoeven, V.J.; Cumberland, P.; Bertelsen, G.; Wolfram, C.; Buitendijk, G.H.; Hofman, A.; van Duijn, C.M.; Vingerling, J.R.; Kuijpers, R.W.; et al. Prevalence of refractive error in Europe: The European Eye Epidemiology (E(3)) Consortium. Eur. J. Epidemiol. 2015, 30, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Neitz, M.; Patterson, S.S.; Neitz, J. Photopigment genes, cones, and color update: Disrupting the splicing code causes a diverse array of vision disorders. Curr. Opin. Behav. Sci. 2019, 30, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.-S.; Chu, R.-Y.; He, J.C.; Sun, S.-H.; Zhou, X.-T.; Zhao, N.-Q.; Hu, D.-N.; Hoffman, M.R.; Dai, J.H.; Qu, X.-M.; et al. Incidence of myopia in high school students with and without red-green color vision deficiency. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1598–1605. [Google Scholar] [CrossRef]
- Carroll, J.; McMahon, C.; Neitz, M.; Neitz, J. Flicker-photometric electroretinogram estimates of L: M cone photoreceptor ratio in men with photopigment spectra derived from genetics. J. Opt. Soc. Am. A 2000, 17, 499–509. [Google Scholar] [CrossRef]
- Hagen, L.A.; Arnegard, S.; Kuchenbecker, J.A.; Gilson, S.J.; Neitz, M.; Neitz, J.; Baraas, R.C. The association between L:M cone ratio, cone opsin genes and myopia susceptibility. Vis. Res. 2019, 162, 20–28. [Google Scholar] [CrossRef]
- Chamberlain, P.; de la Lazon Jara, P.; Arumugam, B.; Bullimore, M.A. Axial length targets for myopia control. Ophthalmic Physiol. Opt. 2021, 41, 523–531. [Google Scholar] [CrossRef]
- Young, T.L.; Deeb, S.S.; Ronan, S.M.; Dewan, A.T.; Alvear, A.B.; Scavello, G.S.; Paluru, P.C.; Brott, M.S.; Hayashi, T.; Holleschau, A.M.; et al. X-linked high myopia associated with cone dysfunction. Arch. Ophthalmol. 2004, 122, 897–908. [Google Scholar] [CrossRef] [Green Version]
- Bar Dayan, Y.; Levin, A.; Morad, Y.; Grotto, I.; Ben-David, R.; Goldberg, A.; Onn, E.; Avni, I.; Levi, Y.; Benyamini, O.G. The changing prevalence of myopia in young adults: A 13-year series of population-based prevalence surveys. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2760–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, C.J.; Andrew, T.; Mak, Y.T.; Spector, T.D. A susceptibility locus for myopia in the normal population is linked to the PAX6 gene region on chromosome 11: A genomewide scan of dizygotic twins. Am. J. Hum. Genet. 2004, 75, 294–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hysi, P.G.; Young, T.L.; Mackey, D.A.; Andrew, T.; Fernandez-Medarde, A.; Solouki, A.M.; Hewitt, A.W.; Macgregor, S.; Vingerling, J.R.; Li, Y.J.; et al. A genome-wide association study for myopia and refractive error identifies a susceptibility locus at 15q25. Nat. Genet. 2010, 42, 902–905. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.; Butterworth, J.; Bradley, D.T.; Hughes, A.E.; Soler, V.; Calvas, P.; Malecaze, F. A genome-wide association study provides evidence for association of chromosome 8p23 (MYP10) and 10q21.1 (MYP15) with high myopia in the French Population. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7983–7988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solouki, A.M.; Verhoeven, V.J.; van Duijn, C.M.; Verkerk, A.J.; Ikram, M.K.; Hysi, P.G.; Despriet, D.D.; van Koolwijk, L.M.; Ho, L.; Ramdas, W.D.; et al. A genome-wide association study identifies a susceptibility locus for refractive errors and myopia at 15q14. Nat. Genet. 2010, 42, 897–901. [Google Scholar] [CrossRef] [Green Version]
- Verhoeven, V.J.; Hysi, P.G.; Wojciechowski, R.; Fan, Q.; Guggenheim, J.A.; Hohn, R.; MacGregor, S.; Hewitt, A.W.; Nag, A.; Cheng, C.Y.; et al. Genome-wide meta-analyses of multiancestry cohorts identify multiple new susceptibility loci for refractive error and myopia. Nat. Genet. 2013, 45, 314–318. [Google Scholar] [CrossRef]
- Schwartz, M.; Haim, M.; Skarsholm, D. X-linked myopia: Bornholm eye disease. Linkage to DNA markers on the distal part of Xq. Clin. Genet. 1990, 38, 281–286. [Google Scholar] [CrossRef]
- Michaelides, M.; Johnson, S.; Bradshaw, K.; Holder, G.E.; Simunovic, M.P.; Mollon, J.D.; Moore, A.T.; Hunt, D.M. X-linked cone dysfunction syndrome with myopia and protanopia. Ophthalmology 2005, 112, 1448–1454. [Google Scholar] [CrossRef]
- Metlapally, R.; Michaelides, M.; Bulusu, A.; Li, Y.; Schwartz, M.; Rosenberg, T.; Hunt, D.; Moore, A.; Zuchner, S.; Rickman, C.; et al. Evaluation of the X-linked high-grade myopia locus (MYP1) with cone dysfunction and color vision deficiencies. Investig Ophthalmol. Vis. Sci. 2009, 50, 1552–1558. [Google Scholar] [CrossRef]
- Guo, X.; Xiao, X.; Li, S.; Wang, P.; Jia, X.; Zhang, Q. Nonsyndromic high myopia in Chinese family mapped to MYP1: Linkage confirmation and phenotypic characterization. Arch. Ophthalmol. 2010, 128, 1473–1479. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.; Guggenheim, J.A.; Bulusu, A.; Metlapally, R.; Abbott, D.; Malecaze, F.; Calvas, P.; Rosenberg, T.; Paget, S.; Creer, R.C.; et al. An international collaborative family-based whole-genome linkage scan for high-grade myopia. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3116–3127. [Google Scholar] [CrossRef]
- Ratnamala, U.; Lyle, R.; Rawal, R.; Singh, R.; Vishnupriya, S.; Himabindu, P.; Rao, V.; Aggarwal, S.; Paluru, P.; Bartoloni, L.; et al. Refinement of the X-linked nonsyndromic high-grade myopia locus MYP1 on Xq28 and exclusion of 13 known positional candidate genes by direct sequencing. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6814–6819. [Google Scholar] [CrossRef] [Green Version]
- Bush, W.S.; Moore, J.H. Chapter 11: Genome-wide association studies. PLoS Comput. Biol. 2012, 8, e1002822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neitz, J.; Kuchenbecker, J.A.; Neitz, M. Ophthalmic Lenses for Treating Myopia. U.S. Patent 10,571,717, 25 February 2020. [Google Scholar]
- Rappon, J.; Neitz, J.; Neitz, M.; Young, G.; Chalberg, T.W. CYPRESS 12-month Results: Safety and Efficacy from a Pivotal Study of Novel Spectacle Lenses Designed to Reduce Myopia Progression. Optom. Vis. Sci. 2020, 97, E-abstract 200036. [Google Scholar]
- Patterson, S.S.; Kuchenbecker, J.A.; Anderson, J.R.; Bordt, A.S.; Marshak, D.W.; Neitz, M.; Neitz, J. An S-cone circuit for edge detection in the primate retina. Sci. Rep. 2019, 9, 11913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neitz, J.; Neitz, M. Evolution of the circuitry for conscious color vision in primates. Eye (Lond. Engl.) 2017, 31, 286–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallman, J.; Winawer, J. Homeostasis of eye growth and the question of myopia. Neuron 2004, 43, 447–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorn, F.; Grice, K.; Held, R.; Gwiazda, J. Myopia: Nature, nurture, and the blur hypothesis. In Myopia Updates II; Springer: Tokyo, Japan, 2000; pp. 89–93. [Google Scholar]
- Schmid, K.L.; Brinkworth, D.R.; Wallace, K.M.; Hess, R. The effect of manipulations to target contrast on emmetropization in chick. Vis. Res. 2006, 46, 1099–1107. [Google Scholar] [CrossRef] [Green Version]
- Hess, R.F.; Schmid, K.L.; Dumoulin, S.O.; Field, D.J.; Brinkworth, D.R. What image properties regulate eye growth? Curr. Biol. 2006, 16, 687–691. [Google Scholar] [CrossRef]
- Diether, S.; Wildsoet, C.F. Stimulus requirements for the decoding of myopic and hyperopic defocus under single and competing defocus conditions in the chicken. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2242–2252. [Google Scholar] [CrossRef] [Green Version]
- Schmid, K.L.; Wildsoet, C.F. Contrast and spatial-frequency requirements for emmetropization in chicks. Vis. Res. 1997, 37, 2011–2021. [Google Scholar] [CrossRef]
- Lam, C.S.Y.; Tang, W.C.; Tse, D.Y.; Lee, R.P.K.; Chun, R.K.M.; Hasegawa, K.; Qi, H.; Hatanaka, T.; To, C.H. Defocus Incorporated Multiple Segments (DIMS) spectacle lenses slow myopia progression: A 2-year randomised clinical trial. Br. J. Ophthalmol. 2020, 104, 363–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullimore, M.A.; Richdale, K. Myopia Control 2020: Where are we and where are we heading? Ophthalmic Physiol. Opt. 2020, 40, 254–270. [Google Scholar] [CrossRef] [PubMed]
- Morgan, I.G.; Rose, K.A.; Ellwein, L.B. Is emmetropia the natural endpoint for human refractive development? An analysis of population-based data from the refractive error study in children (RESC). Acta Ophthalmol. 2010, 88, 877–884. [Google Scholar] [CrossRef]
- Wiesel, T.N.; Raviola, E. Myopia and eye enlargement after neonatal lid fusion in monkeys. Nature 1977, 266, 66–68. [Google Scholar] [CrossRef]
- Wallman, J.; Turkel, J.; Trachtman, J. Extreme myopia produced by modest change in early visual experience. Science 1978, 201, 1249–1251. [Google Scholar] [CrossRef]
- Smith, E.L.; Hung, L.-F.; Huang, J. Protective effects of high ambient lighting on the development of form-deprivation myopia in rhesus monkeys. Investig. Ophthalmol. Vis. Sci. 2012, 53, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Durant, S.; Clifford, C.W.; Crowder, N.A.; Price, N.S.; Ibbotson, M.R. Characterizing contrast adaptation in a population of cat primary visual cortical neurons using Fisher information. J. Opt. Soc. Am. A Opt. Image Sci. Vis. 2007, 24, 1529–1537. [Google Scholar] [CrossRef]
- Semeraro, F.; Forbice, E.; Nascimbeni, G.; Cillino, S.; Bonfiglio, V.M.E.; Filippelli, M.E.; Bartollino, S.; Costagliola, C. Ocular Refraction at Birth and Its Development During the First Year of Life in a Large Cohort of Babies in a Single Center in Northern Italy. Front. Pediatr. 2019, 7, 539. [Google Scholar] [CrossRef]
- Mutti, D.O.; Mitchell, G.L.; Jones, L.A.; Friedman, N.E.; Frane, S.L.; Lin, W.K.; Moeschberger, M.L.; Zadnik, K. Axial growth and changes in lenticular and corneal power during emmetropization in infants. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3074–3080. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.L., 3rd. Prentice Award Lecture 2010: A case for peripheral optical treatment strategies for myopia. Optom. Vis. Sci. Off. Publ. Am. Acad. Optom. 2011, 88, 1029–1044. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Haplotype 1 | Amino Acids 2 | N 3 | Median/Mean SER OS 4 | % Exon-3 Skipping | Split-Halves Rank 5 |
|---|---|---|---|---|---|
| AACGGTGG | MVVVA | 12 | −3.16/−3.14 | 16 | 1 |
| AACGGCAT | MVAIS | 30 | −1.62/−2.00 | 3 | 2.49 |
| AACGGTAT | MVVIS | 23 | −2.01/−2.57 | 0 | 2.51 |
| AACGGCAG | MVAIA | 67 | −1.05/−1.71 | 2 | 4.25 |
| GCGGGCAG | LVAIA | 77 | −0.97/−1.38 | 14 | 4.75 |
| GCGGGCAT | LVAIS | 157 | −0.99/−1.38 | 1 | 6 |
| GCGATCAT | LIAIS | 25 | −1.10/−1.30 | 1 | 7 |
| AAGGGCAT | MVAIS | 9 | +0.15/+0.51 | 0 | 8.99 |
| GCGGGGAT | LVVIS | 5 | −1.06/−1.04 | 1 | 9 |
| GCCGGCAT | LVAIS | 4 | −0.05/−0.25 | 8 | 9.02 |
| GCCGGCAG | LVAIA | 4 | +0.54/+0.92 | 2 | 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neitz, M.; Wagner-Schuman, M.; Rowlan, J.S.; Kuchenbecker, J.A.; Neitz, J. Insight from OPN1LW Gene Haplotypes into the Cause and Prevention of Myopia. Genes 2022, 13, 942. https://doi.org/10.3390/genes13060942
Neitz M, Wagner-Schuman M, Rowlan JS, Kuchenbecker JA, Neitz J. Insight from OPN1LW Gene Haplotypes into the Cause and Prevention of Myopia. Genes. 2022; 13(6):942. https://doi.org/10.3390/genes13060942
Chicago/Turabian StyleNeitz, Maureen, Melissa Wagner-Schuman, Jessica S. Rowlan, James A. Kuchenbecker, and Jay Neitz. 2022. "Insight from OPN1LW Gene Haplotypes into the Cause and Prevention of Myopia" Genes 13, no. 6: 942. https://doi.org/10.3390/genes13060942