
Genome-Wide Association Studies Reveal Candidate Genes Related to Stem Diameter in Cucumber (Cucumis sativus L.)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Phenotypic Data Collection and Analysis
2.3. Genetic Diversity of Stem Diameter in Germplasm
2.4. Resequencing Data Analysis and Genotype
2.5. Genome-wide Association Analysis
2.6. Linkage Disequilibrium Analysis
2.7. Prediction of Candidate Genes
2.8. Candidate Genes Analysis
3. Results
3.1. Genetic Diversity of Stem Diameter in Core Germplasms
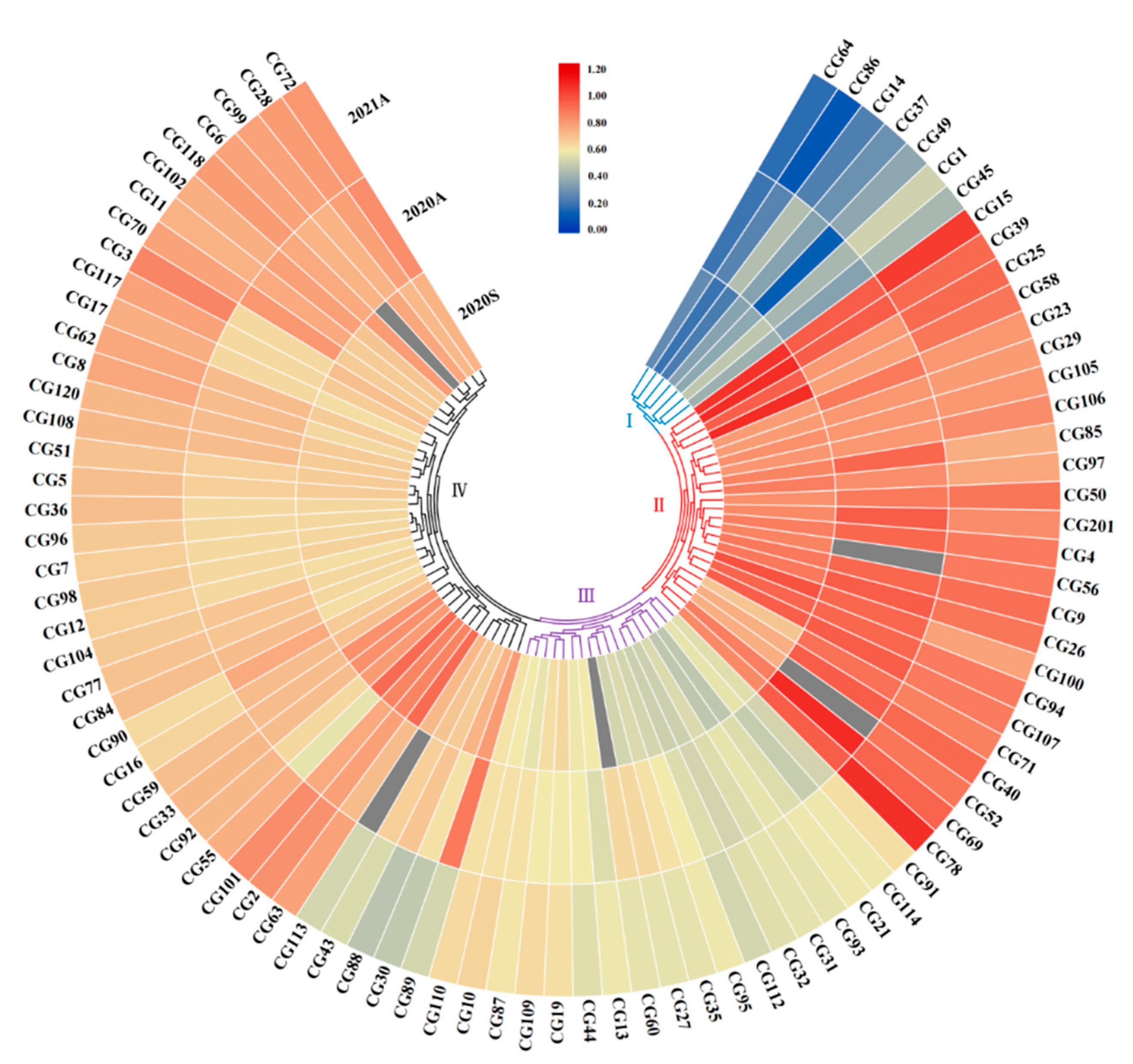
3.2. Clustering Analysis of Stem Diameter in Core Germplasms
3.3. Genome-Wide Association Study of Stem Diameter in Core Germplasms
3.4. Analysis of Candidate Genes for GWAS Stable Loci of Stem Diameter in Core Germplasms
3.4.1. gSD1.1 Candidate Gene Analysis
3.4.2. gSD2.1 Candidate Gene Analysis
3.4.3. gSD3.1 Candidate Gene Analysis
3.4.4. gSD5.2 Candidate Gene Analysis
3.4.5. gSD6.1 Candidate Gene Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. 2020. Available online: http://www.fao.org/faostat/en/#data (accessed on 12 April 2022).
- Wang, B.; Smith, S.M.; Li, J.Y. Genetic Regulation of Shoot Architecture. Annu. Rev. Plant Biol. 2018, 69, 437–468. [Google Scholar] [CrossRef]
- Yang, S.; Wang, L.P. Analysis on Heterosis and Combining Ability of Six Mini-cucumber Inbred Lines. J. Changjiang Veg. 2016, 20, 44–49. [Google Scholar]
- Cao, G.; Zhang, G.B.; Yu, J.H.; Ma, Y.X. Effects of Different LED Light Qualities on Cucumber Seedling Growth and Chlorophyll Fluorescence Parameters. Sci. Agric. Sin. 2013, 46, 1297–1304. [Google Scholar]
- Tomar, G.S.; Khajanji, S.N. Effect of organic manuring and mineral fertilizer on the growth, yield and economics of soybean [Glycine max (L.) Merrill]. Int. J. Agric. Sciences. 2009, 5, 590–594. [Google Scholar]
- Kamil, S.S.; Sarkawt, A.A.; Ismael, A.H.; Pshtiwan, H.A. Effect of Bio-fertilizer and Chemical Fertilizer on Growth and Yield in Cucumber (Cucumis sativus) in Green House Condition. Pak. J. Biol. Sci. 2015, 18, 129–134. [Google Scholar]
- Qi, Z.Y.; Li, J.X.; Zou, X.X.; Cao, L.W.; Rao, L.L.; Yu, J.L.; Chen, L.P. Genetic Analysis of Plant Type Traits of Muskmelon. J. Agric. Biotechnol. 2015, 23, 302–310. [Google Scholar]
- Rao, P.G.; Behera, T.K.; Gaikwad, A.B.; Munshi, A.D.; Srivastava, A.; Boopalakrishnan, G. Genetic analysis and QTL mapping of yield and fruit traits in bitter gourd (Momordica charantia L.). Sci. Rep. 2021, 11, 4109. [Google Scholar] [CrossRef]
- Lin, T.; Wang, M.; Peng, Q.; Jiang, B.; Lin, Y.E.; Liu, W.R.; Liang, Z.J.; Yan, J.Q.; Ye, Z.B.; He, X.Y. Genetic Analysis and QTL Mapping of Important Agronomic Traits of Chieh-qua. Chin. Hortic. Soc. 2019, 3, 198. [Google Scholar]
- Li, X.Z.; Yuan, X.J.; Jiang, S.; Pan, J.S.; Deng, S.L.; Wang, G.; Le He, H.; Wu, A.Z.; Zhu, L.H.; Koba, T.; et al. Detecting QTLs for plant architecture traits in cucumber (Cucumis sativus L.). Breed. Sci. 2008, 58, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.J.; Yuan, J.J.; Zheng, S.L.; Kong, F.L.; Zhong, L.; Cheng, Q.B.; Dou, P. A crop stem thickness measurement caliper. CN Patent CN205102741U[P], 23 March 2016. [Google Scholar]
- Zhang, L. Advance of horticultural plant genomes. Hortic. Plant J. 2019, 5, 229–230. [Google Scholar] [CrossRef]
- Shang, Y.; Ma, Y.; Zhou, Y.; Zhang, H.; Duan, L.; Chen, H.; Zeng, J.; Zhou, Q.; Wang, S.; Gu, W.; et al. Biosynthesis, regulation, and domestication of bitterness in cucumber. Science 2014, 346, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Bo, K.; Miao, H.; Wang, M.; Xie, X.; Song, Z.; Xie, Q.; Shi, L.; Wang, W.; Wei, S.; Zhang, S.; et al. Novel loci fsd6.1 and Csgl3 regulate ultra-high fruit spine density in cucumber. Theor. Appl. Genet. 2019, 132, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Bo, K.; Wei, S.; Wang, W.; Miao, H.; Dong, S.; Zhang, S.; Gu, X. QTL mapping and genome-wide association study reveal two novel loci associated with green flesh color in cucumber. BMC Plant Biol. 2019, 19, 243. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Bao, K.; Reddy, U.K.; Bai, Y.; Hammar, S.A.; Jiao, C.; Wehner, T.C.; Ramírez-Madera, A.O.; Weng, Y.; Grumet, R.; et al. The USDA cucumber (Cucumis sativus L.) collection: Genetic diversity, population structure, genome-wide association studies, and core collection development. Hortic. Res. 2018, 5, 64. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; VandenLangenberg, K.; Wen, C.; Wehner, T.C.; Weng, Y. QTL mapping of downy and powdery mildew resistances in PI 197088 cucumber with genotyping-by-sequencing in RIL population. Theor. Appl. Genet. 2018, 131, 597–611. [Google Scholar] [CrossRef]
- Liu, X.P.; Lu, H.W.; Liu, P.N.; Miao, H.; Bai, Y.L.; Gu, X.; Zhang, S.P. Identification of novel loci and candidate genes for cucumber downy mildew resistance using GWAS. Plants 2020, 9, 1659. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.X.; Li, X.X.; Song, J.P.; Lin, Y.E.; Zhang, X.H.; Qiu, Y.; Yang, W.L.; Lou, Q.F.; Wang, H.P. Genome-wide association study of powdery mildew resistance of cucumber core germplasms. Acta Hortic. Sin. 2021, 48, 1371–1385. [Google Scholar]
- Wang, W.P.; Song, Z.C.; Bo, K.L.; Dong, S.Y.; Wei, S.; Miao, H.; Li, J.B. Evaluation and genome-wide association study (GWAS) of low-temperature tolerance at seedling stage in cucumber core germplasm. J. Plant Genet. Resour. 2019, 20, 1606–1612. [Google Scholar]
- Zhang, S.; Miao, H.; Song, Z.C.; Bo, K.L.; Wang, Y.; Gu, X.F.; Zhang, S.P. Evaluation and genome-wide association study (GWAS) of thermotolerance for cucumber at the germination stage. J. Plant Genet. Resour. 2019, 20, 335–346. [Google Scholar]
- Wei, S.; Zhang, S.; Bo, K.L.; Wang, W.P.; Miao, H.; Dong, S.Y.; Gu, X.F.; Zhang, S.P. Evaluation and genome-wide association study (GWAS) of seedling thermotolerance in cucumber core germplasm. J. Plant Genet. Resour. 2019, 20, 1223–1231. [Google Scholar]
- Lv, J.; Qi, J.J.; Shi, Q.X.; Shen, D.; Zhang, S.P.; Shao, G.J.; Li, H.; Sun, Z.Y.; Weng, Y.Q.; Shang, Y.; et al. Genetic diversity and population structure of cucumber (Cucumis sativus L.). PLoS ONE 2012, 7, e46919. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.J.; Liu, X.; Shen, D.; Miao, H.; Xie, B.Y.; Li, X.X.; Zeng, P.; Wang, S.; Shang, Y.; Gu, X.; et al. A genomic variation map provides insights into the genetic basis of cucumber domestication and diversity. Nat. Genet. 2013, 45, 1510–1515. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute. SAS/OR 9.3 User’s Guide: Mathematical Programming Examples; SAS Institute: Cary, NC, USA, 2012. [Google Scholar]
- Yan, Q. Research on differential privacy decision tree method based on Pearson correlation coefficient. Master’s Thesis, Guangxi Normal University, Guilin, China, 2021. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippert, C.; Listgarten, J.; Liu, Y.; Kadie, C.M.; Davidson, R.I.; Heckerman, D. FaST linear mixed models for genome-wide association studies. Nat. Methods 2011, 8, 833–835. [Google Scholar] [CrossRef] [PubMed]
- Yin, L. CMplot: Circle Manhattan Plot; GitHub, Inc.: San Francisco, CA, USA, 2018. [Google Scholar]
- Available online: http://cucurbitgenomics.org/ (accessed on April 12, 2022).
- TAIR. Available online: https://www.arabidopsis.org/ (accessed on 9 December 2021).
- Xie, Q.; Liu, P.N.; Shi, L.X.; Miao, H.; Bo, K.L.; Gu, X.F.; Zhang, S.P. Combined fine mapping, genetic diversity, and transcriptome profiling reveals that the auxin transporter gene ns plays an important role in cucumber fruit spine development. Theor. Appl. Genet. 2018, 131, 1239–1252. [Google Scholar] [CrossRef] [PubMed]
- Jarošová, J.; Kundu, J.K. Research article Validation of reference genes as internal control for studying viral infections in cereals by quantitative real-time RT-PCR. BMC Plant Biol. 2010, 10, 146. [Google Scholar] [CrossRef] [Green Version]
- Trun, N.J.; Stader, J.; Lupas, A.; Kumamoto, C.; Silhavy, T.J. Two cellular components, PrlA and SecB, that recognize different sequence determinants are required for efficient protein export. J. Bacteriol. 1989, 170, 5928–5930. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Kim, S.G.; Lee, M.; Lee, I.; Park, H.Y.; Seo, P.J.; Jung, J.H.; Kwon, E.J.; Suh, S.W.; Paek, K.H.; et al. HD-ZIP III Activity Is Modulated by Competitive Inhibitors via a Feedback Loop in Arabidopsis Shoot Apical Meristem Development. Plant Cell 2008, 20, 920–933. [Google Scholar] [CrossRef] [Green Version]
- Engelsdorf, T.; Will, C.; Hofmann, J.; Schmitt, C.; Merritt, B.B.; Rieger, L.; Frenger, M.S.; Marschall, A.; Franke, R.B.; Pattathil, S.; et al. Cell wall composition and penetration resistance against the fungal pathogen Colletotrichum higginsianum are affected by impaired starch turnover in Arabidopsis mutants. J. Exp. Bot. 2017, 68, 701–713. [Google Scholar]
- Schweingruber, F.H.; Börner, A. Stem anatomical structures of major taxonomic units. In The Plant Stem; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Huang, J. The Comparative Anatomical Observation of Cucumber. Ph.D. Dissertation, Northeast Agricultural University, Harbin, China, 2010. [Google Scholar]
- Hu, D.Y. Studies of the Mechanisms of Improvement in the Growth of Autumn Cucumbers and Soil Environment with Aerated Irrigation. Ph.D. Dissertation, Hunan Agricultural University, Changsha, China, 2014. [Google Scholar]
- Jois, S.N.; Prasad, N.K.; D’Souza, L. Morphology of Cucumber Plants as Influenced by Pranic Agriculture. Asian J. Agric. Res. 2016, 11, 33–35. [Google Scholar] [CrossRef] [Green Version]
- Lv, P.C.; Wang, C.H.; Lin, Y.X.; Jiang, H.J.; Dong, X.L.; Qi, W.B.; Do, X.C. Effect of compound microbial agents on growth and quality of cucumber in greenhouse. Anhui Agric. Sci. Bull. 2020, 26, 47–49. [Google Scholar]
- Ndimba, B.K.; Chivasa, S.; Hamilton, J.M.; Simon, W.J.; Slabas, A.R. Proteomic analysis of changes in the extracellular matrix of arabidopsis cell suspension cultures induced by fungal elicitors. Proteomics 2010, 3, 1047–1059. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X.; Lin, Z.; Wang, J.; Liu, H.; Zhou, L.; Zhong, S.; Li, Y.; Zhu, C.; Lai, J.; et al. A Large Transposon Insertion in the stiff1 Promoter Increases Stalk Strength in Maize. Plant Cell 2020, 32, 152–165. [Google Scholar] [CrossRef] [Green Version]
- Mayer, K.F.; Schoof, H.; Haecker, A.; Lenhard, M.; Jürgens, G.; Laux, T. Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell 1998, 95, 805. [Google Scholar] [CrossRef] [Green Version]
- Tu, B.; Tao, Z.; Wang, S.; Zhou, L.; Zheng, L.; Zhang, C.; Li, X.; Zhang, X.; Yin, J.; Zhu, X.; et al. Loss of Gn1a/OsCKX2 confers heavy-panicle rice with excellent lodging resistance. J. Integr. Plant Biol. 2022, 64, 23–38. [Google Scholar] [CrossRef]
- Mizokami, Y.; Sugiura, D.; Watanabe, C.K.A.; Betsuyaku, E.; Inada, N.; Terashima, I. Elevated CO2-induced changes in mesophyll conductance and anatomical traits in wild type and carbohydrate-metabolism mutants of Arabidopsis. J. Exp. Bot. 2019, 70, 4807–4818. [Google Scholar] [CrossRef]
- Wang, S.; Lu, G.; Hou, Z.; Luo, Z.; Wang, T.; Li, H.; Zhang, J.; Ye, Z. Members of the tomato FRUITFULL MADS-box family regulate style abscission and fruit ripening. J. Exp. Bot. 2014, 65, 3005–3014. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Maximum | Minimum | Mean ± SD | Kurtosis | Skewness | (%)CV | p Value |
|---|---|---|---|---|---|---|---|
| 2020S | 0.91 | 0.34 | 0.67 ± 0.12 | 0.41 | −0.39 | 17.32 | 0.07 > 0.05 |
| 2020A | 0.74 | 0.25 | 0.51 ± 0.11 | 0.97 | −0.61 | 15.10 | 0.10 > 0.05 |
| 2021A | 1.02 | 0.35 | 0.76 ± 0.14 | 0.11 | −0.86 | 16.08 | 0.20 > 0.05 |
| Season | Signal Sites | SNP | Chr. | Physical Position (bp) | −log10 p Value |
|---|---|---|---|---|---|
| 2020S | gSD1.1 | S1_15337811 | 1 | 15,337,811 | 6.95 |
| gSD2.1 | S2_3445551 | 2 | 3,445,551 | 6.13 | |
| gSD3.1 | S3_7785079 | 3 | 7,785,079 | 6.01 | |
| gSD3.2 | S3_16649668 | 3 | 16,649,668 | 6.11 | |
| gSD5.2 | S5_25600905 | 5 | 25,600,905 | 6.44 | |
| gSD6.1 | S6_17507945 | 6 | 17,507,945 | 6.03 | |
| 2020A | gSD1.1 | S1_15337811 | 1 | 15,337,811 | 6.05 |
| gSD3.1 | S3_7785079 | 3 | 7,785,079 | 6.24 | |
| gSD5.1 | S5_20992082 | 5 | 20,992,082 | 6.41 | |
| gSD5.2 | S5_25600905 | 5 | 25,600,905 | 6.03 | |
| gSD6.1 | S6_17507945 | 6 | 17,507,945 | 6.05 | |
| 2021A | gSD1.1 | S1_15337811 | 1 | 15,337,811 | 7.42 |
| gSD2.1 | S2_3445551 | 2 | 3,445,551 | 8.46 | |
| gSD3.1 | S3_7785079 | 3 | 7,785,079 | 7.81 | |
| gSD4.1 | S4_6318227 | 4 | 6,318,227 | 6.78 | |
| gSD6.1 | S6_17507945 | 6 | 17,507,945 | 6.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Dong, S.; Miao, H.; Liu, X.; Dai, Z.; Li, X.; Gu, X.; Zhang, S. Genome-Wide Association Studies Reveal Candidate Genes Related to Stem Diameter in Cucumber (Cucumis sativus L.). Genes 2022, 13, 1095. https://doi.org/10.3390/genes13061095
Yang Y, Dong S, Miao H, Liu X, Dai Z, Li X, Gu X, Zhang S. Genome-Wide Association Studies Reveal Candidate Genes Related to Stem Diameter in Cucumber (Cucumis sativus L.). Genes. 2022; 13(6):1095. https://doi.org/10.3390/genes13061095
Chicago/Turabian StyleYang, Yingying, Shaoyun Dong, Han Miao, Xiaoping Liu, Zhuonan Dai, Xiangsheng Li, Xingfang Gu, and Shengping Zhang. 2022. "Genome-Wide Association Studies Reveal Candidate Genes Related to Stem Diameter in Cucumber (Cucumis sativus L.)" Genes 13, no. 6: 1095. https://doi.org/10.3390/genes13061095